Effects of Endotoxin and Catecholamines on Hepatic Mitochondrial Respiration
Texte intégral
Figure
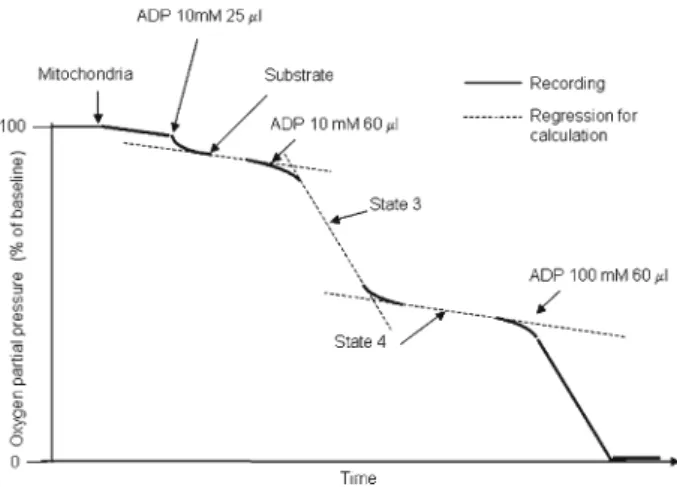
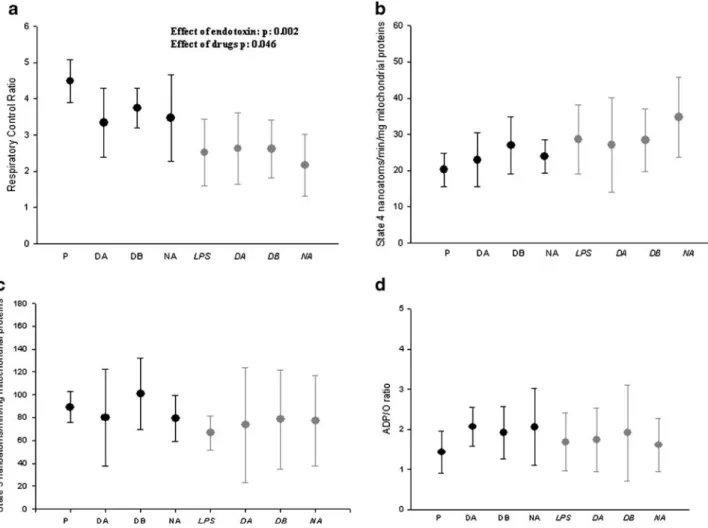


Documents relatifs
Mitochondrial dynamics, Legionella pneumophila, high-content microscopy, Seahorse, mitochondrial respiration assay, mitochondrial fission, mitochondrial morphology,
Susisuchus jaguaribensis has other characteristics shared with eusuchians, indicating their close relationship: (1) participation limited or absent of the frontal within
determining paint quality are consistency, fineness of grind, color, hiding power, brushability, durability, drying time, leveling and gloss.. The quality of the
Both of the indices, the RQI and RCI, of this thesis are based off the strain energy performance of the stable substructures of the original structural system.. Since
A number of scientific topics are addressed with this data-set that are briefly presented in this review: the study of foreground Galactic cirrus at high spatial resolution, telling
This issue of Topics in Catalysis celebrates the successful completion termination of BIOTRAINS, an FP-7 funded Marie Curie Initial Training Network (MC-ITN).. Initiated and
L’étude a démontré que de 100 à 270 jours après la GCSH, le taux global d’événements liés au critère d’évaluation primaire (décès, maladie à CMV, infection
The I/(I+II) ratio corresponds to the fraction of respiration driven by complex I with respect to the maximal respiratory rate driven by complexes I and II. Pooled results
