Observation of extensive chromosome axis remodeling during the "Diffuse-Phase" of meiosis in large genome cereals
Texte intégral
Figure
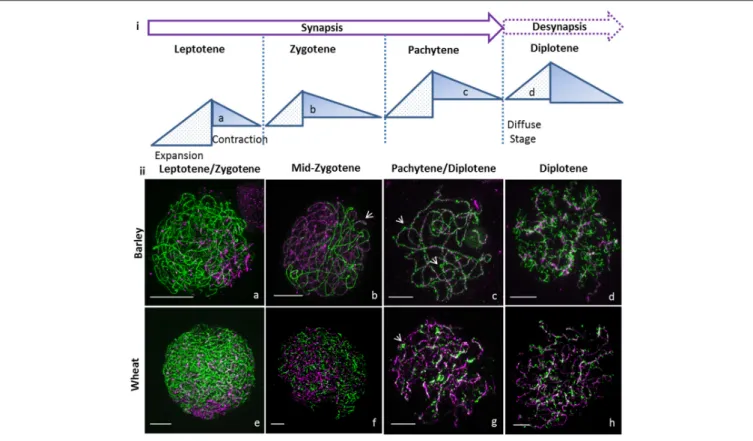
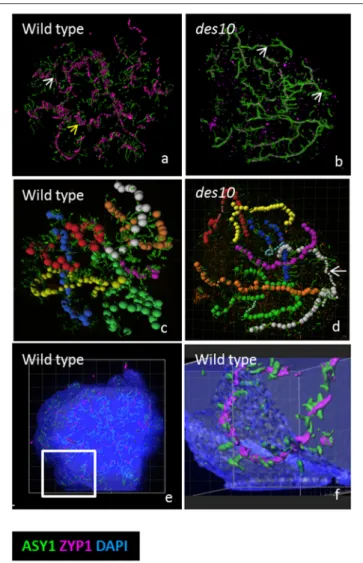
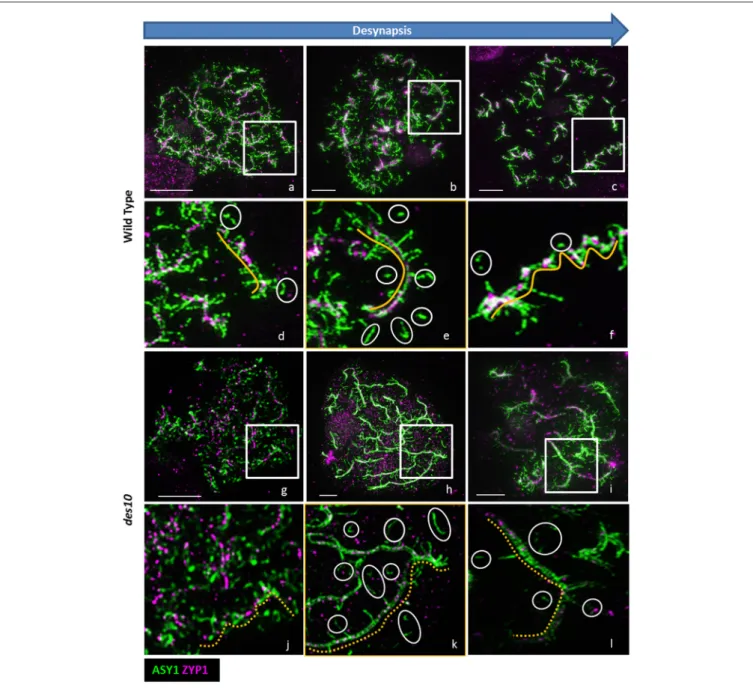
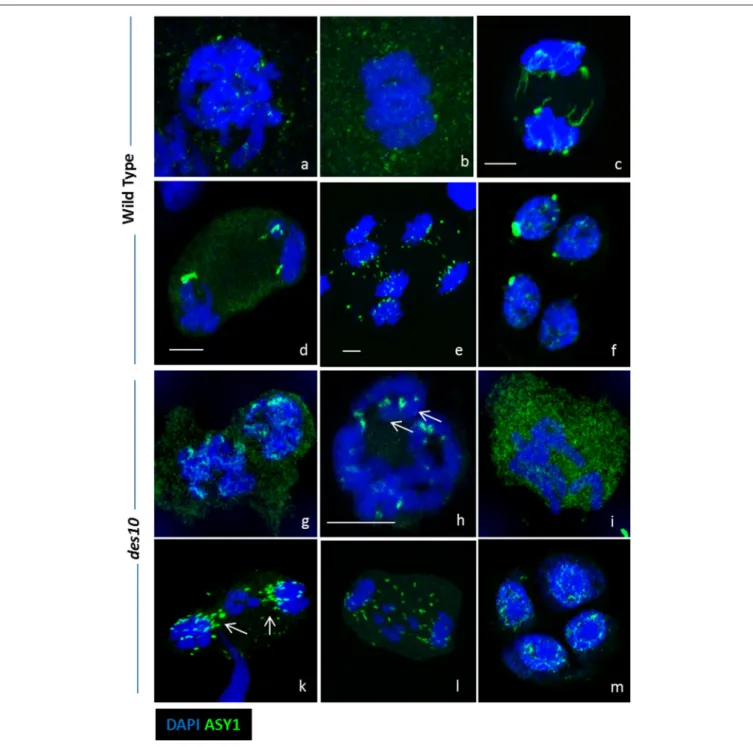
Documents relatifs
Our results (i) identify differentially expressed proteins showing correlated or uncoupled mRNA/protein profiles in mitosis and meiosis, (ii) suggest novel
- Les stomatites de contact allergique : Beaucoup plus fréquentes que les manifestations allergiques immédiates, les manifestations de l’hypersensibilité retardée de la muqueuse
While Theorem 6 applies to the ideal situation when at most one single change point occurs between time t 0 and time t, we now address the case when multiple (possibly
Based on the hypothesis that red meat has a greater carcinogenic potential than other types of muscle food, the main objective of the present study was to explore the
It is expected from the phase diagram (4) that the equilibrium A13Sc phase precipitates in this aging condition; however, the atom probe data show the Sc
Once again, this demonstrates the vast utility of using parametric systems rather than state spaces for interactive environments. Here we only need to change the sign
En effet, dans le contexte mondial actuel où les questions de réchauffement climatique, de déforestation et d’insécurité alimentaire prennent une place de plus en plus
Nous avons vérifié que l’alternance entre des régimes carencés en différents AAI n’af- fecte pas la masse maigre ; (3) il pré- sente une réversibilité totale et rapide de
