TtcA a new tRNA-thioltransferase with an Fe-S cluster
Texte intégral
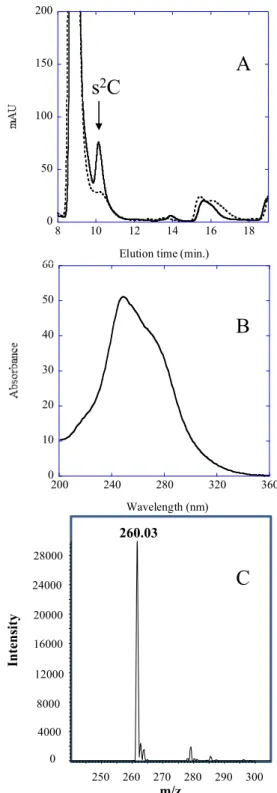
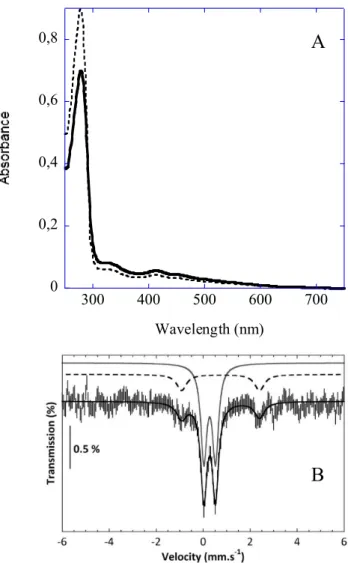
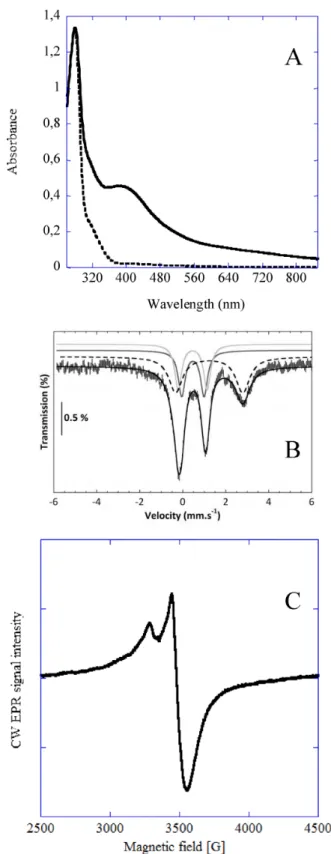
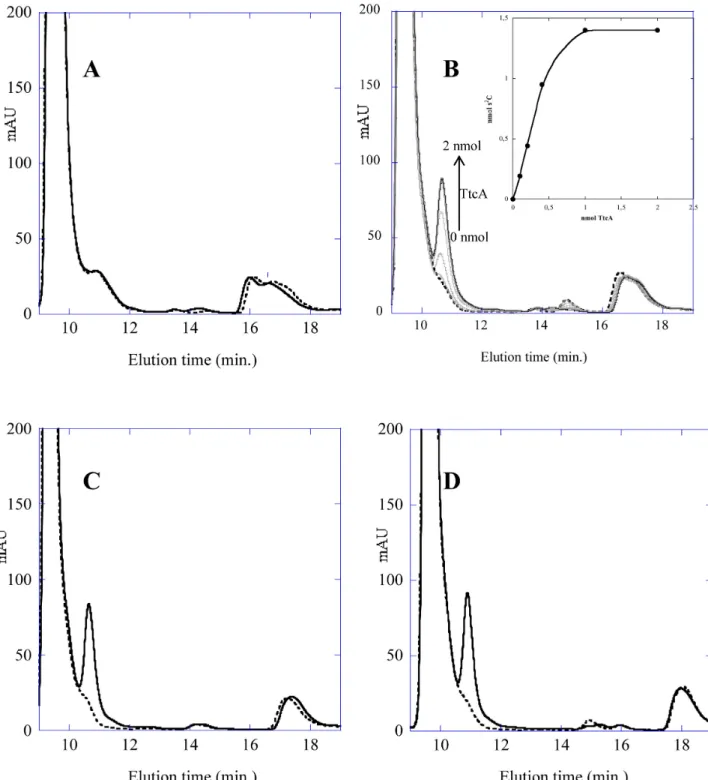
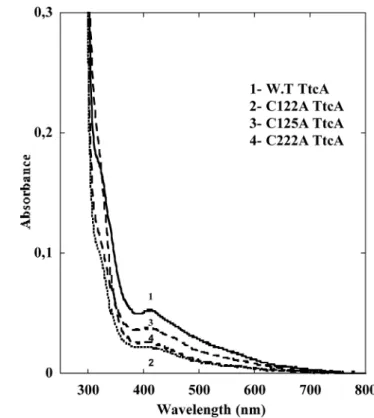
Figure
Documents relatifs
pathogenesis-related protein 1; GO: Gene ontology; HIF1A: Hypoxia-inducible factor 1-alpha; IL8: Interleukin 8; IL24: Interleukin 24; JUN: Jun proto-oncogene; JUNB: Transcription
Notre objectif direct est l’application de la boite à outils de mesure de la richesse WWTK (Weightless Wealth Tool Kit pour la mesure du capital intellectuel,
An extended lattice model for reaction limited cluster-cluster aggregation is studied, where different kinds and states of bonds between.. monomers
dispersion curves of ultrasonic guided waves. The analysis of the synthetic signals and experimental data 16. illustrates that the S-SVD method may provide significant
Phytoplankton community structure was dominated by the diatom Rhizosolenia hebetata in Kavala Gulf, whereas, a very intense bloom of Skeletonema costatum was observed in
Si une équipe reconstitue le modèle à trois étages, elle gagne 3 points, pour 4 étages elle gagne 4 points … Quand tous les modèles ont été testés, chaque équipe
Remplissez les cases vides.. Tableaux Entrées-Sorties
One may finally note that whereas the relation between perpendicular ion temperature and magnetic field, as well as between parallel velocities and perpendicular temperatures of the
