HAL Id: hal-02607126
https://hal.inrae.fr/hal-02607126
Submitted on 16 May 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Détermination de la biomasse et caractérisation de la
biodiversité dans la forêt tropicale humide de la Guyane
française
R. Muakana Mulumba
To cite this version:
R. Muakana Mulumba. Détermination de la biomasse et caractérisation de la biodiversité dans la forêt tropicale humide de la Guyane française. Sciences de l’environnement. 2017. �hal-02607126�
Détermination de la biomasse et
caractérisation de la biodiversité dans la
forêt tropicale humide de la Guyane
française
Ruth MUAKANA MULUMBA
Directeur du projet : Sylvie DURRIEU (IRSTEA/ UMR TETIS) Jean-baptiste FERRET (IRSTEA / UMR TETIS)
Tuteur SILAT : Pierre BAZILE (AgroParisTech / UMR TETIS)
Rapporteurs : Pierre-Yves VIYON (AgroParisTech / UMR TETIS) Fréderic PORTET (AgroParisTech / UMR TETIS)
LETTRE DE MISSION
Commanditaires Tuteur SILAT Chef de projet
Sylvie DURRIEU - Jean-Baptiste FÉRET Projet Biomass UMR TETIS – IRSTEA +33(0)4 67 54 87 32 sylvie.durrieu@teledetection.fr jean.baptiste.feret@teledetection.fr Pierre BAZILE UMR TETIS – AgroParisTech +33(0)467 548 722 pierre.bazile@teledetection.fr Ruth MUAKANA MULUMBA Mastère SILAT +33(0)770331347 ruthmuak@yahoo.fr Intitulé du projet
Détermination de la biomasse et caractérisation de la biodiversité dans la forêt tropicale humide de la Guyane française : apport et complémentarité des données LiDAR et optiques (Sentinel 2)
Contexte du projet
Les mesures de biomasse forestière permettent de mieux connaître le rôle des forêts dans le cycle du carbone, et plus particulièrement dans le changement climatique. La télédétection permet à cet égard un suivi synoptique et pluriannuel permettant des études des vastes zones. Ainsi, IRSTEA et ses partenaires travaillent sur la détermination de la biomasse forestière par télédétection, notamment dans le cadre des projets TOSCA CNES Biomass et Hypertropik. Le LiDAR est une technologie très prometteuse pour mesurer la structure forestière et en déduire la biomasse. L’acquisition de données LiDAR aéroporté est relativement facile grâce au développement de capteurs industriels opérés par des prestataires de service. Par ailleurs, l'imagerie hyper-spectrale a un fort potentiel pour caractériser la biodiversité en forêt tropicale mais l’acquisition de ce type de données reste plus complexe, les capteurs hyper spectraux étant encore peu répandus. En revanche, le satellite Sentinel 2, lancé en 2015, permet d'obtenir facilement des images gratuites en mode « superspectral ».
Un volet du projet Biomass consiste donc à comparer et à déterminer les synergies qui peuvent exister entre différents types de capteurs (LiDAR / Optique), en particulier dans les zones tropicales (à très forte biomasse), pour améliorer les estimations de biomasse. C'est dans ce cadre que s'inscrit la mission SILAT en y intégrant l’évaluation du potentiel de ces données pour caractériser la diversité spécifique de ces forêts.
Finalité du projet
Analyser les données LiDAR-Optique en vue de montrer puis d’exploiter leur complémentarité dans l'estimation de la biomasse et la caractérisation de la biodiversité en forêt tropicale.
Objectifs du projet
• Estimer la biomasse forestière en utilisant les données LiDAR
•
Caractériser la biodiversité a et b avec les images hyper-spectrales Sentinel 2 et avec les données issues d'inventaire forestier• Évaluer l'incidence des données de la biodiversité sur l'estimation de la biomasse . • Évaluer l'apport du couplage avec des données LiDAR pour caractériser la biodiversité
• Déterminer une méthodologie d'analyse pouvant intégrer ces différents types de données dans la cartographie de la biomasse et de la biodiversité forestière.
Démarche envisagée
• Revue bibliographique
• Prise en main de la base de données : collecte et mise en cohérence des données disponibles sur la forêt de Paracou (données LiDAR, téléchargement des images Sentinel 2 de la zone d'étude, données de l'inventaire forestier);
•
Application les modèles d’analyse de données mis au point par l'IRSTEA pour l'estimation la biomasse et la caractérisation de la biodiversité avec ces différents type de données ;• Analyse comparative des résultats obtenus avec les différents types de données ;
• Proposition d'une méthodologie en vue de tirer parti de la complémentarité des différentes sources de données;
• Proposition d'application et/ou d'adaptation de la méthodologie (avec ces 2 types de données) proposée pour la forêt tropicale humide amazonienne à la forêt équatoriale d'Afrique (République Démocratique du Congo).
Livrables
• Un rapport méthodologique décrivant la méthode d’estimation de la biomasse intégrant les 2 types de données .
•
Une évaluation des diversités a et bSuivi du projet et encadrement
S. Durrieu, JB Féret, E. Grau.
TABLE DES MATIÈRES LETTRE DE MISSION...i TABLE DES MATIÈRES...iii LISTE DE FIGURES...iv LISTE DE TABLEAU...iv I. INTRODUCTION...1 I.1. Contexte de la mission...1 I.2. Objectif de la mission...2 I.3. Zone d'étude...2 II. DEROULEMENT DU PROJET...4 II.1. Définition de la demande...4 II.2. Validation de la lettre de mission...4 II.3. Organisation du travail...4 III. DEMARCHE METHODOLOGIQUE...5 III.1. Analyse de l'existant...5 III.1.1. Les méthodes et modèles d’estimations de la biomasse forestières par télédétection. .5 III.1.2. Télédétection et caractérisation de la biodiversité...7 III.1.3. Caractérisation de la biodiversité avec les données d'inventaire forestier...8 III.2. Les données utilisées...9 III.2.1. Données d'inventaires forestiers de Paracou...9 III.2.2. Les données LiDAR...10 III.3. Choix méthodologiques...11 III.3.1. Choix des variables...11 III.3.2. Démarche de traitement retenue...16 III.3.3. Logiciels utilisés...17 IV. RÉSULTATS ET DISCUSSIONS...17 IV.1. Présentation des résultats...17 VI.1.1. Influence de la taille de la placette sur l'estimation de la biomasse...17 VI.1.2. Caractérisation de la biodiversité...17 VI.1.3. Corrélation entre l'estimation de la biomasse et la caractérisation de la biodiversité 19 IV.2. Discussions...23 IV.2.1. de l'estimation de la biomasse...23 IV.2.2. de la caractérisation de la biodiversité...23 IV.2.3. de la relation biomasse et biodiversité...23 IV.1.4. de l'influence de la stratification forestière sur le modèle d'estimation de la biodiversité...23 V. CONCLUSION...25 BIBLIOGRAPHIE...26 ANNEXES...29 Résumé...35 Abstract...35
ACRONYMES ABA Area-Based Approach, Approche à la placette AGB AboveGround Biomass, Biomasse aérienne AGB1 Biomasse mesurée AGB2 Biomasse estimée ALS Airborne Laser Scanning, Lidar aéroporté AMAP botAnique Modélisation de l’Architecture des plantes et des végétations CIRAD Centre de coopération Internationale en Recherche Agronomique pour le Développement CNES Centre National d'Etudes Spatiales CNRS Centre National de la Recherche Scientifique DBH Diameter at Breast Height, Diamètre à Hauteur de Poitrine DEM Digital Elevation Model, Modèle numérique de hauteur ESA European Space Agency, Agence Spatiale Européenne FORESEE : Forest Resource Estimation for IRSTEA Institut national de Recherche en Siences et Technologies pour l’Environnement et l'Agriculture LAD Leaf Area Densité, Densité de Surface Foliaire LiDAR Light Detection And Ranging MNC Modèle Numérique de Canopée MNS Modèle Numérique de Surface MNT Modèle Numérique de Terrain MPE Mean Percentage Error, Pourcentage d'erreur moyenne PEPS Plateforme d'Exploitation des Produits Sentinel RMSE Root Mean Square Error, Erreur quadratique moyenne RSD Relative standard deviation, Ecart-type relatif SNAP Sentinel Application Platform STEP Science Toolbox Explotation Platform TETIS Territoires, Environnement, Télédétection et Information Spatiale TOSCA Terres solide, Océan, Surfaces Continentales, Atmosphère UMR Unité Mixte de Recherche
LISTE DE FIGURES Figure 1: Zone d’étude, dispositif forestier du site de Paracou, Guyafor (Blanc, 2014)...3 Figure 2: Aperçu d'une image LiDAR de la zone...10 Figure 3:Aperçu d’une image Sentinel 2 couvrant une partie de la Guyane Française (Source : CNES / PEPS)...11 Figure 4: Placettes et arbres échantillonnés...12 Figure 5: Aperçu schématique des traitements des données...16 Figure 6:Indice de Biodiversité pour les placettes 60m...18 Figure 7:Indices de biodiversité pour les placettes de 30 m...18 Figure 8:Relation entre la biomasse et la biodiversité pour les placettes de 60 m de rayon...19 Figure 9:Relation entre la biomasse et la biodiversité pour les placettes de 45 m de rayon...20 Figure 10: Profil de végétation...21 Figure 11:Profil de différentes strates forestières...22 LISTE DE TABLEAU Tableau 1: Informations recueillies sur chaque individu...9 Tableau 2: Caractéristiques des images Sentinel 2...11 Tableau 3: Erreurs du modèle d'estimation de la biomasse...17
I.
INTRODUCTION
I.1.
Contexte de la mission
La présente étude s'inscrit dans le cadre de notre mission professionnelle du mastère spécialisé SILAT, qui s'est déroulée en 6 mois entre avril 2016 et mai 2017 et qui a été commandité par l'Institut National de Recherche en Sciences et Technologies pour l'Environnement et l'Agriculture (IRSTEA) / Centre de Montpellier de l'UMR TETIS.
L'IRSTEA conduit avec différents partenaires (établissements d'enseignement et de recherche) diverses études produisant des connaissances fondamentales tout en développant des méthodes de gestion et procédés pour les entreprises, les collectivités territoriales et les décideurs publiques. L'un des thèmes de recherche de l'IRSTEA/ centre de Montpellier est l'étude des peuplements forestiers par télédétection. En effet, les forêts constituent un immense puits de carbone qui contribue à la régulation du changement climatique, particulièrement les forêts tropicales humides contenant par ailleurs le plus grand réservoir continental de la biodiversité. Cependant, cet écosystème, complexe et difficilement accessible, requiert d'être mieux caractérisé pour approfondir la connaissance de son fonctionnement. Ce qui est essentiel à la gestion durable des ressources qu'il fournit. Parmi les connaissances à développer, les mesures de la biomasse forestière permettent de mieux appréhender le rôle des forêts dans le cycle du carbone et dans le changement climatique (Ferraz et al., 2014). Ainsi, la télédétection permet un suivi synoptique et pluriannuel permettant des études sur des vastes régions à l’échelle mondiale.
C'est dans ce contexte que l'IRSTEA et ses partenaires travaillent sur la détermination de la biomasse forestière par télédétection, notamment dans le cadre des projets TOSCA CNES Biomass et Hypertropik. Le projet Biomass propose d'utiliser un capteur satellitaire SAR en bande P, en cours de développement à l'Agence Spatiale Europenne (ESA), pour mesurer la biomasse forestière à l'échelle globale.
Parmi les technologies de la télédétection, le LiDAR est actuellement très prometteur pour mesurer la structure forestière et en déduire la biomasse forestière. L’acquisition de données LiDAR aéroporté est relativement facile grâce au développement des capteurs industriels opérés par des prestataires de service. Par ailleurs, l'imagerie hyper-spectrale a déjà montré un potentiel sans précédent pour caractériser la biodiversité en forêt tropicale. Mais l’acquisition de ce type de données reste plus complexe, les capteurs hyper spectraux étant encore peu répandus.
En revanche, le satellite Sentinel 2, lancé en 2015, permet d'obtenir facilement des images gratuites en mode «superspectral» avec 13 bandes spectrales de résolution spatiale variant entre 10 et 60 mètres. Il apparaît donc opportun d'évaluer le potentiel de ces données pour caractériser la biodiversité en forêt tropicale. Un volet du projet Biomass consiste à comparer et déterminer les synergies qui peuvent exister entre différents types de capteurs (LiDAR-Optique), en particulier dans les zones tropicales (à très forte biomasse), pour améliorer les estimations de biomasse. En effet, l'imagerie hyperspectrale permet de caractériser la biodiversité forestière et cette dernière pourrait améliorer le modèle d'estimation de la biomasse. C'est dans ce cadre que s'inscrit la présente mission SILAT dont il est proposé d'élargir les objectifs en y intégrant l’évaluation du potentiel de ces données pour caractériser la diversité spécifique de ces forêts.
I.2.
Objectif de la mission
Le but de notre mission a été de comparer les données LiDAR et les images optique en vue d’exploiter leur complémentarité dans l'estimation de la biomasse et la caractérisation de la biodiversité en forêt tropicale.
Plus spécifiquement il s'agissait de (d') :
•
Estimer la biomasse forestière en utilisant les données LiDAR (tester et essayer d'améliorer un modèle existant proposé par l'IRSTEA)•
Caractériser les biodiversités a et b avec les images hyper-spectrales Sentinel 2 et les données issues d'inventaire forestier• Évaluer l'incidence des données de la biodiversité sur l'estimation de la biomasse . • Déterminer une méthodologie d'analyse pouvant intégrer ces différents types de
données dans la cartographie de la biomasse et de la biodiversité forestière.
I.3.
Zone d'étude
Notre zone d’étude se trouve en Guyane française,département d’Outre-Mer de la France, situé au Nord-Ouest de l’Amérique du Sud. Elle est géographiquement localisée entre 2° et 6° de latitude nord et entre 52° et 57 ° de longitude Ouest. Appartenant à la zone Intertropicale, elle baigne dans un climat chaud et humide caractérisé par une périodicité saisonnière pluvieuse et sèche. Il existe en Guyane Française un réseau de dispositifs forestiers permanents connu sous le nom de Guyafor. L'objectif principal de ce réseau est l'étude à long terme de la dynamique forestière et de la biodiversité. Guyafor s’intéresse essentiellement, d'une part aux rôles de l’impact de l’activité humaine dans la dynamique des forêts naturelles et exploitées, d'autre part, au cycle du carbone de la biomasse aérienne vivante, mais aussi aux effets des variations climatiques.
Guyafor constitue une plate-forme de recherche disponible pour les chercheurs ; le CIRAD, le CNRS et l'ONF assurent sa gestion. Certains de ces dispositifs ayant été mise en place depuis les années 70, il est un terrain de prédilection pour des études longue durée sur les fonctionnements des forêts tropicales humides.
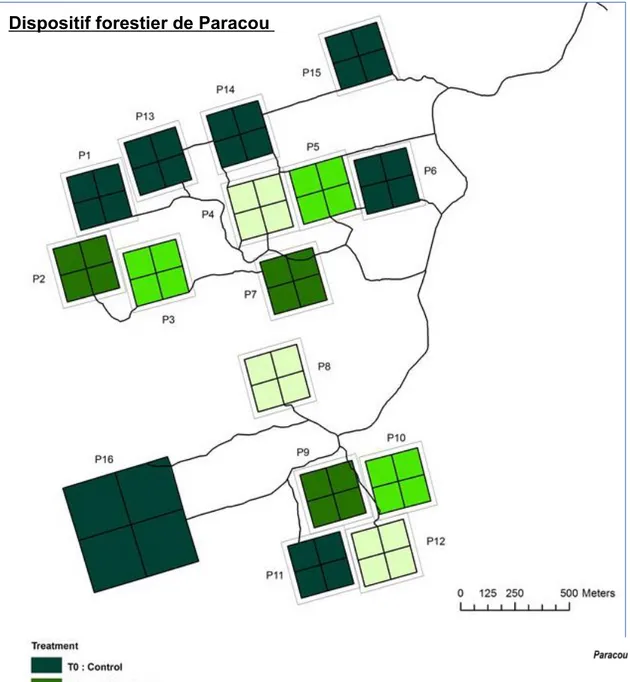
Les dispositifs les plus importants sont ceux de Paracou et des Nouragues qui font continuellement l’objet des programmes de recherche scientifique sur diverses thématiques depuis les années 80. Le site de Paracou, mis en place en 1984, est un dispositif forestier utilisé par l'Office National des Forêts pour l'élaboration et l'amélioration des plans d'aménagements des forêts tropicales. Ainsi, ce dispositif forestier du site de Paracou couvre une surface totale de 5000 ha repartie en : • 15 parcelles de 9 ha par parcelles (nommées P1 à P15) où sont étudiés les impacts de différentes interventions sylvicoles sur le peuplement forestier et sur la régénération. Ces parcelles sont gérées par le CIRAD et le CNRS depuis 1984 ; • 1 parcelle de 25 ha (appelée P16), où sont réalisées les recherches écologiques en milieu très peu perturbé, parcelle gérée par le CIRAD et le CNRS depuis 1991 ;
•
1 parcelle en forêt domaniale où est réalisée le suivie de la régénération forestière après coupe raseCes 15 parcelles constituent notre zone d'étude restreinte. Voici ci-dessous une illustration de la zone ci-haut décrite.
Figure 1: Zone d’étude, dispositif forestier du site de
Paracou Guyafor en Guyane française (Blanc, 2014)
II.
DEROULEMENT DU PROJET
II.1. Définition de la demandeL’objectif principal retenu avec le commanditaire concerne la détermination de la biomasse forestière avec les données LiDAR et la caractérisation de la biodiversité de la forêt de Paracou . Ce travail se rapporte au projet TOSCA BIOMASS implémenté par l'UMR TETIS de l'IRSTEA et l'UMR AMAP du CIRAD.
Notons que ce projet fait suite à une thèse réalisée au sein de l'IRSTEA, qui avait pour objet d'estimer la biomasse aérienne avec les données LiDAR dans les forêts de région tempérée. Il s'agira donc de tester ce modèle (et de l’améliorer dans la mesure du possible) en forêt tropicale. II.2. Validation de la lettre de mission
A partir de l'expression de la demande mentionnée ci-dessus, nous avons abouti à l'établissement de la lettre de mission SILAT présentée au début de ce rapport, lettre de mission qui a été validée par le commanditaire. Cependant, au fil de la réalisation du projet la lettre de mission et le calendrier ont été revus en vue de s'adapter à l’évolution et aux aléas rencontrés tout au long du projet. Ainsi l'utilisation des images Sentinel 2 initialement prévue pour la caractérisation de la biodiversité a été remplacée par l'exploitation des données d'inventaires forestiers. Les images de la zone d'étude ont été pour la plupart couverte de nuages ; son utilisation requérait plusieurs corrections radiométriques supplémentaire dont nous ne disposions pas le temps matériels et les compétences nécessaires sur le moment pour les effectuées.
II.3. Organisation du travail Pour la bonne marche du projet :
• Un calendrier de travail a été proposé et approuvé par le commanditaire; ce calendrier a été modifié et réadapter au fil du projet.
• Les différentes données, les documents reprenant les modèles développés par l’IRSTEA ainsi que les logiciels à utiliser nous ont été transmis par le commanditaire. Des rencontres d’échanges ont été organisé subséquemment à cette transmission du packages matériels-méthodes en vue d’en faciliter la prise en mains. • Des réunions de recadrage pour la compréhension des données et des modèles avec le réalisateur de la thèse, portant sur la caractérisation des structures foresières par LiDAR aéroporté pour l'évaluation de la ressource de la biodiversité floristique, qui a inspiré le présent travail.
• Environs deux rencontres mensuelles de mise au point et de l'évolution de l'avancement ont également eu lieu.
III.
DEMARCHE METHODOLOGIQUE
III.1.
Analyse de l'existant
III.1.1. Les méthodes et modèles d’estimations de la biomasse forestières par télédétection
La technologie de LiDAR aeroporté (ALS : Airborne Laser Scanning) a montré sa fiabilité dans l’estimation de la biomasse aérienne (Ferraz & al, 2014) ce qui a permis de réduire considérablement le besoin des mesures sur terrain. L'analyse des données LiDAR permet de produire les métriques forestières habituellement mesurées sur terrain lors des inventaires forestiers traditionnelles. Ainsi les informations sur la hauteur et la densité des arbres, la hauteur de strate et le taux de couverture de la canopée peuvent être produites à partir des données LiDAR.
Ces métriques produites à partir des données LiDAR permettent d'estimer la biomasse aérienne des forêts autant que les équations allométriques permettent de calculer la biomasse à partir des mesures issues d’inventaires forestiers. Selon Wulder & al (2015), les métriques forestières doivent être calibrées par des modèles de régression linéaire en utilisant les données de la biomasse issues des mesures terrain. De cette calibration découle la possibilité de calculer la biomasse sur les zones dont on dispose des données de télédétection telle que le LiDAR aéroporté. Ces derniers permettent de produire des nuages de points en 3D desquels découlent une modélisation d'une plus grande fiabilité avec moins de bruit mais surtout une très haute résolution spatiale de la structure forestière. Une étude, menée au Portugal sur une forêt de 9 km subdivisée en 40 placettes, a révélé une erreur quadratique moyenne de 0,4 % pour l’estimation de la biomasse par LiDAR et une forte corrélation entre la biomasse mesurée sur terrain et celle calculée avec les données LiDAR. A partir des données LiDAR une triangulation de Delauney a permis de calculer le MNT qui a servi à la production des nuages de points 3D. Ces derniers ont permis avec la méthode de segmentation 3 D de produire le modèle Canopy Probability Density Models (CPDM). Avec ce dernier le taux de recouvrement verticale de la canopée a été déterminé (Ferraz & al, 2014). L'erreur de 0,4% comparée aux estimations terrain de la biomasse, prouve ainsi la précision et l’efficacité de cette méthode. Notons cependant que la fiabilité du résultat dépend de la strate étudiée, la strate de sous-bois étant peu fiable. Au delà de cette corrélation et de la précision de l'estimation de la biomasse par les données Li-DAR, il demeure néanmoins des incertitudes dans la modélisation de la distribution spatiale de la biomasse en forêts tropicales. C'est pour appréhender ce phénomène que l'Université des Antilles a initiée une étude pour modéliser non seulement l'estimation de la biomasse mais aussi l'incerti- tude qui l'entoure en forêt tropicale. Il s'agissait donc de quantifier l'incertitude associée à l'estima-tion de la biomasse d'une parcelle, d’inventorier et d'identifier les sources de cette incertitude, puis de proposer des modèles adaptés permettant de réduire cette incertitude (Molto, 2012). Cette mé-thode a été testé en forêt tropicale amazonienne de la Guyane . De cette étude nous retenons les éléments suivants : • il convient de ne prendre en compte que des arbres avec un diamètre supérieur à 10 cm dans l'estimation de la biomasse, réduisant ainsi l'incertitude qu'apporte la strate sous bois comme l'avait aussi énoncé Ferraz cité plus haut ;
• il ne faut pas négliger la biomasse contenue dans les autres plantes lianes et pal-miers : il existe des modèles développés pour estimer la biomasse de liane en forêt amazonienne ; • pour améliorer les prédictions de biomasse, il serait intéressant de se concentrer sur les gros arbres (plus de 60 cm de diamètre). Car un seul arbre peut peser plusieurs di- zaines de tonnes. Ainsi, 2 ou 3 gros arbres peuvent donc représenter un tiers de la bio-masse d'une parcelle d’un hectare ; • dans un peuplement dominé par des grands arbres, les incertitudes sont fortement ré-duites ; • plusieurs paramètres environnementaux sont susceptibles d’influencer l’estimation de la biomasse et d’en apporter les incertitudes: la géologie, la végétation, la topographie, l'hydrographie, le climat… • Modèle utilisé :
log(M
i) = b
0+ b
1log(D
i) + b
2* log(H
i) + b
3*log(WD
i) +
iavec
Mi : La biomasse, Di le diamètre de l’arbre, Hi la hauteur
WDi la densité du bois.
b0, b1, b2... les coefficients de variabilité i étant l’erreur
Comme indiqué plus haut, le LiDAR demeure une technologie puissante dans l'estimation de la biomasse aérienne forestière. Cependant, la pénétration restreinte du signal dans les forêts limite l’efficacité de la méthode LiDAR dans cette estimation surtout en forêt tropicale. Pour améliorer l'estimation de la biomasse, il conviendrait de combiner les méthodes traditionnelles avec une approche intégrant les paramètres liés au taux de recouvrement horizontal de la canopée (Vega & al, 2015).
C'est pourquoi des études ont été initiées par l'IRSTEA pour développer et améliorer les modèles existants d’estimation de la biomasse forestière. Ceci passera par l’identification des nouvelles variables LiDAR caractérisant la structure forestière et en mettant en exergue les différentes sources d’incertitudes pouvant affecter la précision de ces modèles d’estimation de la biomasse (Bouvier, 2015). La principale de ces études est la thèse de Bouvier (2015) portant sur la caractérisation des struc-tures forestières par LiDAR aéroporté pour l’évaluation de la ressource et le suivi de la biodiversité floristique. Cette étude a utilisé l'approche dite ABA ( Area Based Approch), approche à la placette dans laquelle les attributs forestiers mesurés à l’échelle de la placette lors des campagnes d'inven- taire forestier ont été mis en relation avec les nuages des points LiDAR correspondants. Ainsi plu-sieurs modèles ont été développés pour l’évaluation des attributs forestiers suivants : volume de bois total, volume de bois marchand, biomasse aérienne et surface terrière, ces modèles étant gé-néralisables et adaptables selon les zones d’étude. Les variables LiDAR habituellement utilisées dans cette approche ne décrivaient en général que la distribution verticale de la végétation ; c'est
pourquoi, pour améliorer les modèles de prédictions de la biomasse et d'autres attributs forestiers, de nouvelles variables LiDAR décrivant la structure horizontale de la canopée ont été sélection-nées et introduites dans les modèles. Pour ce faire, quatre variables LiDAR ont été retenus pour décrire la structure 3D des peuplements forestiers. Ce sont ces variables qui ont permis de développer des modèles pour l'estimation des ressources forestières citées ci-dessus. Ces variables LiDAR sont : la valeur moyenne de la hau-teur de la canopée ( µCH), la variabilité de la hauteur de la canopée (2CH), la distribution horizontale de la canopée (Pf), ces trois métriques LiDAR étant liées à la structure de la canopée. Une qua-trième métrique a été ajoutée afin de prendre en compte les arbres non dominants dans le modèle d'estimation de la biomasse forestière : le profil de densité de la surface foliaire (CLAD
). Ce der-nier,
fournit des informations sur l’hétérogénéité verticale de la végétation et sur la végétation en dessous de la canopée. ŷ représentant la valeur ajustée d'un des attributs forestiers (biomasse aérienne, volume to-tal du bois, volume du bois marchand ou surface terrière). µCH : hauteur moyenne de la canopée 2CH : variabilité de la hauteur de la canopée Pf : distribution horizontale de la canopée CLAD : profil de densité de la surface foliaire b0, b1, b2... les coefficients de variabilité Notons que le modèle a été développé pour 3 types de forêt différents : forêt de conifères, forêt dé-cidue et forêt de montagnes. Pour vérifier la qualité du modèle d'estimation des ressources forestières par les données LiDAR, les erreurs suivantes entre la biomasse mesurée avec les données terrains et la biomasse estimée avec les données LiDAR ont été déterminées : • l’écart type relatif (RSD) • l’erreur moyenne en pourcentage (MEP) • Erreur Quadratique Moyenne (RMSE) Ainsi l’erreur de prédiction a varié entre 12,4 % à 24,2% pour le modèle général et pour le modèle spécifique il a été amélioré entre 2,0% à 5,3 %. Le résultat de cette étude a montré une améliora-tion du modèle de l’estimation de la biomasse aérienne. 34 études utilisant le modèle classique d’estimation de la biomasse ont fourni des prédictions d’AGB à partir de données ALS avec une moyenne de RSD autour de 27% tandis que le modèles développés dans la présente étude ont fournis des prédictions d’AGB avec un RSD compris entre 12,9% et 22,3 % en fonction du type de forêt.L’étude ci-dessus a été conduite sur les forêts de région tempérée, il s’agira pour nous de reprendre les modèles qui y ont été développés pour les appliquer dans les forêts tropicales et de les améliorer en rajoutant les métriques LiDAR adaptées et visant une prédiction plus précise de
l’AGB aérienne avec les données ALS.
III.1.2.Télédétection et caractérisation de la biodiversité
La télédétection peut être utilisée dans la caractérisation de la biodiversité pour déterminer la diversité a qui est la richesse et l'abondance des espèces à l’échelle locale) et aussi pour déterminer la diversité b qui est la distribution spatiale des communautés d'espèces). En effet , dans le spectre électromagnétique, chaque espèce végétale répond différemment à la lumière et cette réponse détermine sa signature spectrale. Cette dernière est liée à la composition biochimique de la plante.
La classification supervisée des images satellites permet de faire ressortir les signatures spectrales des différentes espèces avec la structure de la canopée et la chimie foliaire. L'association d'informations spatiales et spectrales permet d’augmenter la qualité de la classification. Notons néanmoins qu'une résolution spectrale très faible rend la différenciation des espèces difficile tandis qu'une résolution spatiale trop forte augmente la variabilité spectrale entre les classes.
Cependant, dans la caractérisation de la biodiversité, la classification supervisée présente quelques lacunes même avec les images de forte résolution spatiale et spectrale. Elle nécessite la connaissance de la zone d'étude et la disposition de zones d’entraînement ce qui n'est pas toujours possible dans les vastes zones de forêt tropicale difficiles d'accès (Mafhoud, 2009). Par ailleurs cette approche ne peut être utilisée que pour un nombre limité d'espèces au risque d'avoir des similarités spectrales entre espèces différentes (Ferret et al 2014).
Pour améliorer la caractérisation de la biodiversité à partir des images hyper-spectrales plusieurs approches de classification non supervisées sont proposées notamment dans les forêts tropicales. Ces approches permettent de caractériser la diversité alpha en déterminant les indices de Shannon et de Fisher, mais aussi la diversité bêta en déterminant l'indice de Bray-Curtis. III.1.3. Caractérisation de la biodiversité avec les données d'inventaire forestier Il existe un grand nombre d'indices dans la caractérisation de la biodiversité. Pour caractériser la biodiversité a de la forêt de Paracou, nous avons opté pour l'indice de Shannon, l'indice alpha de Fisher et la richesse spécifique. L'indice de Shannon permet de révéler la diversité spécifique d'un milieu. Cette diversité se traduit par le nombre d'espèces que regorge ce dit milieu ainsi que par la répartition des individus au sein de ces espèces (Marcon,2015). Quant à lui, l'indice a de Fisher permet de déterminer la diversité en se basant sur le nombre total d'individus d'une communauté et le nombre d'espèces auxquelles appartiennent ces individus. Par ailleurs, la richesse spécifique détermine le nombre d’espèces présentes sur une aire donnée, elle dépend fortement de l'effort d'échantillonnage (Marcon ,2015). Pour la caractérisation de la diversité b il est proposé de calculer l’indice de dissimilarité de Bray-Curtis Il se calcule avec la formule suivante :
D=
∑
s |n
s,1−n
s, 2
|∑
s(n
s, 1−n
s,2)
Où :•
D : dissimilarité de Bray – Curtis ;•
ns,1 et ns,2 sont le nombre d’individus de l’espèce s dans la première et la deuxième communautéIII.2.
Les données utilisées
III.2.1. Données d'inventaires forestiers de Paracou
Les données terrains proviennent d'inventaires forestiers effectués dans le site de Paracou du dispositif GUYAFOR en Guyane Française.Elles nous ont été transmises par l’IRSTEA sous forme d’une base de données Excel.
Le tableau ci-dessous renseigne sur les paramètres recueillis lors de l’inventaire forestier dont découle les données que nous avons utilisées au cours de cette étude Code Informations Nom Foret Nom de la forêt d'appartenance de l'individu Libelle Numéro de la parcelle d'appartenance de l'individu sur le site de Paracou NumCarre Numéro du carré d'appartenance de l'individu NumArbre Numéro de l'individu sur la base de données GUYAFOR X Coordonnée X de l'individu sur la parcelle Y Coordonnée Y de l'individu sur la parcelle Xutm Coordonnée X de l'individu en WGS84 UTM 22N Yutm Coordonnée Y de l'individu en WGS84 UTM 22N Circonference Circonférence de l'individu (cm) Annee Année de mesure lors du dernier inventaire (2011 ou 2010 pour la parcelle 16) MesuMort Statut de l'individu (0 = mort ; 1 = vivant) idCodeMesure Qualité de la prise de données Famille Famille de l'individu Genre Genre de l'individu Espèce Espèce de l'individu
Tableau 1: Informations recueillies sur chaque individu
Les données d'inventaires utilisées ont été collectées sur les places P1 à P15 présentées dans la figure 1. Ces données recueillies datent de 2013, les informations ainsi fournies vont permettre de calculer la biomasse mesurée qui servira de base de comparaison à l'estimation de la biomasse avec les données LiDAR.
III.2.2. Les données LiDAR
Elles ont été produites dans le cadre du projet TOSCA BIOMASS et sont issues d'une acquisition effectuée en 2013 dans le site de Paracou avec une densité de 15 points/m². Ces données permettent de produire des Modèles Numériques de Terrain (MNT), de Canopée (MNC) et de Surface (MNS). Le modèle numérique de canopée a fourni les informations sur la hauteur et la surface des houppiers, variables importantes pour l'estimation de la biomasse à partir des données LiDAR (Bouvier, 2015)
Figure 2: Aperçu d'une image LiDAR de la zone
III.2.3. Les images Sentinels 2 Au cours l'année 2016 les images Sentinel 2 ont été mises à disposition du grand publique. Elles sont téléchargeables sur la plateforme PEPS/ SNAP du CNES, à l'état brut ou sur le site du pôle Theia, sous formes d'images corrigées. Nous avions donc prévu au cours de cette étude d'utiliser l’image hyper-spectrale Sentinel 2 pour la caractérisation de la biodiversité. Voici les caractéristiques des images Sentinel 2 selon l'Agence Spatiale Européenne : Caractéristiques Capteurs Visible, PIR Résolution temporelle 10 jours Résolution spectrales 13 bandes dont 3 PIR Résolution spatiale 10 m, 20m ou 60 m selon les bandes Largeur de la scène / Champ de vue 290 km
Figure 3:Aperçu d’une image Sentinel 2 couvrant une partie de la Guyane Française (Source :
CNES / PEPS)
III.3.
Choix méthodologiques
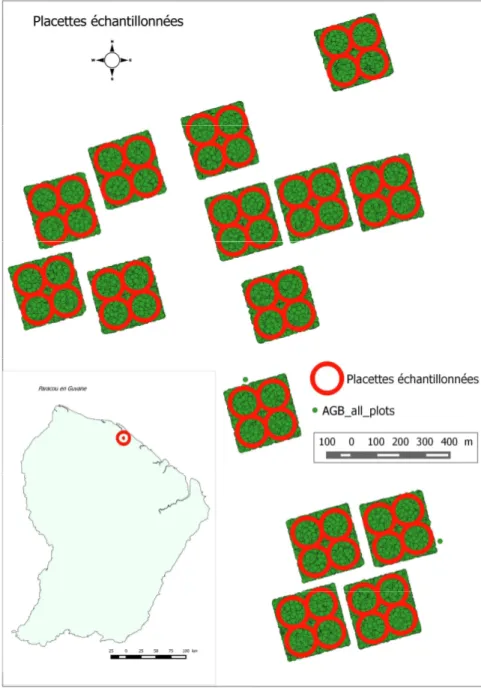
III.3.1. Choix des variablesPartant du modèle proposé par l'IRSTEA, développé au cours de la thèse de Bouvier (Caractérisation des structures forestière par LiDAR aéroporté pour l’évaluation de la ressource et le suivi de la biodiversité floristique), nous l’avons testé pour une forêt tropicale en faisant varier certaines métriques et attributs forestiers. Dans ce modèle, il s'agit d’une approche «à la placette» développée pour mettre en relation les attributs forestiers mesurés à l’échelle des placettes lors d’inventaires avec des métriques calculées à partir des nuages de points LiDAR correspondant à ces placettes. Au cours du développement de ce modèle était ressorti l'impact du nombre et de la taille des placettes sur la précision de l'estimation de la biomasse calculée à partir des données LiDAR. Ainsi nous avons fait le choix de faire varier la taille de placette tout en conservant le même nombre afin de proposer le diamètre adéquat qui améliore l'efficacité de ce modèle pour les forêts tropicales. i. L’échantillonnage Sur base des parcelles du dispositif de GUYAFOR, des placettes circulaires ont été extraites. Ainsi l'échantillon pour la présente étude est composé de 60 placettes. Pour les analyses, ces 60 placettes seront déclinées en 7 séries de 60 placettes de rayon différent allant de 60 m à 30 m avec un intervalle de 5 m. Ainsi les analyses des données s'effectueront sur des séries des 60 placettes à 60m, de 60 placettes à 55 m, 50m, 45m, 40m, 35m et 30m. Ceci permettra entre autre d’évaluer l'impact du diamètre de la placette sur les variables à étudier.
Figure 4: Placettes et arbres échantillonnés
ii. Estimation de le biomasse
Pour l'estimation de la biomasse, plusieurs variables ont été utilisées. Premièrement pour obtenir l'AGB mesurée, calculé à partir des données d'inventaires terrains, le modèle linéaire suivant de Chave a été appliqué : log (B)=α+β1 log (D) x β2 log (H) x β3 log (ρ) Ce modèle prend en compte les variables tels que : • (D) le diamètre à hauteur de poitrine, ( cm)
• (H) la hauteur (m) • ( la densité des arbres (kg/m3) Deuxièmement pour l'AGB prédit, estimé à partir des données LiDAR le modèle suivant mise au point par Bouvier (2015) a été appliqué : Ce modèle prend en compte les métriques LiDAR telles que : • ŷ : biomasse aérienne ( Mg/ha)
•
( µCH) : la valeur moyenne de la hauteur de la canopée•
(2 CH) : la variabilité de la hauteur de la canopée•
(Pf) : la distribution horizontale de la canopée•
(CLAD) :le profil de densité de la surface foliaire•
b0, b1, b2, coefficient de variabilité…Ces deux modèles ont été appliqués sur chacune de 7 séries de 60 placettes que constitue l'échantillon. Ainsi nous avons calculé et estimé l'AGB pour 60 placettes de 30m, 35m, 40m, 45m, 50m, 55m et de 60 m de rayon. Ensuite les erreurs d'estimation suivante ont été calculer pour trouver à quel diamètre de placette les erreurs sont minimiser dans le modèle d'estimation de la biomasse utilisé : • l’écart type relatif (RSD) • pourcentage d’erreur moyenne en (MEP) • Erreur quadratique moyenne (RMSE) iii. Caractérisation de la biodiversité
Trois indices ont été utilisés pour la caractérisation de la biodiversité a : la richesse spécifique, les indices de Shannon et de Fisher. Pour caractériser la biodiversité avec les images hyper-spectrales Sentinel 2, une méthodologie est proposée par l'IRSTEA et mise au point par Ferret (2014).Cette méthodologie préconise de quantifier l’homogénéité spectrale liée à la biodiversité. A cet effet trois méthodes sont proposées : • la détermination de la distribution spectrale des espèces dit SSD • la détermination du NDVI • le calcul de la Distance Moyenne du Centroïde spectrale dit MCD N'ayant pas pu avoir les images Sentinel 2 de la zone d'études sans nuage en vue d'une utilisation correcte, la biodiversité b a été caractérisée qu'avec les données d'inventaires terrains. Ces der- nières ont été utilisées pour calculer les indices de Shannon et de Fisher ainsi que la richesse spé-cifique.
Indice de Shannon Traditionnellement elle se calcule selon la formule suivante :
H ' =−
∑
i=1 SPi log
2Pi
Où : • Pi = abondance proportionnelle ou pourcentage d'importance de l'espèce : pi = ni/N; • S = nombre total d'espèces; • ni = nombre d'individus d'une espèce dans l'échantillon; • N = nombre total d'individus de toutes les espèces dans l'échantillon.Indice alpha de Fisher
Il est obtenu grâce à la formule suivante :
S=α ln
(
1+ N / α
)
avec : • α est la diversité alpha de Fisher, • N le nombre d’individus et • S le nombre d’espèces Richesse spécifique elle est théoriquement calculée par la formule suivante :S=NcAz
• N le nombre d'espèces • A la surface échantillonnée • c et z des constantesVoici les étapes de la démarche initialement prévue pour caractériser la biodiversité avec les images hyper-spectrale (méthodologie mise au point par l'IRSTEA):
• suppression de continuum (Continuum Removal CR) pour éliminer les variations de la luminosité globale
• analyse des Composantes Principales (ACP) pour réduire la dimensionnalité spec-trale
• sélection aléatoire des sous ensemble des pixels de l’image et partition des spectres des espèces et localisation de centroïde de sous ensemble des pixels. • division en unité cartographique des données spectrales pour affectr chaque pixel
de l’image à un spectre donné représentant une espèce.
• obtention d’une distribution spectrale des espèces pour chaque unité cartogra-phique ;
• les étapes ci-dessus sont ensuite répétées 100 fois pour diminuer le risque de l’échantillonnage aléatoire non représentatif. Avec les données d’inventaire forestier, la caractérisation de la diversité b peu se faire en déter-minant l'indice de Bray Curtis (qui se calcule comme suit:)
D=
∑
s |n
s,1−n
s, 2
|∑
s(n
s, 1−n
s,2)
Où :•
D : dissimilarité de Bray – Curtis ;•
ns,1 et ns,2 sont le nombre d’individus de l’espèce s dans la première et la deuxième communauté.iv. Profil de végétation
Le profil de densité de la surface foliaire fournit des informations sur l’hétérogénéité verticale de la végétation et sur la végétation en dessous de la canopée. Le profil de végétation permettra de faire une stratification forestière, en effet le profil représente la densité de nuages de points LiDAR du sol au sommet de la canopée. Il est calculé pour chaque placette à partir des données LiDAR et le MNT de la zone d'étude. La détermination du profil de végétation passe par le calcul de l'indice LAD (Leaf Area Densité) . Cette dernière devra ensuite être normalisé et son coefficient de variation déterminée. De là découlera la densité de la surface foliaire exprimer en m²/m² par intervale d'un mètre de hauteur de la canopée. Les valeurs du profil de végétation (par mètre de hauteur) ainsi obtenues, ont été agrégés pour avoir le profil de végétation par strates forestières. Pour ce faire nous avons utilisé l’échelle de hauteur de Malcom proposée par Ferraz (2012) pour les forêts tropicales. Il s'agit de repartir la canopée comme suit : de 0 à 2m, de 2m à 10m, de 10 à 20m, de 20 à 30m, de 30 à 40m et la hauteur supérieure à 40m.
Il s'agira enfin de déterminer l'influence de la variation de la hauteur ( strates forestières ) de canopée sur la performance du modèle d'estimation de la biomasse utilisé au cours de la présente étude. Ceci en déterminant la corrélation existant entre la biomasse estimée et la hauteur cumulée de la canopée.
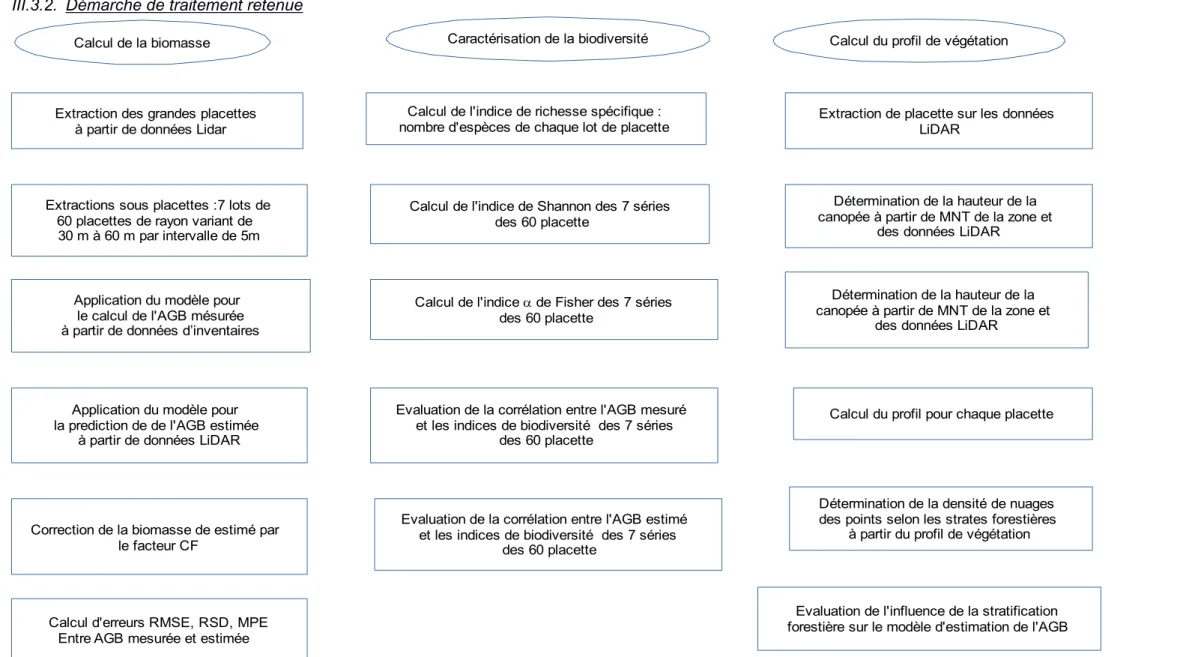
III.3.2. Démarche de traitement retenue
Calcul de la biomasse Caractérisation de la biodiversité Calcul du profil de végétation
Extraction des grandes placettes à partir de données Lidar Extractions sous placettes :7 lots de 60 placettes de rayon variant de 30 m à 60 m par intervalle de 5m Correction de la biomasse de estimé par le facteur CF Application du modèle pour le calcul de l'AGB mésurée à partir de données d’inventaires Calcul d'erreurs RMSE, RSD, MPE Entre AGB mesurée et estimée Calcul de l'indice de richesse spécifique : nombre d'espèces de chaque lot de placette Extraction de placette sur les données LiDAR Calcul de l'indice de Shannon des 7 séries des 60 placette Calcul de l'indice a de Fisher des 7 séries des 60 placette Evaluation de la corrélation entre l'AGB mesuré et les indices de biodiversité des 7 séries des 60 placette Evaluation de la corrélation entre l'AGB estimé et les indices de biodiversité des 7 séries des 60 placette Détermination de la hauteur de la canopée à partir de MNT de la zone et des données LiDAR Détermination de la hauteur de la canopée à partir de MNT de la zone et des données LiDAR Calcul du profil pour chaque placette Détermination de la densité de nuages des points selon les strates forestières à partir du profil de végétation Evaluation de l'influence de la stratification forestière sur le modèle d'estimation de l'AGB Application du modèle pour la prediction de de l'AGB estimée à partir de données LiDAR
III.3.3. Logiciels utilisés
Le logiciel R a été principalement utilisé pour les analyses et les traitements des données au cours de cette étude. Pour le calcul et l'estimation de la biomasse ainsi que le calcul des erreurs liées au modèle de cette estimation, nous avons utilisé la fonction de régression linéaire multiple. Les packages vegan et entropart ont été utilisé pour le calcul d'indices de biodiversité qui a servi à la caractérisation de la biodiversité. Pour déterminer le profil de végétation c'est le package LidR qui a été utilisé. Par ailleurs il avait été prévu d'utiliser la plate forme PEPS (Plate forme d'Exploitation des Produits Sentinel) en vue de l'acquisition des images satellites Sentinel 2. La plate forme SNAP (Sentinel Application Platform) ainsi que la boite à outil Sentinel 2 Toolbox devraient servir pour les analyses et traitement des images hyper-spectraux.
IV.
RÉSULTATS ET DISCUSSIONS
IV.1.
Présentation des résultats
VI.1.1. Influence de la taille de la placette sur l'estimation de la biomasse
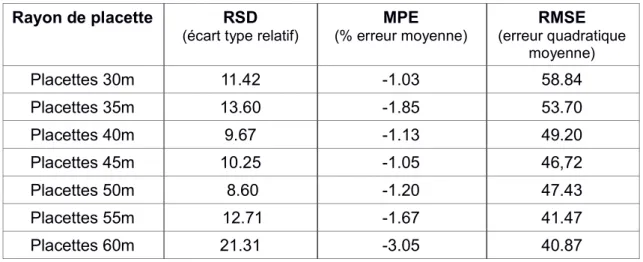
Nous avons calculé l'AGB dit mesuré avec les données terrains et nous avons appliqué un modèle d'estimation de la biomasse à partir des métriques issues de données LiDAR . Pour vérifier la précision prédictive de ce modèle, nous avons calculé les erreurs d'estimation pour chaque série de 60 placettes de différents rayon. Le tableau (3) ressort ces erreurs ainsi calculées : Rayon de placette RSD (écart type relatif) MPE (% erreur moyenne) RMSE (erreur quadratique moyenne) Placettes 30m 11.42 -1.03 58.84 Placettes 35m 13.60 -1.85 53.70 Placettes 40m 9.67 -1.13 49.20 Placettes 45m 10.25 -1.05 46,72 Placettes 50m 8.60 -1.20 47.43 Placettes 55m 12.71 -1.67 41.47 Placettes 60m 21.31 -3.05 40.87
Tableau 3: Erreurs du modèle d'estimation de la biomasse
Pour évaluer la performance du modèle selon la taille de la placette calibrée, nous avons déterminé les écarts entre la biomasse observée et la biomasse prédite. En ayant un focus sur l'écart type relatif (RSD), il ressort, suivant la superficie de la placette déterminée dans la calibration du modèle, que le RSD varie de 21,31 à 8,60. Et au vue de ces résultats, il se pourrait que le modèle ait une performance relativement bonne avec les placette calibrées entre 40 et 50 m. Cependant, cette disparité n'est pas la même en faisant un focus sur le RMSE. Nous remarquons qu'à cet effet, plus la taille de la placette augmente plus le RMSE est minimisé et la performance du modèle semble ainsi s'améliorer. La Cette différence pourrait être dû probablement par le fait que le RMSE exerce une plus grande influence sur de grandes erreurs que sur les petites. Tandis que le RSD représentation la qualité globale du modèle d'estimation de la biomasse.
VI.1.2. Caractérisation de la biodiversité
Pour la caractérisation de la biodiversité 3 indices de biodiversité ont été calculés :la richesse spécifique exprimée en nombre d’espèce, l'indice alpha de Fisher et l'indice de Shannon.
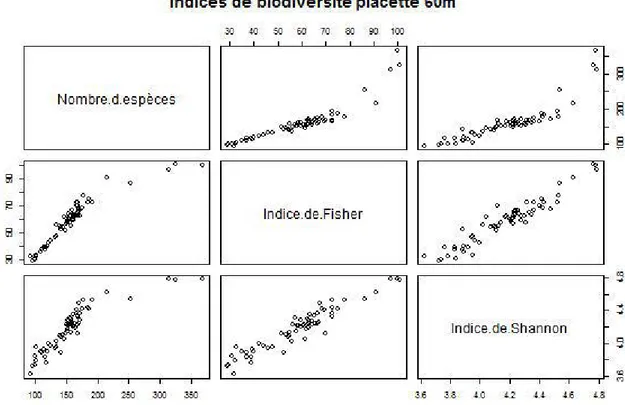
Le graphique ci-dessous ressort les valeurs obtenues pour chacune de ces indices dans le site forestiers de Paracou. Chaque point du graphique ci-dessous représente la valeur de l’indice pour une placette.
Nous avons choisie de présenter ici les extrêmes c’est-à-dire les indices de biodiversité pour les placettes de 60 m et de 30 m de rayon. Ceci pour entrevoir les intervalles de valeurs entre lesquelles naviguent les indices de la biodiversité pour notre zone d'étude.
Figure 6:Indice de Biodiversité pour les placettes 60m
Figure 7:Indices de biodiversité pour les placettes de 30 m
Le nombre d’espèces varient entre 368 et 94 pour les placettes de 60 m de rayon (1,13 ha) et pour les placettes de 30 m (0,28 ha) le nombre d’espèce varie entre 216 et 48. Ce qui révèle une grande richesse spécifique. Notons cependant que cette dernière dépend de la taille de l'échantillon et ne tient pas compte de l'abondance relative de différentes espèces.
L'indice de Shannon pour notre site est compris entre 3,2 et 4,78 sur une échelle de 0 à 5. Ce qui traduit une forte diversité spécifique. Cet indice prend en compte la richesse spécifique, l'abondance des espèces et dans une moindre mesure la dominance d'une ou plusieurs espèces. Sachant qu'une dominance annoncée d'une seule espèce traduit une faible diversité tandis que la co-dominance de plusieurs espèces traduit une grande diversité. L'indice de Fisher quant à elle varie de 100,74 à 29,11 pour les placettes de 60 m de rayon (1,13 ha) et de 87,05 à 24,32 pour les placettes de 30 m de rayon. Nous n'avons pas calculé l'indice de Bray – Curtis qui est obtenu en comparant au moins deux échantillons spatialement différent formant des communauté d'espèces distinct. Ayant travaillé qu'avec un seul échantillon, forêt de Paracou, ceci ne nous a pas permis de calculer l'indice de Bray-Curtis et de l'intégrer dans nos analyses.
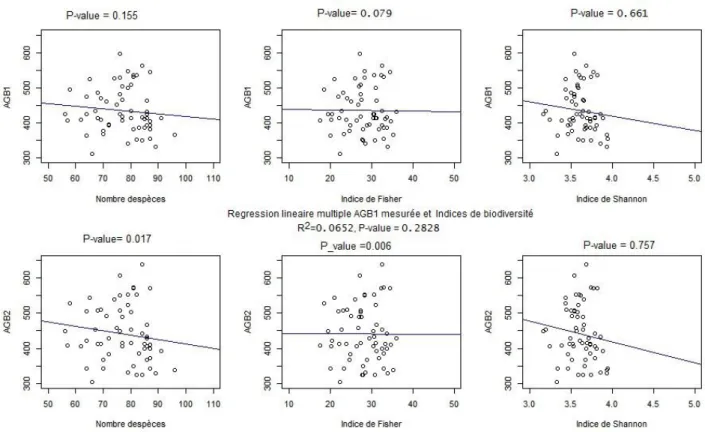
VI.1.3. Corrélation entre l'estimation de la biomasse et la caractérisation de la biodiversité L'un des objectifs de cette étude était d'évalualer l'effet de la biodiversité sur l'estimation de la biomasse avec les données LiDAR. Les graphes ci-dessous montre la corrélation entre les AGB (estimée et mesurée) et les indicatuers de biodiversité. Nous le présentons ici les placettes de 45 m de rayon pour lesquelles notre modèles a enregistré la meilleur performance pour les forêts tropicales.
Figure 8:Relation entre la biomasse et la biodiversité pour les placettes de 45 m de rayon
De ce qui précède, il ressort des coefficients de corrélation inférieur à 0,2 entre les indices de biodiversités utilisées (à savoir les indices de Shannon, de Fischer et la richesse spécifique) et la biomasse estimée. Ceci pourrait être traduit par le fait qu'environ 20 % de la variation de la
biomasse serait expliquée par la variation de la biodiversité exprimées par ces trois indices ci-haut mentionnées.
Ceci pourrait signifier qu'avec notre modèle d'estimation de la biomasse utilisée ici, il serait plus opportun et pertinent d'appliquer d'autres descripteurs de la biodiversité et que l'ajustement linéaire de la biomasse estimée puisse rendre en compte au moins à 80 % la variation de cette biomasse.
Ces indicateurs biodiversité qui pourraient avoir des valeurs de corrélation d'environ 0,8 avec la biomasse estimée à partir des données LiDAR pourraient alors être proposé dans l'amélioration du modèle que nous avons utilisé au cours de cette étude.
IV.1.4 La stratification forestière et l'estimation de la biomasse
Nous avons calculé le profil de densité de la surface foliaire qui nous a fournit les informations sur l’hétérogénéité de la canopée.La densité de la surface foliaire exprimée en m²/m² a été déterminer sur toute la hauteur de la canopée allant de 1,5 m à 49 m a été déterminé par intervalle de 1m. Les différentes densités de végétation ainsi obtenue (pour chaque mètre de hauteur) ont été agrégées sur base de la stratification des forêts tropicales. Ce qui a permis d’obtenir la densité foliaire selon les strates forestiers variant comme suit : de 0 à 2m, de 2m à 10m (strate du sous bois), de 10 à 20m (strate de dominés), de 20 à 30m (strate de dominés), de 30 à 40m (strate de dominants) et la hauteur supérieure à 40m ( strate de émergents).
Le dispositif forestier du site de Paracou (Figure 1) est constitué d'une part des placettes témoins dit parcelles de contrôle et d’autres part des placettes subissant des traitements sylvicoles (exploitation de bois d’œuvres, exploitations des bois énergies). L’analyse ci-haut decrite a été effectuée séparément, d’une part pour les placettes témoins (qui ne subissent aucune exploitation forestière) et d’autre part pour les placettes subissant les exploitations forestières. Ensuite l’influence de cette variation de la hauteur de la canopée sur la performance du modèle a été évalué, en établissant la corrélation entre la variation de la biomasse estimée et la variation de la hauteur de canopée cumulée.
Les tableaux ci-dessous présentent le résultat de cette analyse : Profil pour placette témoins (sans traitement sylvicoles)
Rayon
placettes
2 – 10 m
10 – 20m
20 – 30m
30 – 40m
> 40m
30 m
0,026
0,137
0,183
0,064
0,032
35 m
0,005
0,159
0,293
0,154
0,010
40 m
0,005
0,052
0,120
0,067
0,002
45 m
0
0,030
0,049
0,046
0
50 m
0,005
0,003
0,006
0,023
0,012
55 m
0,002
0,049
0,070
0,054
0,075
60 m
0,003
0,057
0,089
0,073
0,082
Profil pour placette traités (avec traitement sylvicoles)
Rayon
placettes
2 – 10 m
10 – 20m
20 – 30m
30 – 40m
> 40m
30 m
0,010
0,006
0
0,142
0,149
35 m
0,077
0,021
0,002
0,160
0,314
40 m
0,120
0,026
0,016
0,136
0,376
45 m
0,148
0,029
0,031
0,129
0,368
50 m
0,076
0,025
0,020
0,175
0,303
55 m
0,090
0,021
0,010
0,189
0,249
60 m
0,097
0,032
0,017
0,196
0,305
Il ressort de ce résultat des faibles valeur de coefficient de détermination de manière générale traduisant ainsi une moindre corrélation entre la biomasse estimée avec le modèle utilisé ici et la stratification forestière, tant pour les placettes témoins que pour celles subissant des exploitations forestières. Cependant, selon nos résultats, les strates des dominants ( 30-40 m de hauteur) et des émergeant (hauteur de canopée > 40 m) pour les placettes sous exploitations, ont quant à eux des valeurs un peu plus supérieurs que les autres strates. Cela pourrait peut être indiquer que pour ces placettes sous exploitation, bien que dans une moindre, mesure les strates plus haut contribuerai plus à la biomasse.IV.2. Discussions
IV.2.1. L'estimation de la biomasse
Selon Bouvier (2015), les modèles classiques d'estimation de la biomasse aérienne révèlent des erreurs de prédiction oscillant autour de 27 % de RSD . Cependant son modèle que nous avons utilisé minimise cette erreur avec des valeurs du RSD échelonnée entre 12,4 % à 22,3 %. Nous avons obtenu des valeurs de RSD allant de 8,60 à 21,31 %, ce qui se rapproche aux valeurs obtenus par Bouvier. Par ailleurs, le RMSE que nous avons obtenu au cour de cette étude est compris entre 58,84 et 40,87 ce qui rentre dans l’intervalle des valeurs de RMSE trouvé par Vega & al (2015) variant entre 76, 42 et 28,83. Le résultat obtenu par Bouvier (2015) en utilisant le même modèle prévoit un RMSE 53,1 et nos résultats s'y rapprochent relativement, surtout pour les placettes de 35 m de rayon. Notons que la performance de ce modèle d'estimation de la biomasse que nous avons utilisé est impactée par plusieurs sources d'erreurs (Bouvier ,2015). Notamment,le nombre de placettes utilisées, la taille de la placette, la précision du positionnement GPS du centre de la placette, la calibration des données LiDAR et la précision des mesures récoltées sur terrain.
Le nombre de placettes préconisé pour une bonne performance du modèle est de 100 à 200 placettes ; les données d'inventaires à notre disposition nous ont permis de n’avoir que 60 placettes. Néanmoins c'est en dessous de 50 ou 40 placettes échantillonnées que l'erreur augmente significativement. Concernant la taille de placette , Bouvier (2015) recommande un rayon de placette variant de 15 à 6 m pour les forêts tempérées ; cependant, pour une estimation de la biomasse en forêt tropicales humides, la taille de placette minimum recommandée est de 30
m de rayon. Nous avons aussi respecté la précision du positionnement GPS du centre de la placette, préconisée entre 0 et 5 m en fixant notre précision GPS à 0,5 m.
L'impact de la calibration des données LiDAR ainsi que de la précision de la collecte des données terrain sur la qualité du modèle étant jugées mineurs, nous nous sommes pas apesantie sur ces sources d’erreurs.
Ainsi, au vu de nos résultats (tableau 3) et des paramètres recommandés pour appliquer ce modèle en forêt tropicales, le taux d'erreur (RSD) semble minimiser pour les placettes de taille variant entre 40 m et 50 m de rayon. Par ailleurs, concernant le RMSE, sa valeur diminue avec l'augmentation de la taille de la placette, avons nous par ailleurs observé. Cela serait dû au fait que la grande taille de placette diminue les biais dû au effets des bord. Ce qui expliquerai probablement la performance du modèle qui s’accroît avec l'augmentation de la taille de la placette. Par contre le nombre des placettes a été pour nous une source majeur d'erreur.
IV.2.2. La caractérisation de la biodiversité
Les forêts tropicales regorgent d'une grande richesse spécifique,selon Blanc (2014). Dans certaine partie de la forêt amazonienne il est possible de trouver de zone où l'on rencontre plus de 300 espèces par hectare. C'est dans cette ordre d'idées que nos analyse ont révélé une forte biodiversité alpha. Cette dernière est caractérisée dans la forêt de Paracou par des indices biodiversité élevée. Ainsi nous avons enregistré pour les placettes de 60 m (Figure 7) des indices de Shannon allant de 3,2 à 4,78 ce qui traduit une forte diversité telle que l'indique aussi le nombre d'espèce allant jusqu'à plus de 350 espèces pour une placette d'environ 1,1 ha.
La grande variabilité des indices des biodiversités (368 et 94 espèces) entre les placettes de la notre zone d'étude serait dû au différent traitements appliqués aux placettes (Figure 1).
Les placettes témoins gardent une diversité inférieur à celle des placettes exploitées car avec les ouvertures créées par l'exploitation il y a une invasion de nouvelles espèces héliophiles non présentent sur les placettes non exploitées. Cependant cette variable ne faisant pas l'objet de notre étude nous ne l'avons pas approfondi.
IV.2.3. La relation biomasse et biodiversité
La biomasse est généralement influencée par la biodiversité, l'environnement et la répartition spatiale des espèces. Selon Chisholm cité par Husson 2014 la relation entre la diversité et la biomasse est plus prononcée pour les petites placettes inférieurs à 0,04 ha : plus la diversité floristique est grande plus la biomasse augmente. Cependant en effectuant les analyses avec les placettes dont le superficie varie entre 0,25 et 1 ha il ne ressort qu'une très faible relation entre la biodiversité et la biomasse qui est généralement négatif. Cette thèse semble être vérifié par les résultats obtenus au cours de cette étude, en effet les placettes d'échantillonnage que nous avons utilisées ont un rayon minimum de 30 m et maximum de 60 m ce qui correspond à des placettes dont les superficies varient de 0,2826 à 1,1304.
La biomasse dépend entre autre de la répartition spatiale des individus d'une espèce (Marcon, 2015). L'indice de Shannon quant à elle prend en compte le nombre d'individus d'une même espèce. Nous savons par ailleurs qu'en forêt tropicales humides on retrouve une grande quantité d'espèces à l'hectare mains une moindre représentation d’individus de la même espèce. Ce qui conduit à reposer la question sur la pertinence de l'indice de Shannon pour améliorer ce modèle d'estimation que nous avons utilisée en forêt tropicale.Selon nos résultats