A 3D synthetic extracellular matrix that can be dissolved to
interrogate the cellular local microenvironment on demand
By
Jorge L. Valdez Macias
B.S. Bioengineering Arizona State University, 2010
MASSACHUSETTS INSTITUTE OF TECHNOLOGY
JUN 0
8
2016
LIBRARIES
ARCHIVES
Submitted to the Department of Biological Engineering in Partial Fulfillment of the Requirements for the Degree of
Doctor of Philosophy in Biological Engineering at the
MASSACHUSETTS INSTITUTE OF TECHNOLOGY June 2016
C 2016 Massachusetts Institute of Technology. All rights reserved.
Signature of author:
Certified by:
Approved by:
Signature redacted
V Department of Biological Engineering
May 20, 2016
Signature redacted
Linda G. Griffith S.E.T.I. Professor o d Mechanical Engineering Thesis Supervisor
Signature redacted
Forest M. White Professor of Biological Engineering Chair, Graduate Program Committee
This doctoral thesis has been examined by the following
committee members:
Linda G. Griffith, Ph.D.
Department of Biological Engineering Department of Mechanical Engineering Massachusetts Institute of technology Alan J. Grodzinsky, Sc.D.
Department of Biological Engineering Department of Mechanical Engineering Department of Electrical Engineering Massachusetts Institute of Technology Paula T. Hammond, Ph.D.
Department of Chemical Engineering Massachusetts Institute of Technology Darrell D. Irvine, Ph.D.
Department of Biological Engineering
A 3D synthetic extracellular matrix that can be dissolved to interrogate the cellular local microenvironment on demand
by
Jorge Luis Valdez Macias
Submitted to the Department of Biological Engineering on May 20, 2016, in partial fulfillment of the
requirements for the degree of
Doctor of Philosophy in Biological Engineering Abstract
In vitro multi-cellular 3D cultures can provide tremendous insight into pathologies arising from dysregulation of extracellular communication by recapitulating key features of the microenvironment, such as ECM-cytokine
interactions, spatially regulated cell-cell interactions, and matrix biophysical cues. Relevant 3D in vitro systems have been limited by the difficulty to assay the local microenvironment that cells directly experience, and by the lack of modularity of naturally-derived hydrogels such as Matrigel and collagen gels. Here, we exploit the modularity of synthetic hydrogels to overcome these limitations by designing a novel 3D culture system that can be locally and accurately interrogated.
We begin by investigating the underlying principles directing morphogenesis in 3D synthetic PEG hydrogel cultures by screening the relevant parameter space
-specifically, the intertwined relationship between adhesion and matrix degradation, the effect of cell-cell contacts, and cell-mediated matrix deposition. We use
vasculogenesis of human iPSC-derived endothelial cells as a case study. Understanding vasculogenesis is a crucial requirement for engineering tissue models exceeding 100 gm due to nutrient diffusion limitations. Here we define microenvironment conditions that are permisive of the formation of 3D
interconnected structures of iPSC endothelial cells in PEG hydrogels.
Lastly, we adapted the synthetic hydrogels to enable local interrogation of the 3D microenvironment -a hitherto unavailable feature. Here we report a novel hydrogel system that can be used to both encapsulate complex co-cultures, and recover intact cells and local cytokines from the 3D matrix by repurposing a mutant of sortase A (SrtA). We demonstrate SrtA-mediated gel dissolution preserves
important cell morphological features and can be used to measure recovered cytokines much more accurately compared to other proteolytic degradation
methods. We observe discrepancies between the cytokine concentrations inside the gel and those in the culture medium. These local measurements reveal behaviors in response to an inflammatory stimulus that cannot be captured by monitoring only the cytokine concentrations in the medium. This system allows for better
understanding of protein communication networks in relevant complex 3D multicellular cultures and can aid in identifying and testing targets in drug discovery and development.
Thesis Supervisor: Linda G. Griffith
Acknowledgements
First, I thank my advisor Linda Griffith for welcoming me into her lab. Her inexhaustible drive for improving people's lives through science has been a constant source of inspiration to push the boundaries of the bioengineering field. It is under her mentorship that I have grown as a scientist. I am immensely grateful for the
attention and resources she has given me -from scientific discussions to helping me prepare conference presentations.
I thank my committee for their scientific insights and professionalism. Paula Hammond has taught me the huge potential of advanced polymeric materials in life sciences, always with a smile on her face. Having the privilege of taking a class taught by Alan Grodzinsky was an utter enjoyment and one of the highlights of my
doctorate training. And being a teaching assistant for Darrell Irvine was more educational than many other classes I have taken. I am grateful for having the support of such bright and outstanding scientists.
I thank my undergraduate research advisor Brent Vernon for taking me into his lab, and Bae Hoon Lee for teaching me the lab skills that would make my future research experience a much easier process.
I thank Michael Caplan for believing in me and encouraging me to apply to the BE Department at MIT and advising me through the process.
I thank the members of the Griffith and Lauffenburger labs. I thank Caroline Chopko for the countless scientific discussions that led to a lot of the ideas
presented in this thesis, and her company during long late nights in lab. Edgar Sanchez and Jaime Rivera for their friendship and continued support. I thank the colleagues that I've shared my office with for the last six years for making it an amicable and stimulating environment: Miles Miller, Shannon Hughes, Allison Claas, Alex Wang, Alex Brown, Stephanie Wang, Charlotte Fonta. I thank Hsinhwa Lee, our lab manager, for making it so we never have to worry about anything but the science when being in lab, and for being a motherly figure for those of us whose families are abroad.
I thank the professors and colleagues I have had the pleasure of working with: Forest White, Roger Kamm, Harry Asada, Ron Weiss, Daniel Rothenberg, Elizabeth Gordon, Jordan Whisler, Sebastian Uzel, Vivek Sivathanu, Vincent Chan, and many others.
I thank the BE community as a whole, especially those who I have made close connections with: Anthony Soltis, Chris Ng, Matthew Adendorff, Joel Wagner, Ran Li, Jennifer Brophy, Marcus Parrish, Alice Tzeng, and Nina Lee.
I thank the NSF EBICS center for funding and for creating a stimulating scientific community.
Finally, I thank my family for their unconditional love and support.
Especially my mother, whose devotion and sacrifice have been instrumental for my success. To you, I dedicate this thesis.
Contents
1 Introduction...14
1.1 In vitro three-dimensional tissue models... 14
1.2 Synthetic biomaterials as a modular ECM...16
1.3 Key features of the cell microenvironment ... 18
1.4 T hesis objectives... 21
2 Developing a workflow to promote a desired morphogenesis
response: vasculogenesis case study ...
23
2.1 Introduction ... 23
2 .2 R esu lts ... 29
2.2.1 IPSC-derived endothelial cells as a cell model for vasculogenesis...29
2.2.1.1 2D attachment assay reveals differential attachment to RGD, SYNKRGD, and GFOGER for human IPS-derived endothelial cells ... 30
2.2.1.2 Increased 2D attachment correlates with more complex vascular 3D netw orks of hIPSE Cs ... 32
2.2.1.3 Hydrogel MMP-sensitivity and crosslinker:macromer stoichiometric ratio affect hIPSECs network formation... 34
2.2.1.4 Fibronectin-binding peptide increases fibronectin accumulation by IPSC-derived endothelial cells... 37
2.2.2 C ell-cell effects... 39
2.2.2.1 Co-culture of hIPSECs with mesenchymal stem cells promotes com plex structures ... 39
2.2.2.2 hIPSECs seeding density affects network formation ... 41
2.2.2.3 Comparison of hIPSECs to human brain vascular endothelial cells underscores cell context dependence ... 42
2 .3 D iscu ssion ... 44
2.4.1 Peptides and m acrom ers ... 49
2.4.2 C ell culture ... 49
2.4.3 2D attachm ent assay ... 50
2.4.4 Human IPSC endothelial cell encapsulation... 50
2.4.5 Immunostaining and imaging...51
2.4.6 Network feature quantification ... 51
3 Adapting synthetic PEG hydrogel system to allow for fast,
facile, and cell friendly dissolution and cell recovery using
sortase A (SrtA) ...
52
3.1 Introduction ... 52
3 .2 R esu lts ... 54
3.2.1 Using SrtA to crosslink and dissolve synthetic PEG hydrogels ... 54
3.2.2 SrtA-mediated cell recovery from PEG hydrogels yields viable cells and preserves cell-cell contacts ... 59
3.2.3 Dissolution kinetics of PEG hydrogels using SrtA and soluble GGG .... 63
3.3 D iscu ssion ... 66
3.4 M aterials and M ethods ... 70
3.4.1 Peptides and functionalized polymers...70
3.4.2 Cell culture of endometrial cells. ... 70
3.4.3 Synthesis of PEG-GGG/SynKRGD, PEG-MMP-LPRTG, and PEG-GGG m acrom ers...7 1 3.4.4 Encapsulation of endometrial co-culture and primary rat hepatocytes in SrtA-cross-linked functionalized PEG hydrogel for functional assay and cell recovery . ... .. ... --- 7 1 3.4.5 PEG norbornene (PEG-NB) hydrogel synthesis for dissolution rate qu an tification ... 72
3.4.6 PEG vinyl sulfone (PEGVS) hydrogel synthesis for dissolution rate
qu an tification ... 73
3.4.7 3D stromal and epithelial (cell lines) co-culture cell encapsulation in PEG vinyl sulfone hydrogels for signaling studies...74
3.4.8 Hydrogel dissolution quantification... 75
3.4.9 ELISA for IG FBP-1. ... 75
3.4.10 Immunostaining and Microscopy. ... 76
4 Leveraging SrtA-mediated gel dissolution to interrogate
local protein cell-cell communication networks in complex 3D
cultures 78
4.1 Introduction ... 784 .2 R esu lts ... 8 0 4.2.1 SrtA/GGG incubation does not affect phosphorylation of ERK and MET in tw o colon cancer cell lines ... 80
4.2.2 Incubating signaling inflammatory cytokines with SrtA does not deplete their concentration compared to trypsin and Liberase ... 81
4.2.3 SrtA-mediated gel dissolution shows discrepancies between local in-gel and culture m edium concentrations ... 83
4.2.4 SrtA-mediated gel dissolution increases temporal sensitivity to an inflammatory response to IL-16 and reveals temporal behaviors not captured in th e m ediu m ... 87
4.2.5 Protein interaction networks derived from local cytokine measurements contain richer information than those derived from medium measurements .... 88
4.3 D iscussion ... 90
4.4 M aterials and M ethods ... 94
4.4.1 Cell culture of colon cancer cell lines for phosphosignaling study ... 94
4.4.3 Encapsulation of endometrial cell line co-cultures for signaling studies 95
4.4.4 Incubation of cytokines with SrtA and proteases ... ... 96
4.4.5 IL-1 stim ulation ... 96
4.4.6 Multiplex measurement of protein concentrations inside hydrogel and in culture medium of 3D epithelial/stromal co-culture ... 96
4.4.7 Cytokine Luminex Assay...97
4.4.8 Dynamic correlation network generation...98
5 Conclusion ...
99
5.1 Sum m ary ... 99
F uture perspectives ... 101
List of Figures
Figure 1-1. Cell microenvironment interactions ... 19
Figure 2-1. Parameter space governing cell-matrix interactions in vasculogenesis ... 2 6 Figure 2-2. Workflow for determining microenvironment conditions for a given m orphogenesis assay ... 29
Figure 2-3. 2D attachment response to adhesion ligands ... 32
Figure 2-4. 3D endothelial structures in response to different adhesion ligands.. 33
Figure 2-5. 3D endothelial structures in response to changes in stoichiometric ratio "r"...3 5 Figure 2-6. 3D endothelial structures in response to crosslink MMP-degradability ... 3 7 Figure 2-7. Fibronectin-binding peptide effect on fibronectin accumulation...38
Figure 2-8. IPSC endothelial cells co-cultured with mesenchymal stem cells ... 40
Figure 2-9. 3D endothelial structures in response seeding density...42
Figure 2-10. Comparison between performance of IPSC endothelial cells and brain microvascular endothelial cells (hCMEC) with human brain vascular pericytes (H B V P ) ... 4 3 Figure 3-1. SrtA-mediated crosslinking of functionalized PEG macromers ... 54
Figure 3-2. Cytotoxicity study of MSCs exposed to SrtA crosslinking ... 55
Figure 3-3. Cell encapsulation strategy using SrtA-mediated crosslinking...56
Figure 3-4. Encapsulation of an endometrial stromal-epithelial co-culture ... 58
Figure 3-5. Encapsulation of primary rat hepatocytes...58
Figure 3-6. SrtA-mediated hydrogel dissolution and cell recovery...59
Figure 3-8. Epithelial acini passaged in 3D hydrogels...62 Figure 3-9. Recovery of large acini from endometrial epithelial-stromal co-culture
... 6 3 Figure 3-10. SrtA-mediated hydrogel dissolution kinetics...64 Figure 3-11. Dissolution of a wide variety of hydrogels under different conditions
... 6 6 Figure 4-1. SrtA incubation effect on intracellular kinase phosphorylation...81 Figure 4-2. Evaluating the effect of SrtA on soluble cytokines concentration,
compared to trypsin and Liberase, as evaluated with multiplex Luminex
tech n ology ... 8 2 Figure 4-3. Method for measuring and comparing local in-gel protein
concentrations to media concentrations using Luminex technology ... 84 Figure 4-4. Comparison of cytokine concentrations between culture media and local gel in response to an inflammatory cue for endometrial co-culture...85 Figure 4-5. Regression analysis of ratio of inside the gel and media concentrations as a function of cytokine molecular weight...86 Figure 4-6. Dynamic temporal response to IL-1b stimulation...88 Figure 4-7. Dynamic correlation networks ... 90
Chapter 1
1 Introduction
1.1 In vitro three-dimensional tissue models
While animal models have been routinely used in research and drug
discovery for decades, their limitations have become evident in the low success rate of clinical trials [1], underscoring some critical differences between animal models and humans. Beyond the lack of reliability in predicting efficacy, animal models pose considerable ethical considerations. Similar ethical challenges make it
impossible to use human subjects for early discovery and target identification, and although the use of human cadaveric tissue could offer some potential as an in vitro model, the limited availability and subject-specific background differences (such as disease or genetic variation) make this an impractical route. These challenges have motivated numerous efforts to develop in vitro models for testing drug efficacy and mechanisms of disease, aiming to produce systems with reproducible, quantitative, and reliable outputs for systematic screenings.
The most widely used in vitro model is the cell monolayer (e.g. caco-2 intestinal monolayer for drug transport studies). Although these 2D models have contributed immensely to the advancement of physiological models research and drug development, their significant limitations have motivated the development of three-dimensional tissue models. 3D models have the potential to capture relevant interactions that are often not present in 2D platforms. The transport properties of the system, the presentation of mechanical cues, and spatial distribution of cell populations are some key aspects of the cell-ECM interactions that can be better represented through a 3D model [2-91.
There are several reported cases where 3D formats have shown to behave more representatively compared to their 2D counterparts. In vitro liver tissue for example, which is of high interest for studies of drug metabolism and homeostasis
[10-18], has been shown to exhibit more tissue specific functionality in a 3D format compared to the 2D standard assay [19-251. As a specific example, albumin and
urea secretion, enzymatic activities, and long term viability were improved in a 3D model built by Miranda et a] [25] compared to a 2D standard model. 3D platforms can also better support complex morphogenesis of the cultured cells. Thomas et a] [261 introduced supporting stellate cells in a 3D hepatocyte model and showed the formation of bile canaliculi, desmosomes, and tight junctions in the 3D aggregates. These sophisticated features were accompanied by increased P450 enzymatic
activity in metabolizing testosterone. These examples highlight how 3D formats can be exploited to capture key physiological features.
Beyond recapitulating biologically relevant normal physiology, 3D models can be used to emulate disease to further understand its mechanism of action and test drug efficacy against it. For example, 3D breast cancer models have shown to capture certain aspects of tumor development that 2D platforms fail to show.
Mammary epithelial cells cultured in 3D matrices rich in basement membrane (BM) components formed polarized acinar structures, while culturing the cells similar gels lacking the BM proteins did not produce the same polarized acini [27].
Mammary carcinoma cells cultured in the same BM-rich matrices failed to form the spherical structures and continued to proliferate. While the 3D format distinguishes between the normal and diseased phenotypes, 2D assays do no discern between the morphology of the two cell types. 3D formats offer the potential of developing more
1.2 Synthetic biomaterials as a modular ECM
The increasing agreement over the advantages of 3D environment compared to standard 2D formats has prompted researchers to develop matrices that capture key features of the cell-ECM interactions [28-36]. A range of ECM materials has been devised, which can be broadly classified into naturally derived, and
synthetically designed.
Natural ECM models are derived from purified matrix components from animals or recombinant proteins, and reconstituted into a matrix. Examples of this type of ECM model include collagen I, fibrin, and Matrigel gels. Their natural source makes them attractive because of their intrinsic physiological relevance. Their composition varies from complex mixtures of proteins, like in the case of Matrigel, to purified or recombinant proteins. Matrigel for example, is derived from the basement membrane of mouse tumors and enriched with laminin [37]. It
consists of multiple ECM proteins (e.g. laminin, collagen, entactin) and an
undefined number and concentration of growth factors, which can vary significantly from batch to batch. While Matrigel has been useful in a number of studies serving as a matrix for epithelial cell types [38, 39], its batch to batch variability and
undefined nature makes it a less than ideal model for many studies. Fibrin gels have been widely used for vasculogenesis and angiogenesis applications [40-43], which they are very permissive for, but they suffer from extreme volume contraction after a few days of culture. Reconstituted collagen I gels have also been used by a number of researchers, but their heterogeneity and lack of modularity remain a significant limitation in designing meaningful and easily interpretable studies.
The need for modular ECM materials, where variables such as adhesion, stiffness, and permeability, can be independently tuned in a systematic and
reproducible way, has generated a number of synthetic scaffold types. Polyethylene glycol (PEG) is a prominent example of such materials. PEG is a bio-inert
hydrophilic polymer that is used in multiple applications in life sciences: drugs are sometimes "PEGylated" to extend their half-life, and materials that will come in contact with the body are coated with PEG to minimize protein adsorption, among many other examples. Multimeric PEG polymers can be cross-linked into an insoluble swollen network that can form a hydrogel. PEG hydrogels can be
crosslinked through a diverse number chemistries: Michael-type addition reactions between a thiol and an electron donor (acrylate, vinyl sulfone, or maleimide) [44], a click reaction (e.g. azide/norbornene, thiolene/norbornene, etc.) [45, 46], radical polymerization (UV, thermal, or chemically initiated) [471, or an enzyme-catalyzed reaction (e.g. Factor XIII crosslinking) [48]. A limitation of most of these
crosslinking schemes is that they are not compatible with the presence of serum, since the serum protein may interfere with the crosslinking process. The approach described in Chapter 3 here provides a potential solution to this problem.
Because PEG hydrogels are for the most part bio-inert (depending on the crosslinking procedure), it is necessary to introduce biological cues that recapitulate
relevant aspects of the ECM. In most cases it is essential to incorporate an adhesion ligand that engages cell surface adhesion receptors (e.g. integrins). Although the vast majority of the literature examples have used the minimal fibronectin-derived adhesion sequence Arg-Gly-Asp (RGD) [49-55], different cell types may need ligands targeting different integrins or with higher affinity than simple RGD. Dynamic remodeling and matrix degradation are other crucial aspects of the ECM that need to be incorporated into PEG hydrogels, especially for application that require cell migration. Because the mesh size of PEG hydrogels is in the nanometers range, and the cell size is in the micrometers range, cells require to break the hydrogel
crosslinks to spread and migrate through the gel. This has been achieved by
incorporating MMP-sensitive moieties as integral crosslinks of the gel. A limitation of PEG hydrogels had been that they impeded cell migration through them, but the development of new highly MMP-degradable sequences has much attenuated this problem [56, 57].
The modular nature of PEG hydrogels makes them a powerful platform to systematically study the interactions between the ECM and cells, and to design these interactions to foster a given phenotype. For example, engagement with
different integrins in a dose dependent manner can be screened independently from matrix stiffness and without significantly affecting the transport properties of the system. Such studies are not possible using naturally derived matrices, since variables such as ligand density, stiffness, and protein diffusivity are intrinsically coupled.
1.3 Key features of the cell microenvironment
The cell microenvironment has tremendous influence over the cell's phenotypic outcome (Fig 1.1, [31]). It drives tissue homeostasis, and its
dysregulation is known to lead to pathological conditions such as chronic
inflammation and cancer [58-62]. The local microenvironment comprises not only the complex interactions between the cell and the ECM, but also interactions between multiple cell types. The ECM has been traditionally known as the
structural support in tissues. Later it became clear that it also provided direct bidirectional signaling through engagement of cell surface receptors such as
integrins, and cell-mediated matrix remodeling. There is now growing awareness of the role that the ECM plays in modulating extracellular signaling networks by serving as a reservoir and in some cases direct effector for growth factors and cytokines. In fact, in some cases, cytokine interactions with the ECM are a requirement for the molecule's biological activity, like it is the case with basic fibroblast growth factor (bFGF). bFGF interacts with glycosaminoglycans synergistically to activate its receptor (FGFR)[63-72]. The ECM therefore has profound effect in both autocrine and paracrine signaling, if not by directly
activating or inhibiting the soluble signaling molecules, by serving as a reservoir for them and therefore spatially modulating their signaling dynamics. Diverse cell types often influence each other through both paracrine signaling networks and
direct cell-cell contacts through cell surface receptor signaling. For example, cytokine signaling interactions between stromal cells and epithelial tissues are known to play a strong role in cancer development and drug resistance [60, 73-76]. Studying these interactions in 3D formats can prove a challenging task.
Physical signals " Fibronectin " Vitronectin -Laminin " Collagen " Fibrillin " GAGs. PGs Cell fate processes 1 Replication 2 Differentiation 3 Migration Solut * Gro *Cyto -Che N ~ Ras PLC P13K Signal IN transducl --transcript on Cell-cell interactions Gene 'li{rI enreso Cadherins expression -CAM's ble signals with factors kines mokines 4 Apoptosis
Figure 1-1. (Figure and description adapted from [31]). Cell microenvironment interactions.
The ECM provides physical signals that interact with the cell bi-directionally, and it also serves as a reservoir for cytokines and growth factors, in some cases acting as an activating or inhibiting agent for these molecules. The microenvironment also includes cell-cell
interactions, through paracrine soluble signals and also through cell surface receptor interactions. These cues affect cell gene expression and protein transcription to ultimately determine cell phenotypes like replication, differentiation, cell migration, and apoptosis. Ellipsis (...) indicate that the list of signals is not intended to be complete.
19
I
4
Employing a 3D matrix for in vitro cellular assays offers the benefit of
capturing the microenvironment effects more faithfully. It also, however, introduces challenges in accurately characterizing the local molecular interactions and
recovering the cells from the matrix in a way that does not impact or alter their current state significantly. Naturally derived matrices such as Matrigel and collagen I gels can be degraded using proteases like collagenase and trypsin, but this process is slow, often incomplete, and most importantly, leads to degradation of the potential analytes of interest. Exposure to such promiscuous proteases can also negatively impact the cultured cells, making the interpretation of any further assays done on them challenging (e.g. FACS or western blots). Because of their modularity, synthetic matrices are more amenable to be dissolved for cell recovery in a bio-orthogonal way. Although there have been efforts in developing methods for recovering cells from some types of synthetic matrices, there is still the need for a method that does not alter the soluble molecules and cells in significant ways. Thermal [77], chemical [78], ionic [79] shifts, proteolytic degradation [80] have all been deployed to release cells, as has photo-degradation [81, 82], but these
approaches are either relatively slow, have variable success in minimizing cell damage, or are limited in application to relatively thin tissues. In a recent study, 3D-cultured human embryonic stem cell derived pancreatic cells were recovered from gels using Liberase as a gel dissolution agent [80]. Liberase is a mixture of collagenases and other residual enzymes. While this method indeed allowed them to recover the cell aggregates from the hydrogel, it would not be fitted to characterize soluble molecules, due to the promiscuous proteolytic activity of the enzymes included in Liberase. In this thesis we describe a novel bio-orthogonal method for recovering both intact cell aggregates and single cells as well as cytokines and growth factors without significantly depleting them.
1.4 Thesis objectives
The efforts in this thesis are focused around exploiting novel and cutting-edge biomaterials expertise to develop a platform that empowers researchers to gain biological insights from in vitro culture systems. Chapter 2 focuses on
understanding the ECM-cell interactions governing synthetic PEG 3D cell cultures, using vasculogenesis as a case study to define a relevant parameter space. Here, we explore the balance between cell adhesion and matrix degradation and study their impact in the cells' ability to form microvascular interconnected structures.
Incorporating more sophisticated adhesion ligands than simple RGD as well as more highly degradable peptide crosslinkers should lead to microenvironment
conditions permissive of vasculogenesis. Geometrical effects can also create artifacts in the observed cell response, so those are carefully considered here.
We also developed a novel crosslinking mechanism by using mutants of the enzyme sortase A to provide a PEG gel formation method that can be used in the presence of serum, which is particularly crucial for delicate primary cell types. We compared sortase-mediated crosslinking to other standard crosslinking chemistry and to a naturally derived ECM to assess its viability as a gel formation
mechanism. To provide evidence of its robustness and versatility, we deployed sortase crosslinking in two in vitro models: a human endometrial stromal/epithelial co-culture (using primary cells for the stromal component) and a primary rat
hepatocyte culture. Microscopy and immunostaining were used to assess
morphological features of the cultures, and different characteristic functional assays were used to further demonstrate the utility of these novel hydrogels.
A central objective of this thesis was to adapt PEG synthetic hydrogels to allow for accurate measurements of the local microenvironment. Here, we
repurposed the coupling enzyme sortase A to dissolve the synthetic hydrogels in a rapid, easy, and bio-orthogonal manner to be able to recover cells. Because of the
specificity of sortase, the dissolution process did not cleave cell-cell contacts,
preserving multicellular morphological features that a promiscuous protease would ablate. The same complex hydrogel cultures formed by sortase crosslinking were also dissolved by the addition of sortase and a small soluble GGG peptide, and the cultured cells were recovered and replated without disturbing their cell-cell
contacts.
Finally, the dissolution method was exploited to interrogate the local concentration of cytokines in the dissolve solution. This method led to the finding that the local cytokine concentration differs significantly from the concentration in the culture media across multiple analytes, both basally and in the context of an inflammatory response. Furthermore, the method increased the temporal
sensitivity to secreted cytokines in response to a strong inflammatory stimulus, and even revealed behaviors that were not captured by monitoring the culture medium concentration. We explored the implications of the local measurements in
developing cytokine correlation networks and compared the networks generated by using the in-gel concentrations to those generated by using the measurements of the culture medium concentration. The overarching goal of this thesis was to develop a platform that can aid in the early identification of mechanisms of disease and drug development and to eventually increase the success rate of clinical studies by improving the pre-clinical screening process.
Chapter 2
2 Developing a workflow to promote a desired morphogenesis
response: vasculogenesis case study
Contributions: Alexander Brown contributed with the data comparing IPS-derived endothelial cells and brain microvascular endothelial cells. Jeremy Velazquez assisted with experiments characterizing the IPS endothelial cells in a bioreactor platform in response to flow that are included in Blair et a] [831.
2.1 Introduction
Functionalized PEG hydrogels are widely used for in vitro tissue engineering, but the majority of studies have focused on a relatively limited range biological functionalities, with phenotypic outcomes typically examined in a univariate and empirical fashion. For example, the RGD peptide motif from the 10th Type III repeat in FN is ubiquitously and arguably indiscriminately employed to induce cell
interactions with synthetic matrix, perhaps because many studies employ cell lines inadvertently selected to adhere to vitronectin via av63. Further, the concentration of peptide is often very high to compensate for low affinity binding to cells [84].
Cellular remodeling by proteinases or other enzymes is also a common approach, but few studies have parsed how crosslink density and enzyme-substrate catalytic
parameters independently contribute to the rate of remodeling and subsequent phenotypic outcomes. It is has been reported in standard (2D) culture systems that the nature of cell adhesion can influence subsequent proteinase activity, in part through activation of intracellular signaling pathways [85-90], but how the cellular adhesion status in 3D environments influences proteolytic activity is not well understood.
The biophysics of migration and adhesion have been extensively studied in 2D platforms. For instance, mechanistic models describe the biphasic migration behavior in response to adhesion ligand density, where maximum migration speed occurs at an intermediate adhesion strength relative to intracellular contractile force, at which cells can form attachments in the leading end, but can also detach at the rear end [91-93]. Other studies have extended these models to incorporate an appreciation of the effect of the biophysical presentation of the adhesion ligand on cell response. Adhesion ligand clustering, for example, can facilitate integrin clustering enabling assembly of multiple cytoplasmic proteins and triggering
certain intracellular signaling pathways [94, 95]. For instance, single cell migration speed and adhesion were increased with increased ligand clustering [94].
Interestingly, Maheshwari at a] did not observe the expected biphasic behavior as a function of adhesion ligands density, but this is likely due to the fact that they used a low affinity peptide sequence: YGRGD. There have been attempts to extend the principles of cell migrations observed in 2D to 3D matrices through systematic studies [96, 97]. Zaman et a] demonstrate that 3D environments introduce additional governing forces in cell migration -namely, the balance between cell-matrix adhesiveness and cell-matrix stiffness, and between traction force and cell-matrix sterics [96]. A significant conclusion derived from these studies is that cell
migration was increased in less dense, softer gels. Although these studies used Matrigel, which introduces an undefined amount of growth factors and other proteins, we focused on using low crosslinked PEG gels as a starting point for our studies.
The objective of this work is to incorporate an appreciation for known biophysical principles that govern morphogenic behaviors into a semi-empirical approach for efficiently defining synthetic extracellular matrix conditions to achieve a particular morphogenic event (here, vasculogenesis). A semi-empirical approach is desirable because both gel and cell properties dictate the response, but the cellular parameters governing response to defined gel conditions - e.g., absolute or relative
integrin expression levels, proteinase activity levels, cellular contractility potential, etc. -- are typically not known a priori and are typically prohibitively resource-intensive to measure. For example, an elegant study in a 2D model system used independent tuning of cellular integrin expression, adhesion ligand density, and integrin-ligand bond affinity to demonstrate that for any combination of
parameters, the resulting total number of substrate bonds, and resulting cell-substrate adhesion strength, dictated the cell migration speed [91]. In other studies, migration speed was further tuned by altering cellular contractility with growth factors [94, 98-100]. It is not practical to control or even know the cellular
parameters involved in most tissue engineering applications. However, appreciation
of how they might change among cell types, and that the cognate gel parameters
can systematically be varied to capture true independent variables like cell-matrix bond number, allows one to develop a workflow plan that will allow systematic and efficient identification of gel compositions and stimulus for any particular
morphogenesis event. The approach described here combines high throughput 2D screens with more resource-intensive 3D screens. As a case study for developing this workflow, the important phenomenon of in vitro vasculogenesis is analyzed, using an induced pluripotent stem cell (IPSC)-derived endothelial cell line.
Vasculogenesis is the de novo formation of microvessels from endothelial cells or endothelial cell progenitors dispersed in a gel. This process has been extensively studied using various natural protein matrices (e.g. collagen, fibrin) [43, 101-105], however, efforts to effect vasculogenesis in synthetic hydrogels and to understand how gel parameters influence the process for specific endothelial cell types are
scarcer. The modular nature of synthetic hydrogels offers an attractive means to independently tune key parameters to interrogate biological response to different microenvironment conditions [48, 106].
The endothelial cell vasculogenic response is a particularly interesting study for this purpose due to the vast heterogeneity in endothelial cell phenotypes from
different sources. For example, HUVECs, one of the most widely used endothelial cell type in the tissue engineering literature, lack the crucially important
inflammation response receptors CXCR1-3 that are characteristic of microvascular endothelial cells [107]. Anecdotally, experiments in the Griffith lab have revealed vastly different vasculogenic behaviors using endothelial cells of different origin. The focus here is on IPSCs, as they represent a well-defined primary source with potential for adapting to host tissue [108- 110]. Due to its potential to exhibit tissue
specificity, we chose to use an IPSC-derived endothelial cell line recently shown to be a promising endothelial cell model [83].
A
B
Cell-cell contacts (seeding density)
Adhesion environment
e/ 0' Biophysics: clustering, tether ligand
- Matrix accumulation
O -~ Ligand identity (RGD, Syn-K-RGD, GFOGER) v03 a501 a2p1
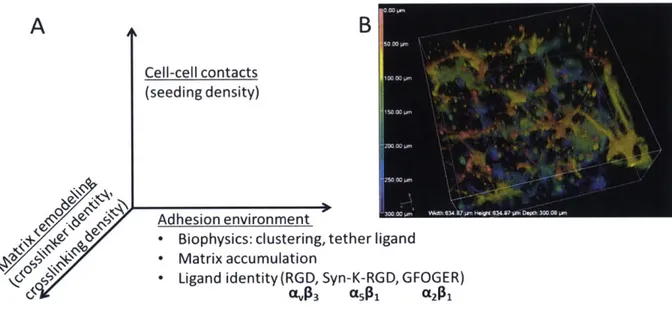
Figure 2-1. Parameter space governing cell-matrix interactions in vasculogenesis. (A) Parameter space showing axes of matrix adhesion, mechanics, and cell-cell interactions. (B)
3D
reconstruction of a 300 tLm stack of endothelial networks, color coded to indicatez-position.
Here, the parameter space capturing crucial interactions between synthetic ECM and endothelial cells is organized into a 3D space illustrated in Fig 2-1A. The adhesion interactions are governed first by the identity and average concentration of integrin ligand, where small peptide ligands targeting integrins the
pro-angiogenic integrins C5631 [111-113], CA63 [114, 115], and a261 [116] are employed. Although integrins G5B1 and CL2B have been clearly shown to be pro-angiogenic, aV63 has been proposed to be anti-angiogenic when interacting with certain ligands
[117]. We included the av83 ligand RGD in our studies both as a benchmark, given its widespread use in the literature and its at least low affinity interactions with several integrins. Cells, however, secrete their own ECM, which may accumulate in the local environment and further influence cell adhesion responses; this matrix accumulation can be influenced by the presence of moieties that bind to the secreted
matrix, representing another variable on the adhesion axis. A further means to control the biophysics of adhesion involves local clustering and local mobility of the ligands on tethers; this level of control is not explored in these studies and these parameters are kept constant [118, 119]. Along the degradation speed axis, the both crosslink density and proteinase substrate catalytic parameters influence
degradation, for a fixed set of adhesion parameters. Finally, the cell seeding density axis incorporates several independent phenomena - distant cells must stretch to make contacts, accumulation of autocrine and paracrine factors, and local durotaxis microphenomena from neighboring cells.
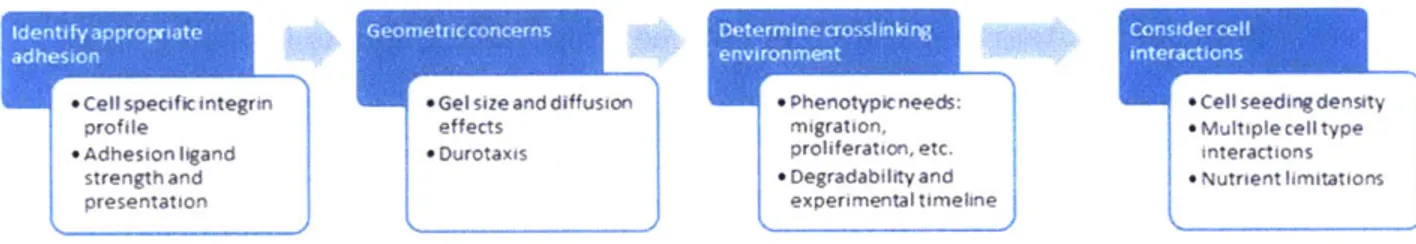
By independently varying these parameters, combinations of conditions that foster robust formation of microvessel networks by iPS-derived endothelial cells were identified (Fig 2-1B). The workflow used to define microenvironment conditions for a given morphogenesis application (Fig 2-2) includes:
i) 2D short term attachment screen to identify appropriate adhesion ligand for cell interactions; the appropriate ligand density in 2D and 3D may be different, but the 2D screen of attachment is a surrogate for the integrated effects of ligand bond affinity and integrin expression level.
ii) Define desired gel geometry and identify conditions that enable isotropic swelling following crosslinking. Anisotropic swelling (e.g. due to bonding to a
substrate of confinement in one dimension) can create artefactual durotaxis forces and inhomogeneous phenotypic behaviors throughout the gel. Durotaxis may be useful in some contexts, but the baseline properties of the gel are a starting point for vasculogenesis potential.
iii) Screening of crosslinking conditions. The crosslinking density and degradability of the system can be tailored depending on factors such as the timeframe of the experiment and the phenotype of interest. For example, a multi-weeks experiment may require a crosslinker of relatively low catalytic efficiency to avoid full degradation of the gel during the experiment, and a phenotype requiring extensive cell migration will likely need low crosslinking and/or fast degradation. A list of peptides commonly used as crosslinkers can be found in these studies [56, 57]. Here, we repurposed a peptide sequence originally used as a FRET probe due to its extremely high reported catalytic efficiency for MMPs 2, 9, 13, and 14, among other proteases [120]. MMP-2, for example, has been reported to be involved in
vasculogenesis [121-123]. The number of crosslinks in the gel can be modulated by changing the polymer weight percent of the gel while keeping the crosslinker:PEG ratio constant, or alternatively, by adjusting the stoichiometric ratio of
crosslinker:PEG while keeping the PEG weight percent constant, as done here [54]. We chose the latter approach due to the fact that changing the total PEG
concentration can introduce significant changes in properties such as
macromolecule diffusivity through the gel. Additionally, maintaining a fixed concentration of macromer while decreasing the crosslinker concentration leaves available functional groups that can be used to introduce different bio-cues, such as adhesion peptides and growth factors.
iv) Consider if cell-cell interactions or interactions between multiple cell types are crucial for the phenotype of interest. Cell-cell interactions can be
influenced through the cell seeding density. For example, sparsely seeded cells will need to degrade and migrate significantly to establish cell-cell contacts, requiring
relatively high seeding densities, but low seeding density may be more appropriate to study single cell migration. The presence of multiple cell types can be an
important factor to consider. For instance in the context of vasculogenesis, stromal cells and pericytes contribute to the formation and stabilization of the endothelial
microvessels.
Finally, cells produce and assemble their own ECM molecules, hence the synthetic adhesion ligands in the initial gel may function in a transient manner if the secreted matrix molecules accumulate and are remodeled into a mature natural cell-secreted ECM. The nature of matrix accumulation can be influenced if peptides that bind to secrete matrix are included in the gel. Thus, an additional variable in parameter space is the time-dependent matrix stabilization.
* cespecif o integr n eGelsizeand dtffusion + ipneeds: *c'l seecog 'esi'y
cfde effects Migration. * Muliple cell ty)e
' --neston igand * Durotaxis orohferaticn, etc. mteracTions
rengtn and * Degradabdity and * Nutrient hoitations
oresentation experime-ntal timehne
Figure 2-2. Workflow for determining microenvironment conditions for a given morphogenesis assay
2.2 Results
2.2.1 IPSC-derived endothelial cells as a cell model for vasculogenesis
IPSCs were acquired from Cellular Dynamics International (CDI) in a differentiated state toward endothelial lineage. CDI characterizes the cell line as endothelial based on a set of characteristic markers (CD31, von Wilebrand factor, tight junction protein ZO-1) and the cells ability to demonstrate barrier functions. Additionally, a recent study [83] showed evidence that this cell line exhibited characteristic endothelial functions: proliferation in response to VEGF,
upregulation of MCAM and ICAM1 in response to TNF-a, thrombin dependent barrier function, lumen formation in natural matrices, and the common capillary formation on Matrigel. Furthermore, the IPSC endothelial cells form complex structures and align in the direction of flow in a 3D Matrigel gel in a bioreactor when visualized through fluorescence microscopy. This IPS-derived cells have also been used to repopulate decellularized matrix with reasonable success [124]. The studies referenced here and our experience suggest that this cell line is a suitable model for endothelial morphogenesis studies.
2.2.1.1 2D attachment assay reveals differential attachment to RGD, SYNKRGD, and GFOGER for human IPS-derived endothelial cells
Although the IPSCs used here have shown capillary structures in natural matrices [831, their integrin profile has not been reported. Ligands for integrins
a5B1, Qv63, and a261 were screened for 2D attachment due to the reported role of
these integrins in angiogenesis and vasculogenesis [111-116]: minimalist RGD
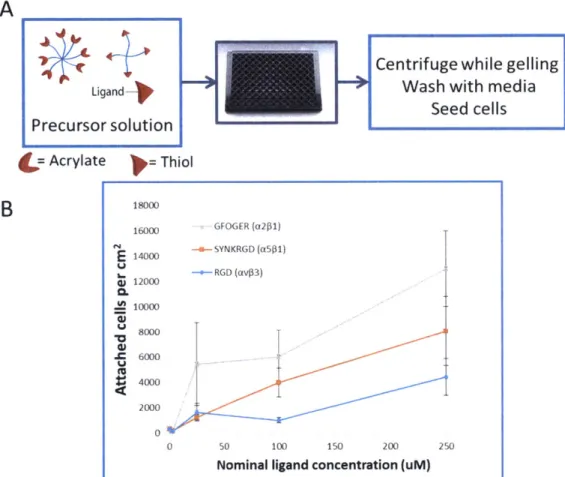
(MV63), SYNKRGD ([561), and GFOGER (Q261) (see methods for full sequences). The ligands were presented on a flat PEG hydrogel surface at different concentrations (Fig 2-3A). The gel swelling behavior did not seem to introduce local attachment differences in the format and timespan of the experiment. This could suggest that the gel swelled isotropically in the z direction (upwards) while being constrained at the bottom and sides.
RGD had the lowest attachment response, while GFOGER elicited the highest attachment across concentrations from the ligands tested (Fig 2-3B). SYNKRGD promoted higher attachment response than the minimal RGD, but not as high as GFOGER. Higher attachment response to GFOGER could be attributed to either the cells expressing integrins with higher affinity to peptide (a21), or a higher number integrins for that particular peptide relative to other integrins. The three ligands exhibit different affinity for different integrins. A minimal RGD
sequence is a promiscuous but weak ligand for a number of integrins, but it mostly engages av63 [125-127]. SYNKRGD presents, in a branched configuration with RGD, the synergy site derived from fibronectin type III domain 9: PHSRN. The
combination of these two motifs along with the proper spacing is known to foster interactions more strongly with the canonical fibronectin integrin a.s1 [128-132]. The GFOGER peptide is presented as a triple helix formed by the flanking repeats
in GGYGGGPG(GPP)5GFOGER(GPP)GPC, and it is known to interact with the
canonical collagen I receptor a261 [133-135]. It is possible that the IPSCs have an integrin profile for which GFOGER is more favorable, but we must also consider differences in the biophysical presentation of this peptide compared to the RGD variants. The GFOGER peptide self-assembles in trimers. It is known that adhesive ligands that are presented in a clustered manner elicit stronger adhesion and
increased migration compared to ligands presented at a similar density but in a randomly distributed manner [94, 136-138]. GFOGER timers could be engaging integrins as a ligand cluster, therefore facilitating the formation of focal adhesions, although it is also possible that the close spacing between the GFOGER individual peptides is not optimal for integrin clustering. It is also known that hydrophobic peptides can non-specifically bind proteins. It is conceivable that the GPP repeats non-specifically bind adhesion proteins from the serum or secreted by the cells and facilitate cell binding and spreading. Mechanistic studies with specific integrin inhibitors or variants of the GFOGER peptide would be required to elucidate the reason for GFOGER to promote highest attachment of the IPSC endothelial cells.
A
B
Ligand Precursor solution = Acrylate Thiol 18000 16000 GFOGER (12161) 14000 SYNKR()D (3501) - - RGD (av$3) q 12000 c. Q-" 1,tCentrifuge while gelling Wash with media
Seed cells
0 -.
0 50 100 150 2C0 250
Nominal ligand concentration (uM)
Figure 2-3. 2D attachment response to adhesion ligands. (A) 2D attachment assay diagram. (B) Attachment response to PEG surfaces presenting three different ligands: RGD, SYNKRGD, and GFOGER. Each data point represents the average of 3 wells. Error bars represent SEM.
2.2.1.2 Increased 2D attachment correlates with more complex vascular 3D
networks of hIPSECs
IPS endothelial cells were encapsulated in PEG hydrogels presenting three
different ligands (2 mM, nominal): minimal RGD, SYNKRGD, and GFOGER (full
sequences in the methods section). We hypothesized that higher 2D attachment
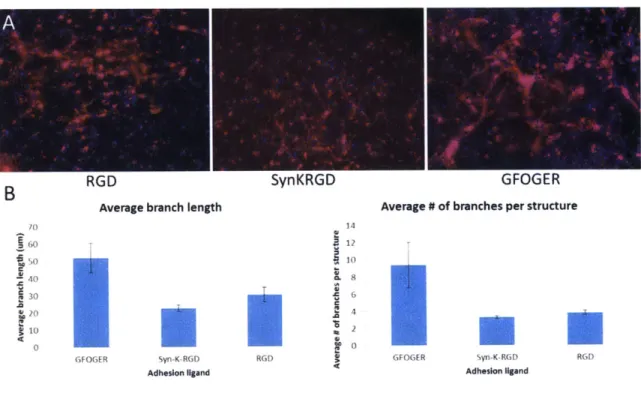
would result in more complex network formation in 3D. As we expected, cells encapsulated in hydrogels presenting GFOGER formed more complex networks
2-4). The average branch length and the average number of branches per structures were ~2.5-fold higher in GFOGER compared to RGD and SYNKRGD. Surprisingly, these parameters were not significantly different in RGD and SYNKRGD gels. All other gel parameters, such as crosslinker degradability and crosslinking density, were constant across conditions at a relatively low crosslinker:norbornene
stoichiometric ratio r=0.58. As discussed above, GFOGER presents potential differences in how the cells interact with the hydrogels compared to the RGD
variants (more total peptide could contribute to non-specific protein adsorption and clustering potential), and the 3D format could enhance some of these effects.
Cell-secreted matrix deposition could be affected by non-specific interactions with the large coil of the peptide. Although experiments in 2D have shown that the coil section does not promote attachment in the absence of the GFOGER motif [133], this could not hold true when translating the system to 3D since diffusion of cell secreted proteins could be diminished.
A
B RGD SynKRGD GFOGER
Average branch length Average # of branches per structure
Adhesion ligand Adhesion ligand
Figure 2-4. 3D endothelial structures in response to different adhesion ligands. (A) Micrographs of encapsulated IPSC endothelial cells at day 5. Red= phalloidin, blue=
DAPI. (B) Quantification of network parameters. Error bars represent SEM, n=10-20 fields of view.
2.2.1.3 Hydrogel MMP-sensitivity and crosslinker:macromer stoichiometric ratio affect hIPSECs network formation
Because the nominal mesh size of PEG hydrogels is in the order of
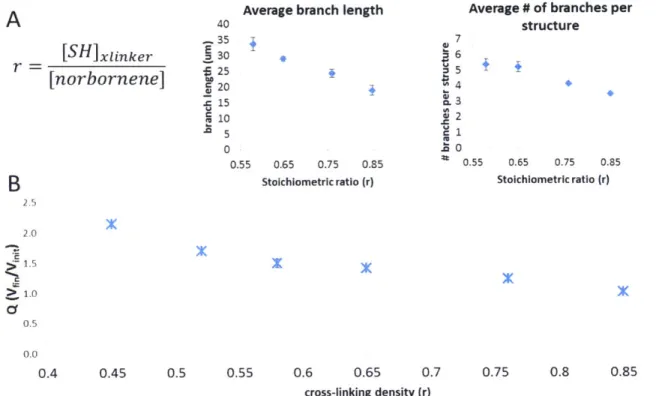
nanometers, single cells need to degrade the hydrogel crosslinks in order to change shape and migrate to find other cells. We modulated the stoichiometric ratio (r) of the crosslinking thiols to the PEG norbornene groups and measured its effect on the network parameters, while keeping the adhesion ligand concentration constant (2 mM nominal GFOGER) and a constant relatively high nominal seeding density (7.5x106 cells/mL) to enhance cell-cell contacts. We observed that when we
decreased r from 0.85 to 0.58, the average branch length and the average number of branches increased (Fig 2-5A), likely because the cells need to degrade fewer
crosslinks in order to change morphology and migrate to find each other and form interconnected networks. Hydrogels with r < 0.5 showed very robust network formation at early time points (day 1), but broke down before day 5. It is worth noticing that this hydrogels were formed within a silicone ring in 96-well plates and it is possible that the hydrophobic initiator (IRGACURE 2959) partially adsorbed to it, leading to the real extent of crosslinking being significantly different from the intended nominal one. A gel formation method that does not come in contact with a hydrophobic surface could likely generate hydrogels of lower nominal r that are stable during a 5 day culture.
While there has been increasing interest in the effect of matrix stiffness in different cell phenotypes such differentiation and migration [96, 139-142], we hypothesis that the effect observed here is likely related to the number of peptide crosslinks that the cells are required to degrade in order to migrate, rather than the stiffness differences that arise from changing r.
Average branch length Average # of branches per
A
40 structure -.,35 (U7 [SH ]xIinker. E 30 - 6 r 25 5 [norbornene] U 2 15 3 10 2 5 1 0 . U 0.55 0.65 0.75 0.35 0.55 0.65 0.75 0.85B
Stoichiometric ratio (r) Stoichiometric ratio (r)2.0 > 1. 0.5 0.0 0.4 0.45 0.5 0.55 0.6 0.65 0.7 0.75 0.8 0.85 0.9 cross-linking density (r)
Figure 2-5. 3D endothelial structures in response to changes in stoichiometric ratio "r". (A)
Quantification of network parameters as a function of stoichiometric ratio "r". Error bars represent SEM. n=40- 100 fields of view (B). Swelling behavior of PEG gels as a function of stoichiometric ratio "r". Error bars represent SEM. n=3 gels
Altering r also has an effect on the swelling of the hydrogel (Fig 2-5B), and therefore affects variables such as effective ligand concentration, seeding density,
and protein diffusivity through the gel. For example, a more loosely crosslinked hydrogel will exhibit a higher degree of swelling and will effectively dilute the nominal adhesive ligand concentration, as well as the original seeding density. We therefore investigated changing the degradability of the peptide crosslinker, rather than changing the number of crosslinks through r. The adhesion ligand was fixed at a nominal 2 mM GFOGER, and the seeding density was kept at a relatively high 7.5x106 cells/mL. There are a number of reported MMP-degradable sequences [56]. The GPQGIWGQ degradable peptide (dp) is one of the most commonly used. To assess the effect of the crosslinker identity, we identified a peptide sequence with extremely high reported catalytic efficiency for MMPs 2, 9, 13, and 14, among others
(Fig 2-6C), and adapted it as a hydrogel crosslinker by introducing flanking cysteines. By comparing the conventional crosslinking MMP-degradable peptide (dp) to the more highly degradable one (h-dp, full sequence in methods) (Fig 2-6C), we see that increased MMP sensitivity in the hydrogel yields to more complex
networks (higher average branch length and number of branches per structure) (Fig 2-6A-B). It is worth noting that the catalytic efficiencies reported in Fig 2-6C have been taken from different studies [56, 120]. Measuring both peptides' kcat/Km in the same study using a fluorescamine assay [143] or other methods would strengthen the interpretation of these data. It is also possible that the crosslinking sequences differentially interact with cell proteases in an inhibitory manner, by competing with targets on the cell surface, but this hypothesis is not investigated in this thesis. Altogether, these results suggest that IPSC endothelial cell network formation process relies on the ability of the cells to degrade through the PEG
hydrogel crosslink sites, and that the process can be modulated either by changing the number of crosslinks, or by changing the MMP-sensitivity of the crosslinks.
A
conventional (dp)highlv degradable (h-dp)
B Crosslinker Identity Effect
Average branch length Average # of branches per structure
C
C
dp h-dp dp h-do
crosshnker crosshnker crossnnker cross nker
C
cleavage catalytic efficiencyFigure 2-6. 3D endothelial structures in response to crosslink MMP-degradability. (A) Micrographs of encapsulated IPSC endothelial cells at day 5 in hydrogels crosslinked with
either a conventional degradable peptide (dp) or a highly degradable peptide (h-dp). Red= phalloidin, blue= DAPI. (B) Quantification of network parameters for cells encapsulated
through the dp and h-dp crosslinker. Error bars represent SEM. n= 40-60 fields of view. (C) Reported catalytic efficiencies for dp and h-dp peptides for different MMPs and ADAMs [56, 120].
2.2.1.4 Fibronectin-binding peptide increases fibronectin accumulation by IPSC-derived endothelial cells
Because of the tremendous complexity of the extracellular matrix -proteins are secreted and assembled by the cells, exposing cryptic sites upon mechanical strain, and providing signals back to the cells- we hypothesized that a single adhesive molecule, such as GFOGER or RGD, cannot capture all features of the ECM. Rather, the hydrogel must allow cells to secrete and assemble their own microenvironment. Our strategy to promote cell-driven ECM assembly is to
covalently incorporate a matrix-binding peptide in our hydrogel, to promote matrix stabilization. The adhesion ligand used in this experiment was RGD, with the rationale that phenotypic differences caused by FN accumulation would be more noticeable with the lowest attachment peptide. We chose a peptide with binding affinity to fibronectin (GCRE-TLQPVYEYMVGV) [144], because fibronectin
crosslinks other matrix proteins in early matrix assembly functioning as a scaffold for them. IPS endothelial cells were encapsulated using the lowest r (0.58) at a relatively high cell seeding density of 7.5x106 cells/mL to promote cell-cell contacts. We observe that structures formed in hydrogels that present a FN-binding peptide, in addition to an adhesion ligand, show localized staining in the cells vicinity at day 5, while the staining was not present in the absence of the FN-binding peptide (Fig 2-7B). Fibronectin, while being a large protein, is secreted in globular form, and cells then bind and unfold it [145-149]. We hypothesize that globular FN can diffuse
out of the hydrogel in the absence of the FN-biding peptide, but when the peptide is included, FN can be retained longer in the vicinity of cells to the cells can bind to it. Additionally, FN molecules can crosslink with each other so longer retention can promote FN-FN binding and stabilization [147, 148]. The impact of the matrix binder is likely to be dependent on the secretion rate of the ECM molecules.
Extensive secretion of FN, for example, could lead to FN-FN crosslinking and stabilization of other ECM molecules. Although we did not observe a significant difference in phenotype from ECM accumulation in the timeframe of this
experiment, it is possible that this could have an influence in longer term network stability [150, 151].
A
B
0 (9 fibronectinLE1I
f ibronectin + binding peptide 1Figure 2-7. Fibronectin-binding peptide effect on fibronectin accumulation. (A) Diagram of fibronectin secretion and stabilization through matrix-binding peptide functionalized to
PEG hydrogel. (B) Micrographs of encapsulated IPSC endothelial cells at day in the
presence (bottom) or absence (top) of fibronectin-binding peptide. Green= anti-fibronectin, red= phalloidin, blue= DAPI (B). Quantification of network parameters as a function of cell seeding density. Error bars represent SEM.
2.2.2 Cell-cell effects
2.2.2.1 Co-culture of hIPSECs with mesenchymal stem cells promotes complex structures
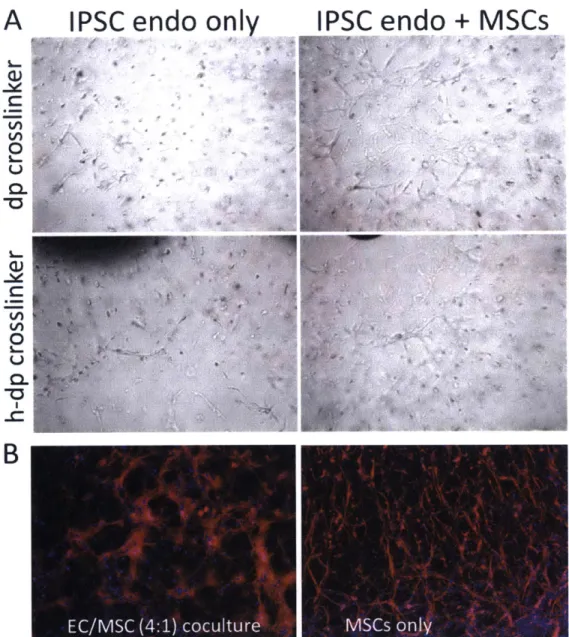
A number of mesenchymal cell types are known to stabilize and support the formation of vascular networks in vivo [43, 152-156]. We examined the effect of introducing mesenchymal stem cells in the structures formed by the IPSC endothelial cells in PEG-NB hydrogels. We encapsulated an IPSC:MSC (4:1) co-culture and observed the networks formed in comparison to the IPSC monoco-cultures, using the most permissive conditions discussed above: r=0.58 using h-dp or dp crosslinkers (the MSCs are more proliferative and could potentially degrade the entire gel more quickly than the IPSCs alone), 2 mM GFOGER, and 7.5x106
cells/mL .We observed that the presence of the MSCs enhanced network formation at earlier time points, with noticeable difference in the complexity of the formed networks by day 2 (Fig 2-8A). All conditions were cultured with the IPSC
endothelial cell media formulation described in the methods and recommended by CDI. Using the h-dp did not seem to visibly affect the network formation process in the presence of the MSCs. Because MSCs have been reported to form
interconnected structures on their own, especially in the presence of factors such as VEGF (present in the medium used here), we compared the structures formed by the MSCs alone to those formed by the co-cultures. Fig 2-8B shows z-projections of fixed and actin-stained micrographs (200 gm) of either MSC monoculture or
IPSC:MSC at day 5. Both conditions show interconnected structures, but they are markedly different. The IPSC:MSC co-cultures formed networks with thicker branches compared to the MSC cultures, where the structures were mostly formed by single cell-cell connections. Although the cells were seeded at the same seeding density (7.5x106 cells/mL), the fact that the two cell types exhibit different
proliferation rates makes interpretation of the results challenging. Nevertheless, we can capture emergent behaviors arising from multicellular interactions with our synthetic hydrogel system, such as robust thicker networks formed at earlier time points when having both cell types.
A
IPSC
endo only
V) 0 CL) W~
IPSC endo + MSCs
4 -f 4B
Figure 2-8. IPSC endothelial cells co-cultured with mesenchymal stem cells. (A)
Micrographs of encapsulated 1PSC endothelial cells (left column) or ISPC endothelial cells co-cultures with MSCs at a 4:1 IPSC:MSC ratio (right column) at day 2, in hydrogels
![Figure 1-1. (Figure and description adapted from [31]). Cell microenvironment interactions.](https://thumb-eu.123doks.com/thumbv2/123doknet/13804010.441293/19.918.94.794.253.809/figure-figure-description-adapted-cell-microenvironment-interactions.webp)