Modeling dermatophytosis in reconstructed human epidermis: A new tool to study infection mechanisms and to test antifungal agents
29
0
0
Texte intégral
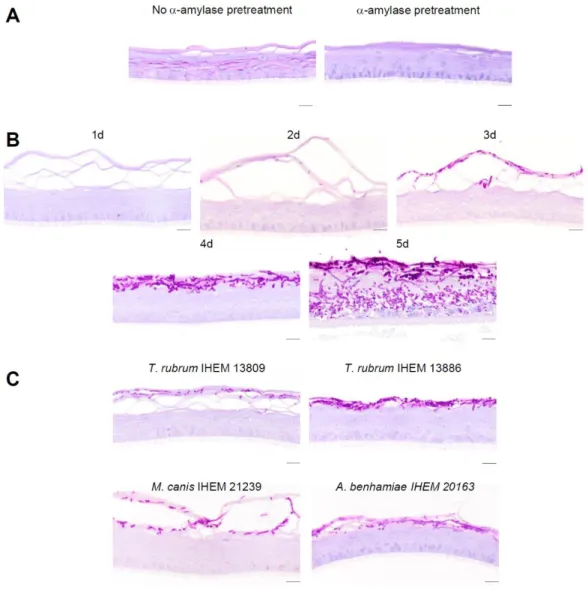
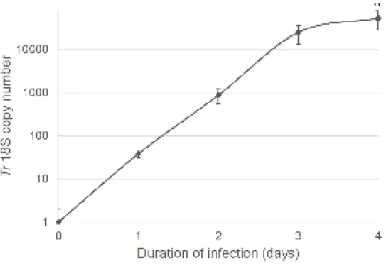
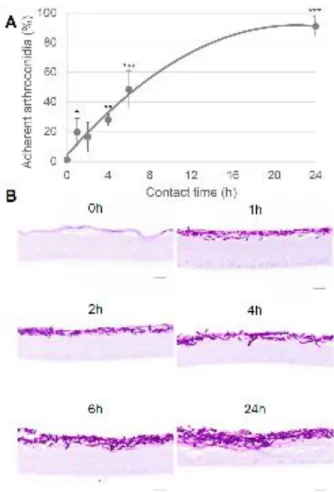
Figure

Documents relatifs
