An experimental model of cryoglobulin-associated vasculitis in mice
Texte intégral
Figure
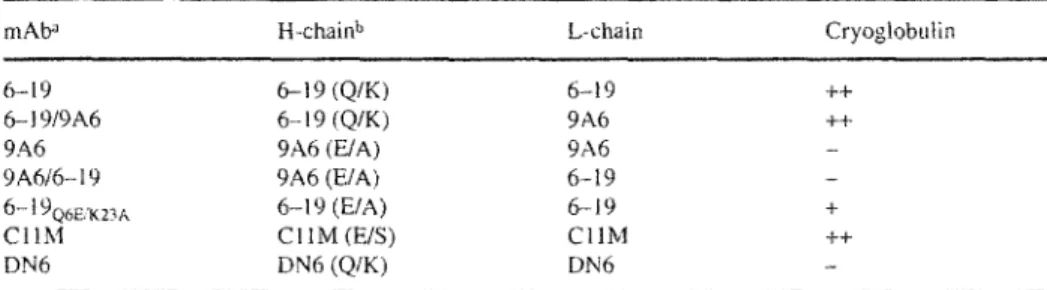

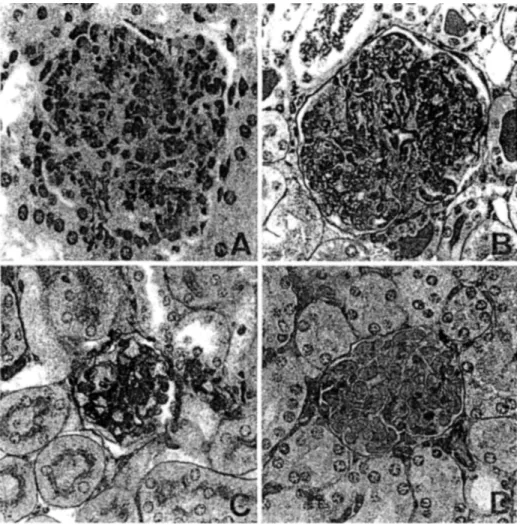
Documents relatifs
Considering the lack of data concerning olfactory laterality in rodents in literature, the aim of this study was to test: (1) whether mice show nostril asymmetries in
Absence of hyperplasia in Gasp-1 overexpressing mice is dependent on myostatin up-regulation Caroline Brun, Luce Périé, Fabienne Baraige, Barbara Vernus, Anne Bonnieu,
MBLR matrices can be represented using the cluster tree modelization typically used in the H literature; this provides a convenient tool to formalize the key difference between the
Access and use of this website and the material on it are subject to the Terms and Conditions set forth at The salinity of artificial built-up ice made by successive floodings of
stars near the Sun, indicates that sometime in the late Quater- nary the Sun, which has been moving through the very low density region known as the Local Bubble, encountered
Indeed, GBM6 cells acquired an endothelial phenotype when co-cultured with human endothelial cells, as a typical capillary network including fluorescent cells (GBM6-GFP cells)
(E) Histological scores of infiltrating cells in muscle and peripheral nervous system of recipient mice after transfer of T-cells isolated from muscles (10 6 cells) and
In mice, rhythmic expression of the Bmal1 gene e.g., in the liver appears unnecessary for core oscillator function and circadian rhythm generation [31].. However, in the SCN
