ENSAT – Ecole Nationale Supérieure Agronomique de Toulouse Avenue de l’Agrobiopole – 31326 Castanet-Tolosan
Projet de fin d’études pour l’obtention du diplôme d’ingénieur agronome
Validation de l’effet de huit régions chromosomiques sur la résistance au parasitisme chez quatre races ovines françaises
Sophie AGUERRE Spécialisation : Systèmes et produits de l’élevage
Promotion 2014/2015
Encadrant INRA :
Dr Carole Moreno Romieux INRA UMR1388 GenPhySE
Génétique, Physiologie, Systèmes d’Elevage B.P. 52627 31326 Castanet Tolosan
Remerciements
Je remercie tout d’abord l’INRA et en particulier le laboratoire GenPhySE pour m’avoir acceptée comme stagiaire et m’avoir donné l’opportunité de découvrir le monde de la recherche.
Je tiens à exprimer ma reconnaissance au Dr Carole Moreno-Romieux qui m’a encadrée tout au long de ce stage pour ses nombreuses explications en génétique, statistiques et programmation ainsi que pour ses conseils pendant la rédaction de ce mémoire.
Je remercie également mes tuteurs ENSAT, Zulma Vitezica et Hervé Rémignon pour avoir accepté de m’encadrer pendant ce stage ainsi que Véronique Gayrard qui a accepté de m’encadrer dans le cadre du master 2 Elaboration de la qualité et sécurité alimentaire.
Enfin je remercie les chercheurs, les doctorants et les techniciens pour leur accueil au sein du laboratoire GenPhySE ainsi que le personnel des domaines expérimentaux de La Fage et de la Sapinière pour leur accueil lors de mes déplacements aux domaines.
Résumé
Les parasites gastro-intestinaux constituent l’un des problèmes majeurs en élevage ovin. Le recours aux molécules anthelminthiques qui a longtemps représenté le principal moyen de lutte contre ces parasites est aujourd’hui remis en question. La sélection génétique d’animaux résistants constitue aujourd’hui une des stratégies complémentaires les plus efficaces pour la maîtrise de ces parasites. Toutefois, cela nécessite de mieux connaître les mécanismes impliqués dans la résistance au parasitisme chez les ovins. L’objectif de l’étude présentée ici est de confirmer la présence de QTL détectés par de précédentes études sur les chromosomes 3, 4, 5, 7, 12, 13, 14 et 21 chez quatre races françaises : Martinik Black Belly, Romane, Blanc du Massif Central et Manech Tête Rousse. L’expérimentation a consisté en deux infestations successives d’animaux âgés de six mois par Haemonchus contortus. Les animaux ont été génotypés et phénotypés pour l’intensité d’excrétion d’œufs (OPG), leur hématocrite et concentration en pepsinogène. Des SNP significatifs au seuil du chromosome ont été identifiés sur les chromosomes 3, 5, 7, 13 et 21 pour les OPG ; 3, 4, 5, 7,13, 14 et 21 pour l’hématocrite ; 3, 4, 12, 13 et 21 pour le pepsinogène.
Abstract
Gastrointestinal parasites are one of the main issue in sheep breeding. The use of anthelmintics molecules that has long been the main way to fight against these parasites is now being questioned. Genetic selection of resistant animals is nowadays one of the most effective complementary strategies to control these parasites. However, this requires a better understanding of the mechanisms involved in resistance to parasitism in sheep. The aim of the present study is to confirm the presence of QTL detected by previous studies on chromosomes 3, 4, 5, 7, 12, 13, 14 and 21 in four French breeds : Martinik Black Belly, Romane, Blanc du Massif Central and Manech Tête Rousse. The experiment consisted of two successive infestations of animals aged six months with Haemonchus contortus. Animals were genotyped and phenotyped for egg excretion intensity (FEC), hematocrit and concentration of pepsinogen. Significant SNP at the chromosome level were identified on chromosomes 3, 5, 7, 13 and 21 for FEC; 3, 4, 5, 7, 13, 14 and 21 to hematocrit; 3, 4, 12, 13 and 21 for pepsinogen.
Introduction
Les parasites gastro-intestinaux représentent aujourd’hui une véritable contrainte pour l’élevage des petits ruminants. Ils constituent une des causes d’importantes pertes de production voire de mortalité chez les ruminants conduits à l’herbe. Le parasitisme a donc un fort impact économique en raison des pertes de production, les coûts associés aux traitements et une surcharge de travail pour la gestion des troupeaux et des pâtures. L’utilisation des traitements anthelminthiques est depuis plusieurs années remise en cause du fait de l’apparition de résistances chez les parasites dans plusieurs régions du monde. Plusieurs stratégies complémentaires doivent aujourd’hui être mise en place pour la maîtrise de ces parasites : usage raisonné des traitements anthelminthiques, gestion des pâtures, amélioration des rations et amélioration de la résistance des individus.
Ce rapport traite de la sélection d’animaux génétiquement résistants qui représente une des solutions les plus durables.
Une première partie décrit les connaissances actuelles sur la résistance au parasitisme et les outils disponibles pour la sélection d’individus résistants. Un point sera en particulier fait sur les travaux portant sur la recherche des quantitative trait loci (QTL) associés à la résistance des animaux au parasitisme. Les QTL sont des régions du génome étroitement associées à un caractère quantitatif.
L’objectif de l’étude qui sera ensuite présentée est de confirmer la présence de QTL chez quatre races ovines françaises dans des régions précédemment identifiées. Pour cela, des expériences d’infestation ont été réalisées par l’INRA au sein de troupeaux de race Romane et Martinik Black Belly ainsi que par les organismes de sélection des races Romane, Blanc du Massif Central et des races ovines laitières des Pyrénées.
Partie 1 : Contexte bibliographique
Introduction
Les nématodes gastro-intestinaux représentent une préoccupation majeure pour les élevages ovins dans le monde. Les strongyloses gastro-intestinales entrainent d’importantes pertes économiques pour les élevages du fait d’une baisse de la qualité et de la quantité des productions. Par exemple, on observe une baisse de croissance chez les agneaux (Kyriazakis 1996) et donc une réduction des poids de carcasse à l’abattage associée à une moindre qualité de la viande. Au sein des troupeaux laitiers, les infestations par les nématodes gastro-intestinaux se traduisent par une baisse de la production de lait et une altération de la qualité (Chartier 1994, 1997). Le parasitisme a également des conséquences sur la production et la qualité de la laine (Knox 2006).
La maîtrise des strongles gastro-intestinaux grâce aux anthelminthiques ayant montré ses limites, de nouvelles stratégies de lutte sont étudiées. La sélection d’individus résistants permettrait de réduire l’utilisation des anthelminthiques. Actuellement, on se base sur l’estimation de l’intensité de l’excrétion d’œufs dans les fèces (Jacquiet 2009). Cependant, cette méthode est lourde à mettre en place et ne peut pas s’appliquer à de grands effectifs. L’utilisation des outils de génétique quantitative et de génomique est donc étudiée afin d’identifier des outils plus simples à mettre en œuvre : les mutations causales des gènes.
Tout d’abord, quelques éléments sur les nématodes gastro-intestinaux, leur cycle biologique et la réponse immunitaire déclenchée chez l’hôte sont présentés. La seconde partie traite des stratégies de lutte employées et met en évidence leurs limites. Enfin, les apports de la génétique sont abordés à travers une étude des variabilités inter et intra-races et un point sur les travaux de recherche concernant l’utilisation des QTL.
Chapitre 1 : Généralités sur les nématodes gastro-intestinaux des ovins
La majorité des espèces de nématodes gastro-intestinaux infestant les petits ruminants sont de l’ordre des Strongylida et de la famille des Trichostrongylidae, elle-même divisée en quatre sous-familles : Haemonchinae, Trichostrongylinae, Ostertagiinae et Cooperiinae.
En France, les principales espèces de nématodes sont Teladorsagia circumcincta et Trichostrongylus axei dans la caillette et Cooperia curticei dans l’intestin. La proportion des différentes espèces varient selon la saison (figure1). On note une plus grande part d’Haemonchus contortus en automne du fait des températures et d’une humidité élevées à cette période de l’année. Haemonchus contortus a été choisi comme modèle dans le cadre de cette étude car il est le plus prolifique et le plus dommageable pour l’animal.
Figure : Proportions des différentes espèces de nématodes du tube digestif des brebis à l’herbe au printemps (à gauche) et en automne (à droite). (Source : Jacquiet)
Sous-chapitre 1.1. Le cycle biologique des Trichostrongles gastro-intestinaux
Le cycle biologique des Trichostrongles se compose de deux phases (figure 2) (Bowman 1999):
- une phase libre dans le milieu extérieur. - une phase parasitaire chez l’hôte.
La première étape de la phase libre est l’élimination dans les fèces des œufs pondus en grand nombre par les femelles dans la caillette ou dans l’intestin grêle de l’animal. L’excrétion des œufs (70-150 µm) est à l’origine de la contamination du pâturage. Ils donnent naissance à des larves de stade 1 (L1). Ces larves vont ensuite muer en larves de stade 2. Les larves se nourrissent de matières organiques et de micro-organismes contenus dans les fèces. Elles évoluent ensuite en larves de stade 3. A ce stade, les larves possèdent une gaine qui les protège mais les empêche de se nourrir. Elles sont également capables de se déplacer. La durée de la phase libre dépend des conditions de température et d’humidité. Elle dure généralement entre 8 et 10 jours (Belanger 2007, Roeber 2013).
La phase parasitaire débute par l’ingestion des larves L3 par l’hôte. Les larves perdent leur gaine lors de leur passage dans l’estomac. Puis elles migrent vers la caillette ou l’intestin grêle. Les larves subissent une nouvelle mue en larves L4 dans la muqueuse gastrique. Les larves L4 évoluent en adultes immatures (L5). Les adultes matures vivent dans la lumière du
tractus gastro-intestinal de l’hôte. La période entre l’ingestion des larves et la ponte des œufs par les femelles est appelée période pré-patente (Belanger 2007, Roeber 2013).
Figure : Cycle biologique des Trichostrongles gastro-intestinaux chez les ovins. (Adapté de Lacroux, 2006)
Sous-chapitre 1.2. Haemonchus contortus et pathologies associées
Haemonchus contortus est un parasite de la caillette des petits ruminants (ovins et caprins). Sa distribution géographique est mondiale.
Ces vers ont une taille comprise entre 1 et 3 cm et une couleur rouge. L’une des particularités de ce parasite est que les femelles produisent une grande quantité d’œufs (de 5000 à 7000 œufs par jour).
Ce parasite se nourrit du sang de l’animal à partir du stade L4. Il est à l’origine de pertes importantes car il cause une dégradation de l’état général de l’animal : laine de mauvaise qualité, perte de poids, altération de la fonction de reproduction. Les migrations des larves et des adultes provoquent des hémorragies de la muqueuse de la caillette. L’animal soufre souvent d’anémie qui est une cause importante de mortalité, en particulier pour les agneaux de pâturage. L’anémie se traduit par la pâleur des muqueuses. L’animal peut aussi présenter un œdème dit de « la bouteille » (Belanger 2007, Bouharaoua 2012).
L’haemonchose existe sous trois formes (Chermette 1982):
- suraigüe : les animaux sévèrement infestés meurent subitement.
- aigüe : caractérisée par une anémie sévère et une perte de poids progressive.
- chronique : la plus fréquente et cause d’une morbidité élevée. Elle conduit à une dégradation de l’état général de l’animal et est donc à l’origine de pertes économiques importantes.
Sous-chapitre 1.3. Réponse immunitaire de l’hôte (cas des ovins) Il existe deux types de réponse immunitaire (Clark 2005, Tosi 2005) :
- la réponse immunitaire innée implique des mécanismes non spécifiques qui sont mis en place rapidement contre le pathogène.
Lors de l’infestation par les SGI, la muqueuse du tractus gastro-intestinal joue le rôle de barrière entre l’hôte et le parasite.
- la réponse immunitaire adaptative correspond à des mécanismes mis en jeu plus tardivement mais très spécifiques.
De nombreuses études s’intéressent aux étapes de la réponse immunitaire des ovins vis-à-vis des nématodes gastro-intestinaux. La réponse immunitaire mise en place est de type Th2 (lymphocytes T auxiliaires de type 2) (Terefe 2007). Ces lymphocytes sécrètent des cytokines nommées IL4 qui induisent la production d’immunoglobulines E, IL5 qui induisent la production d’éosinophiles, IL 9 et IL13. L’activation des lymphocytes induit également la production de mastocytes et de leucocytes au niveau de la muqueuse gastro-intestinale. Des immunoglobulines A et G1 spécifiques sont également produites. La variation du nombre de globules leucocytes explique un tiers de la variation du nombre de vers. La variation de la réponse IgA dans la muqueuse abomasale explique 40% de la variation de la longueur et de la fécondité des femelles (Jacquiet 2009). Le nombre d’éosinophiles dans la muqueuse de la caillette explique 35% de la variation de la longueur des femelles (Jacquiet 2009).
Ainsi, le cycle biologique des Trichostrongles gastro-intestinaux comprend une phase libre et une phase parasitaire. Leur cycle est dit monoxène car ils n’ont besoin que d’un hôte. Certains d’entre eux, tels que Haemonchus contortus (fréquent chez les ovins) ont une répartition mondiale et entrainent des pertes économiques importantes.
Chapitre 2. Les stratégies de lutte contre les strongles gastro-intestinaux et leurs limites
Trois principaux axes de lutte contre les nématodes gastro-intestinaux ont été définis : éliminer les strongles gastro-intestinaux, tarir les sources de contamination et augmenter la résistance de l’hôte.
Sous chapitre 2.1. Les traitements anthelminthiques et limites de leur utilisation
Trois familles d’anthelminthiques peuvent être utilisées pour lutter contre les nématodes gastro-intestinaux chez les ovins (Bouharaoua 2012, Lacroux 2006).
Les benzimidazoles agissent en empêchant la formation des microtubules du cytosquelette des parasites sans altérer ceux de l’hôte. Les multiplications cellulaires étant bloquées, les femelles adultes ne peuvent plus pondre. Enfin, les benzimidazoles provoquent des lésions au niveau des cellules intestinales et empêchent ainsi les strongles de se nourrir.
Les imidazothiazoles, en particulier le lévamisole, agissent comme un agoniste des récepteurs nicotiniques de l’acétylcholine au niveau des synapses des cellules musculaires des nématodes. Cela provoque une paralysie spastique qui conduit à la mort du parasite (Kohler 2001). Cependant, le lévamisole n’agit pas sur les larves inhibées.
Les lactones macrocycliques induisent une paralysie flasque des muscles somatiques chez le parasite du fait d’une augmentation de la perméabilité des cellules musculaires aux ions chlorures (Martin 1997).
Tableau : Principaux anthelminthiques actifs chez les ovins, noms déposés, posologies recommandées et temps d’attente à respecter. Pour le lait, les interdictions concernent les brebis laitières dont le lait est destiné à la consommation humaine. Légende : VO : voie orale, SC : sous-cutané, IM : intra-musculaire. (Adaptée de Lacroux 2006)
Famille Molécule Noms déposés Temps d'attente Résistance Monde Résistance France Benzimidazoles Oxfendazole Oxfenil® Viandes et abats : 14 jours Cawthorne E R.J. and Whitehead J.D., 1983 (Grande-Bretagne) Geurden et al, 2014 Synanthic® Lait : nul
Fenbendazole Panacur® Viandes et abats: 8 jours Lait : nul Fébantel Rintal® Viandes et abats : 10 jours Lait : nul Albendazole Valbazen® Viandes et abats : 10 jours Lait : interdit Imidazothiazoles Lévamisole Lévamisole®
Viandes et abats : 3
jours Le Jambre L.F. et
al, 1978
Chartier et
al, 1998
Salicylanilides
Closantel Supaverm®
Viandes et abats : 28 jours
Van Wyk J.A., Malan F.S., 1988 (Afrique du Sud) Seponver® Lait : interdit
Nitroxinil Dovenix®
Viandes et abat : 28
jours Cezar A.S. et al, 2010 (Brésil) Lait : 10 traites Lactones macrocycliques Ivermectine Ivomec® Viandes et abats : 3 jours Hughes P.L. et al, 1991 (Nouvelle-Zélande) Paraud et al, 2014 Oramec® Lait : interdit
Doramectine Dectomax® Viandes et abats : 35 (IM) ou 56 (SC) Borgsteede F.H. et al, 2007 (Pays-Bas) Lait : interdit
Moxidectine Cydectine® Viandes et abats : 3 jours Love S.C. et al, 2003 (Australie) Paraud et al, 2014 Lait : interdit
L’utilisation de traitements anthelminthiques est aujourd’hui discutée principalement du fait de l’apparition de résistances chez les nématodes gastro-intestinaux qui remet en cause leur efficacité. De plus, ces molécules peuvent être toxiques pour la microfaune du sol responsable de la dégradation des fèces des ruminants dans les prairies. Elles posent également problème en santé humaine avec la présence de résidus médicamenteux dans les denrées alimentaires d’origine animale.
En 1957, l’OMS a défini la chimiorésistance comme « l’apparition dans une population de la faculté de tolérer des substances toxiques à des doses qui exerceraient un effet létal sur la majorité des individus composant une population normale dans la même espèce ».
On distingue plusieurs types de résistance. On parle de résistance simple lorsque les parasites résistent à une substance unique, de résistance de famille lorsqu’elle concerne un groupe de substances ayant le même mode d’action, et enfin de résistance multiple pour une résistance à plusieurs familles différentes (Berrag 2008, Euzeby 2008).
La résistance est la conséquence d’une préadaptation par mutation. Son déterminisme est génétique. Les individus résistants, au départ peu nombreux, sont sélectionnés du fait de la pression environnementale (utilisation des anthelminthiques dans ce cas). Le cycle biologique des nématodes gastro-intestinaux étant court, le nombre d’individus résistants devient rapidement important (Euzeby 2008).
Plusieurs facteurs favorisent l’apparition de résistance chez les parasites (Euzeby 2008, Menzies 2010).
Une fréquence d’utilisation élevée d’un anthelminthique mène à une pression de sélection élevée et favorise donc le développement des résistances. Le risque est maximal lorsque la fréquence de traitement correspond à la période pré-patente du parasite. Dans ce cas, chaque génération est soumise au traitement (Berrag 2008).
Des anthelminthiques à libération prolongée ont été mis au point afin de diminuer le nombre de manipulations des animaux. Cette méthode est efficace mais peut induire une pression de sélection permanente.
Les erreurs de dosage favorisent également l’émergence de parasites résistants. Un sous-dosage exerce une pression de sélection sur la population en n’éliminant que les individus les plus sensibles. Le surdosage va, lui, entraîner l’apparition d’individus extrêmement résistants. Enfin, l’introduction d’animaux porteurs de parasites résistants dans un troupeau est un facteur de risque important.
La résistance aux anthelminthiques a pris une importance d’ampleur mondiale. Ce phénomène est en augmentation sur le plan géographique ainsi que par le nombre d’espèces de nématodes résistants et le nombre de molécules impliquées. Les ovins et les caprins sont particulièrement touchés. Les pays de l’hémisphère Sud sont particulièrement concernés (Australie, Nouvelle-Zélande, Afrique du Sud, Amérique du Sud). Le climat y est favorable et le développement d’un élevage intensif a favorisé ce phénomène (Waller 1997). Les pays européens sont moins touchés mais pas épargnés. En France la résistance d’H.contortus aux benzimidazoles a pu être mise en évidence chez les ovins dans plusieurs régions.
Ainsi le nombre de familles efficaces contre les strongles est de plus en plus limité. Des stratégies complémentaires doivent être employées pour lutter plus efficacement contre ces parasites
Sous chapitre 2.2. Limiter la contamination
Deux méthodes permettent de réduire le contact entre les animaux et les larves infestantes.
D’une part, la relation hôte-nématode est relativement spécifique. Cela permet d’assainir les parcelles grâce à une gestion raisonnée des pâturages. L’éleveur peut par exemple pratiquer un pâturage mixte ou alterné entre différentes espèces (ovins/bovins ou caprins/bovins). On peut également assainir une parcelle par une mise au repos et le retournement par labour (Kyriazakis 1996, Menzies 2010).
D’autre part, les capacités nématophages de certaines bactéries et champignons ont été mises en évidence. Par exemple, Duddingtonia flagrans peut être utilisé pour contrôler les parasites. Le mycélium du champignon immobilise et envahit les larves présentes dans les matières fécales. Larsen et al (1998) ont mené un essai dans lequel des ovins étaient infestés expérimentalement par plusieurs espèces de nématodes. Une réduction du nombre de larves infestantes dans les fèces de plus de 80% pour les animaux ayant reçu par voie orale des spores de D.flagrans.
La capacité de B.thuringiensis à détruire les nématodes aux stades larvaires ou adultes a également été étudiée in vitro pour trois espèces de nématodes H. contortus, T.colubriformis et T.circumcinta (Kohler 2001).
Sous-chapitre 2.3. Augmenter la résistance de l’hôte 2.3.1 La vaccination
La vaccination contre les nématodes gastro-intestinaux a été assez peu étudiée, à part pour Haemonchus contortus. L’utilisation de vaccins serait complexe à mettre en place. Il faudrait développer un vaccin pour chaque espèce de nématodes, ce qui compte tenu de leur nombre, paraît difficilement réalisable. De plus, les vaccins agiraient contre une molécule particulière chez le parasite qui pourrait donc développer des stratégies de contournement.
2.3.2 La nutrition
- Enrichissement de la ration en protéines
Il a été montré que des supplémentations en protéines permettent d’améliorer la résilience des animaux infestés du fait d’une amélioration de la réponse immunitaire de l’hôte. Des agneaux ayant reçu une ration enrichie en protéines présentaient des vers adultes plus courts que les agneaux non supplémentés. Les vers femelles adultes mesuraient 2,00 cm chez les agneaux supplémentés en protéines contre 2,13 cm chez le lot témoin. Une corrélation significative entre la diminution de la taille des vers femelles adultes et l’augmentation de la production d’immunoglobuline A. L’IgA agirait en affectant la fécondité des femelles et la taille des vers (Strain and Stear 2001). Chez les caprins, Bambou et al. (2011) ont montré qu’une ration enrichie en concentrés permet d’améliorer la résistance à H.contortus. Les chevreaux ont été répartis en quatre lots en fonction de la ration qu’ils recevaient (pas de concentré, 100g de concentrés, 200g de concentrés ou 300g). Chaque lot comprend autant d’animaux sensibles que résistants. Les animaux n’ayant pas reçu de concentrés présentaient une excrétion d’œufs dans les fèces deux fois plus importante que les animaux complémentés. Dans le lot non complémenté, les animaux sensibles présentaient une excrétion d’œufs dans
les fèces 1,6 fois plus importante que les animaux résistants. Aucune différence entre résistants et sensibles n’a été observée au sein des lots complémentés.
- Plantes à tanins
Les tanins sont des métabolites secondaires des plantes ayant un rôle de protection contre les prédateurs. On en trouve dans certaines plantes fourragères telle que le sulla (Hedysarium coronarium), le lotier pédonculé (Lotus pedunculatus), le lotier corniculé (Lotus corniculatus) ou le sainfoin (Onobrychis Viciaefolia). On distingue les tanins hydrolysables et les tanins condensés. Les tanins condensés ne traversant pas la barrière intestinale sont moins toxiques que les tanins hydrolysables.
Plusieurs études ont montré que la consommation de plantes riches en tanins était associée à une réduction du nombre d’œufs dans les fèces du fait d’une baisse de la charge parasitaire (nombre de vers) ou d’une diminution de la fertilité des femelles. Athanasiadou et al, (2000) ont montré une réduction des OPG de 25% au cours des cinq premières semaines d’expérimentation chez les moutons ayant reçu des tanins par rapport aux moutons n’en ayant pas reçus. Cette différence a atteint les 40% lors de la deuxième période de l’expérimentation. Ils ont constaté une diminution de la charge parasitaire d’environ un tiers entre les moutons ayant reçu des tanins et ceux n’en ayant pas consommés. Enfin la fécondité par femelle de Trichostrongylus colubriformis a été réduite de plus de moitié chez les animaux ayant reçu la plus forte dose de tanins en comparaison avec le lot témoin.
Différentes doses de tanins ont été testées. Au-delà de 8 à 10% de tanins condensés dans la matière sèche de la ration, des effets toxiques ont été observés chez les ovins.
Les modes d’action des tanins sont encore mal connus. Deux hypothèses sont faites :
- un effet direct : les tanins pourraient perturber l’intégrité de la cuticule des adultes ou de la gaine des larves infestantes.
- un effet indirect : les tanins condensés ont la capacité de former des complexes avec les protéines au niveau du rumen. Ces protéines d’origine alimentaire non dégradées au niveau du rumen seraient donc disponibles au niveau intestinal et constitueraient ainsi un apport supplémentaire de Protéines Digestibles dans l’Intestin d’origine Alimentaire(PDIA), ce qui stimulerait la réponse immunitaire de l’hôte. Cela permettrait de renforcer la résistance et la résilience de l’hôte vis-à-vis des parasites.
Ainsi, les produits anthelminthiques ont été largement utilisés et ont joué un grand rôle dans la lutte contre les strongles gastro-intestinaux. Leur utilisation est aujourd’hui remise en
cause du fait du développement de résistance au sein des populations de parasites et des préoccupations chez les consommateurs vis-à-vis des résidus de médicaments dans les aliments. Il faut donc mettre en œuvre d’autres stratégies afin de contrôler les populations parasitaires.
Chapitre 3. Apports de la génétique pour la maîtrise des strongles nématodes gastro-intestinaux
La sélection génétique ouvre de nouvelles perspectives dans la maîtrise contre les strongles gastro-intestinaux.
Sous chapitre 3.1. Utilisation en sélection
Généralement un petit nombre d’individus d’un troupeau porte une grande proportion de la population de parasites.
On distingue résistance et résilience. La résistance correspond à la capacité de l’hôte à réguler l’installation, le développement, la fécondité et la survie des nématodes. La résilience correspond à la capacité de l’hôte à maintenir son niveau de production tout en étant infesté.
3.1.1. Variabilité intra-race
Il a été mis en évidence que les mécanismes de résistance de l’animal sont aussi efficaces si celui-ci a été infesté de manière expérimentale que de façon naturelle. Cela peut être mis à profit dans le cadre de la sélection des reproducteurs (De la Chevrotière 2011). En effet, une infestation expérimentale permet de contrôler la dose de vers reçue par les animaux. En conditions naturelles, on ne peut pas savoir si les OPG d’un animal sont moins élevés que ceux des autres parce qu’il est plus résistant ou parce qu’il a ingéré moins de vers. En infestation expérimentale, on peut également contrôler de nombreux paramètres tels que l’âge, la ration ou la période de l’année par exemple. Autre point à souligner, il semble que les ovins ne présentent pas de spécificité de résistance vis-à-vis de la souche ou de l’espèce de parasite (De la Chevrotière 2011, Jacquiet 2009).
En vue d’intégrer la maîtrise des nématodes gastro-intestinaux aux schémas de sélection, plusieurs études ont essayé de quantifier la variabilité génétique au sein d’une même race à travers des estimations d’héritabilité. On appelle héritabilité la part moyenne de la variabilité phénotypique qui est d'origine génétique additive pour un caractère et une population donnée. L’héritabilité moyenne de la résistance aux strongles gastro-intestinaux
varie de 0,2 à 0,4. Ces héritabilités modérées nous permettent d’espérer une bonne efficacité de la sélection (De la Chevrotière 2011).
De plus, la plupart des études ont montré que les caractères de reproduction ou de production et ceux de résistance ne sont peu ou pas corrélés. Assenza et al. (2014) ont montré que les caractères de production et de résistance lors d’une primo infestation pouvaient être sélectionnés indépendamment. Cependant sur le long terme, une sélection sur les caractères de croissance en milieu non infesté pourrait conduire à sélectionner des animaux plus sensibles au parasitisme. Toutefois sélectionner les animaux en milieu infesté permettrait d’améliorer les caractères de croissance aussi bien que la résistance au parasitisme. On pourrait donc sélectionner les reproducteurs sur leur capacité de résistance sans dégrader les performances des animaux.
Ainsi, l’existence, au sein d’une même race, d’individus ayant des génotypes différents permet d’envisager une sélection d’animaux résistants aux strongles gastro-intestinaux. Cependant, même si les premières études sont plutôt encourageantes, il faudra veiller à ce que la sélection sur la résistance au parasitisme n’ait pas d’effet négatif sur les capacités de production des animaux ou sur la sensibilité à d’autres maladies.
3.1.2. Variabilité inter-race
De nombreuses études sur la variabilité inter-race de la résistance aux nématodes gastro-intestinaux ont été réalisées, aussi bien chez les espèces ovines que caprines.
Les races locales des pays tropicaux semblent plus résistantes que les races des régions tempérées. Cela peut s’expliquer par une sélection naturelle plus forte. En effet, ces animaux ont été élevés dans un milieu présentant une forte pression parasitaire. Cela leur permet de maintenir un bon niveau de production malgré la présence du parasite. Ces races ont donc une meilleure résilience (capacité à maintenir leur production). Les races de métropole sont, elles, plus sensibles et n’arrivent pas à maintenir une bonne productivité dans un milieu fortement parasité sans traitement (De la Chevrotière 2011).
Dans les pays des régions humides, ces résultats ont amené les éleveurs à choisir des races locales bien adaptées au milieu. Les animaux sont élevés en race pure ou en croisement terminal avec des races productives. Cependant, cette approche ne convient pas aux systèmes d’élevage ayant un niveau de productivité élevée (De la Chevrotière 2011).
D’autres stratégies pourraient être mises en place pour ces systèmes d’élevages si les gènes contrôlant la résistance au parasitisme étaient identifiés :
- par croisement
- par introgression : l’introgression génique consiste à inclure dans le génome d’une race sélectionnée A, un seul gène favorable G, d’une race B par ailleurs moins productive. Ceci s’effectue par un premier croisement A×B, suivi d’une série de croisements en retour (Backcross), des descendants portant le gène G, par la race A (Hospital 1992).
Cependant, elles nécessitent de maintenir des noyaux de race pure et d’introduire des béliers reproducteurs d’une nouvelle race. Elles demandent donc des investissements dans de nouvelles infrastructures et représentent un coût important. De plus, l’introgression est encore impossible à utiliser puisque les gènes responsables de la résistance au parasitisme ne sont pas connus.
3.1.3. Les critères de sélection
Afin de sélectionner les individus résistants, il faut définir un ou plusieurs indicateurs permettant de les identifier. Ces critères doivent être facilement mesurables, répétables et héritables. Le critère habituellement utilisé est le nombre d’œufs dans les fèces (Jacquiet 2009). Le comptage des œufs reflète le nombre de vers et permet donc mesurer le niveau d’infestation par les parasites. Cependant, l’excrétion des œufs dépend de plusieurs facteurs tels que le stade physiologique de l’hôte ou le stade de l’infestation (période pré-patente). D’autres critères permettant de mesurer la résilience peuvent être utilisés. L’hématocrite est également corrélé à la charge parasitaire puisque les vers se nourrissent du sang de l’hôte. Le pepsinogène est un indicateur des lésions de la caillette.
Sous-chapitre 2 : Recherche de QTL : état de l’art
Afin d’améliorer la sélection, des travaux de recherche sont menés sur l’utilisation de marqueurs moléculaires permettant de détecter les polymorphismes de l’ADN. Ces marqueurs sont des fragments d’ADN proches d’un gène d’intérêt. Ils servent de repère lorsqu’on suit la transmission d’un segment de chromosome d’une génération à l’autre. On s’intéresse en particulier aux Single Nucleotide Polymorphism (SNP). Il s’agit d’un polymorphisme d’un seul nucléotide qui correspond à une variation stable de la séquence d’ADN génomique portant sur une base affectant au moins 10% de la population.
Un QTL est une région chromosomique sur laquelle sont localisés un ou plusieurs gènes affectant potentiellement caractère quantitatif. La recherche des QTL permet de mettre en évidence des régions associées aux caractères même si on ne peut pas identifier et localiser précisément les gènes responsables.
Sous-chapitre 3 : Recherche de QTL affectant la résistance à Haemonchus contortus chez les ovins
Sallé G et al. (2012) ont mené une expérimentation pour identifier les QTL affectant la résistance à Haemonchus contortus. Une analyse de génome a été réalisée chez des agneaux issus d’un rétrocroisement Romane × Martinik Black Belly afin de profiter de la différence de résistance aux nématodes gastro-intestinaux des deux races (la Romane étant sensible et la Martinik Black Belly résistante). Les agneaux ont été exposés aux parasites lors de deux infestations expérimentales consécutives. Le dénombrement des œufs excrétés dans les fèces et l’hématocrite ont été mesurés. Un groupe de 332 agneaux (valeurs extrêmes pour le dénombrement d’œufs de parasites dans les fèces) a été autopsié afin d’étudier des paramètres tels que la charge parasitaire, la longueur des nématodes femelles, la proportion de mâles et de femelles. La concentration en pepsinogène a été mesurée dans un autre groupe de 229 agneaux.
La localisation des QTL a pu être précisée grâce au génotypage des animaux réalisé à l’aide d’une puce 50K (contenant 50 000 SNP répartis sur l’ensemble du génome). Des QTL ont pu être identifiés sur les chromosomes 5, 12, 13 et 21. Sur le chromosome 12, une région de 10 Mbp affecte le nombre d’œufs excrétés après la première et la deuxième infestation. Sur le chromosome 21, une région influe sur la concentration en pepsinogène et correspondant au locus du gène codant pour le pepsinogène.
En Ecosse, Riggio et al. (2013) ont mis en évidence des QTL associés à la résistance au parasitisme chez la race Scottish blackface. Les agneaux ont été infestés naturellement au pâturage. Le dénombrement des œufs dans les fèces, l’activité de l’IgA ainsi que leur poids ont été mesurés. Les agneaux ont aussi été génotypés à l’aide de la puce 50K. Les données ont été analysées grâce à deux méthodes. La première approche est une étude genome-wide association (GWA) qui teste chaque marqueur et regional heritability mapping (RHM) qui teste de petites régions du génome. Des QTL ont pu être mis en évidence sur les chromosomes 14 et 6, respectivement pour la résistance au genre Nematodirus et aux strongles.
Riggio et al. (2014) ont réalisé une méta-analyse à partir des données récoltées chez trois populations ovines : Scottish Blackface, Sarda×Lacaune et Martinik Black Belly×Romane. Cette étude a permis d’identifier des régions impliquées dans la résistance au parasitisme sur les chromosomes 4, 12, 14, 19 et 20 (genome-wide significant regions) et d’autres régions sur les chromosomes 1, 3, 4, 5, 7, 12, 19, 20 et 24 ayant un effet moins important (suggestive level).
Les résultats de ces expérimentations ont été mis en commun dans le cadre du projet Sustainable Solutions for Small Ruminants. Les principaux QTL en lien avec la résistance aux strongles gastro-intestinaux ont été détectés sur les chromosomes 4, 5, 7, 12, 13, 14 et 21.
Conclusion :
En conclusion, la maîtrise des strongles gastro-intestinaux passe par une combinaison de plusieurs stratégies : une utilisation des traitements anthelminthiques raisonnée, la gestion des pâturages, la nutrition ainsi que la sélection génétique qui représente un bon moyen de gérer durablement le parasitisme chez les ovins.
L’utilisation de puce SNP ouvre de nouvelles perspectives dans le domaine de la sélection génétique. Les SNP permettent d’identifier des marqueurs en lien avec la résistance aux strongles gastro-intestinaux. Ils permettent également de localiser les QTL impliqués, ce qui est très important pour identifier ensuite les gènes impliqués.
Lors de travaux de recherche récents, notamment ceux menés au sein du laboratoire GenPhySE, plusieurs QTL ont pu être localisés pour la race Romane et pour des animaux Romane × Martinik Black Belly. Les principaux QTL en lien avec la résistance aux strongles gastro-intestinaux ont été détectés sur les chromosomes 4, 5, 7, 12, 13,14 et 21.
Partie 2 : Contexte du stage
2.1. L’organisme d’accueilJ’ai réalisé mon stage de fin d’études à l’Institut National de la Recherche Agronomique (INRA) au sein du laboratoire GenPhySE. L’INRA est le premier institut de recherche dans les domaines de l’agronomie, l’alimentation et l’environnement en Europe. Il est le deuxième institut dans le monde pour ses publications en sciences agricoles.
Quelques chiffres clés permettent de mesurer l’importance de l’INRA :
- 19 centres de recherche en métropole et outre-mer,
- 208 unités de recherche, certaines associant l’INRA à d’autres organismes de recherche ou d’enseignement supérieur et à des instituts techniques,
- 51 unités expérimentales,
- 8 532 agents titulaires et 1891 doctorants encadrés.
Sept axes scientifiques sont développés au centre de recherche Toulouse Midi-Pyrénées dont les axes « Génétique et biologie animale intégrative » et « Santé animale et systèmes d’élevages durables » sur lesquels travaillent les chercheurs du laboratoire GenPhySE. Les recherches des équipes de GenPhySE concernent l’amélioration génétique des animaux d’élevage avec pour objectif une meilleure adaptation des animaux à leur environnement. L’équipe GeSPR au sein de laquelle j’ai réalisé mon stage travaille sur les caractères lainiers, les caractères de résistance aux maladies, le comportement maternel et l’efficacité alimentaire.
2.2. Problématique du stage
La résistance au parasitisme est l’un des caractères étudiés au sein du laboratoire GenPhySE. Ce caractère n’est pas encore pris en compte dans les schémas de sélection des petits ruminants en France. Cependant, la résistance aux anthelminthiques chez les parasites gastro-intestinaux ayant une ampleur mondiale, des stratégies complémentaires de lutte sont mises en place. Parmi ces stratégies, la sélection d’animaux génétiquement résistants est certainement la plus durable et la plus efficace. Pour pouvoir sélectionner les individus résistants, nous devons comprendre les mécanismes sous-jacents et identifier les régions chromosomiques mises en jeu (QTL). La méta-analyse réalisée par Riggio et al. en 2014 a mis évidence des régions associées à la résistance au parasitisme communes à trois populations chez les ovins. L’objectif du stage est de confirmer la présence de QTL sur ces régions chez quatre races française : Romane, Martinik Black Belly, Blanc du Massif Central et Manech Tête Rousse.
La faisabilité d’une sélection génétique des individus pour leur résistance au parasitisme avec les connaissances actuelles sera également discutée.
Partie 3 : Matériel et méthodes
Plusieurs études ont permis de mettre en évidence des QTL affectant la résistance au parasitisme chez les ovins. Les régions étudiées dans le cadre de notre étude ont été définies à partir des résultats obtenus par Sallé et al. (2012) et Riggio et al. (2014). Notre objectif est d’évaluer si on retrouve un effet de ces régions sur la résistance au parasitisme chez plusieurs races françaises : Romane (ROM_INRA et ROM_OS), Martinik Black Belly (MBB), Blanc du Massif Central (BMC) et Manech Tête Rousse (MTR).
3.1. Protocoles expérimentaux et populations étudiées
L’espèce Haemonchus contortus a été choisie car elle est la plus répandue au niveau mondial et la plus dommageable pour les animaux. De plus, les animaux résistants à cette espèce le sont également aux principales espèces de strongles. H.contortus constitue donc un
bon modèle pour d’autres parasites. Enfin, H.contortus ayant été utilisé pour de nombreuses expérimentations, les protocoles sont maintenant bien définis.
Le protocole d’infestation expérimentale par H.contortus réalisé sur les races Romane et Martinik Black Belly au domaine expérimental INRA de la Sapinière est décrit sur la figure 3. Trois caractères ont été mesurés : le nombre d’œufs dans les fèces (OPG), la concentration en pepsinogène et l’hématocrite. L’expérimentation comporte deux infestations réalisées à J0 suivies d’une période de 30 jours. A la fin de la première période, les animaux reçoivent un traitement suivi d’une période de repos de 15 jours.
A J0, des prélèvements sont réalisés pour vérifier l’absence d’excrétion d’œufs dans les fèces et mesurer le niveau de base pour le pepsinogène et l’hématocrite pour chaque animal. Les agneaux âgés de six mois sont ensuite infestés avec une dose de 10 000 larves de stade 3 (L3). Puis deux prélèvements sanguins sont réalisés à 14 et 30 jours après l’infestation pour le pepsinogène et l’hématocrite. A 24 et 35 jours après l’infestation l’intensité d’excrétion des œufs est mesurée. Les animaux sont ensuite traités avec de l’ivermectine (Oramec©, Merial) à la posologie de 0,2 mg/kg de poids vif. Le même protocole est suivi pour la deuxième infestation.
Figure 3 : Protocole d’infestation utilisé pour les populations Black Belly et Romane du domaine expérimental de Bourges.
Quatre races ont été étudiées, dont la Romane représentée par deux populations : une population issue du troupeau expérimental de la Sapinière, l’autre étant issue du troupeau de l’organisme de sélection (tableau 2). La race Martinik Black Belly, adaptée au climat tropical, présente une très bonne aptitude pour la résistance au parasitisme. La race Romane, issue du croisement entre les races Romanov et Berrichon du Cher, est plus sensible au parasitisme comme les autres races métropolitaines. Les données des autres populations ont été récoltées par les organismes de sélection de races confrontées au parasitisme : la Manech Tête Rousse
et la Blanche du Massif Central, deux races souvent menées à l’herbe. Le climat océanique tempéré des Pyrénées-Atlantiques est particulièrement favorable au développement des strongles gastro-intestinaux.
Les protocoles mis en place dans les organismes de sélection diffèrent en certains points de celui appliqué à la Sapinière. Les doses utilisées en station de contrôle individuel sont plus faibles que celles du protocole suivi à la Sapinière afin de préserver les performances des béliers. De plus, seuls deux prélèvements ont été effectués à J0 et J35 après l’infestation pour mesurer l’intensité d’excrétion des œufs et l’hématocrite. La concentration en pepsinogène n’a pas été évaluée.
Tableau 2 : Présentation des populations étudiées et des protocoles appliqués.
Race Dose phénotypésAnimaux génotypésAnimaux prélèvements par infestation)Caractère (nombre de
MBB 10 000 L3
H.contortus 79 79
OPG (3) Pepsinogène et hématocrite (3)
ROM_INRA H.contortus10 000 L3 277 274 OPG (3) Pepsinogène et hématocrite (3)
ROM_OS 3500 L3 puis 5000 L3 H.contortus 189 189 OPG et hématocrite (2) BMC 74 73 OPG et hématocrite (2)
MTR 3000 à 7500 L3 H.contortus 410 203 OPG et hématocrite (2)
3.2. Données génotypiques
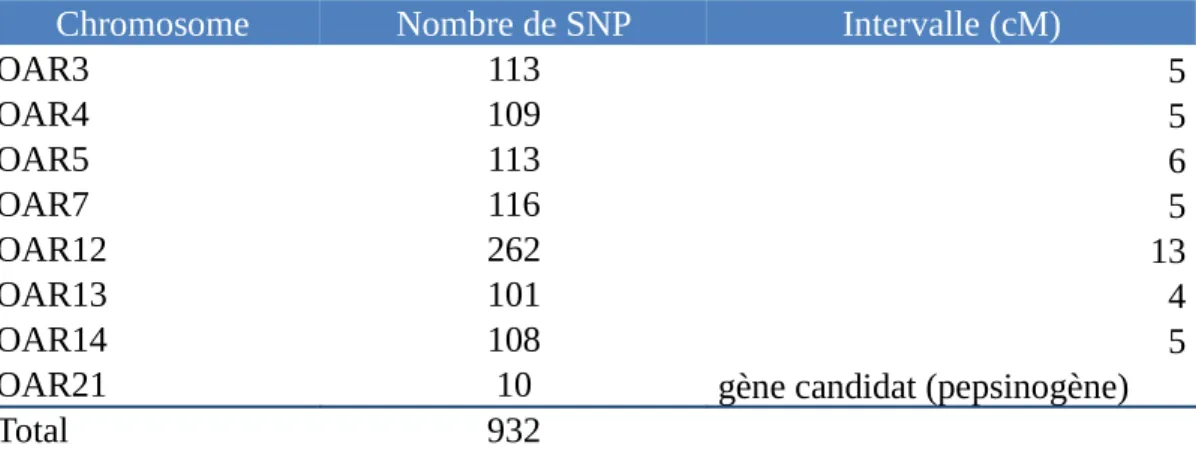
Neuf cent trente-deux marqueurs ont été utilisés pour la recherche des QTL sur 8 chromosomes (tableau 3). Une centaine de marqueurs ont été choisis sur 5 centimorgans (cM) autour de zones QTL identifiées par de précédentes études. Pour le chromosome 12, le nombre de marqueurs est plus important car deux régions QTL proches ont été identifiées
dans deux études. Pour le chromosome 21, seuls 10 marqueurs sont utilisés car on souhaite tester les SNP du gène contrôlant la production du pepsinogène.
Tableau 3 : Répartition des marqueurs utilisés lors des analyses sur les huit chromosomes étudiés. Le nombre de SNP étudiés est indiqué ainsi que la taille de l’intervalle sur lequel ils se trouvent.
Chromosome Nombre de SNP Intervalle (cM)
OAR3 113 5 OAR4 109 5 OAR5 113 6 OAR7 116 5 OAR12 262 13 OAR13 101 4 OAR14 108 5
OAR21 10 gène candidat (pepsinogène)
Total 932
Pour les Manech Tête Rousse, l’information génétique disponible est beaucoup plus importante puisque 39 874 marqueurs répartis sur 26 chromosomes ont été utilisés. Cela correspond à une dizaine de SNP par centimorgan.
3.3 Analyses statistiques
3.3.1 Analyses préliminaires des données
Pour chaque caractère, les données ont été analysées grâce à la procédure univariate de SAS® (SAS Institute, Cary, NC, USA) afin d’observer la distribution des données et d’estimer les moyennes et les écarts-types.
Les méthodes d’analyse utilisées par la suite sont basées sur l’hypothèse que les données suivent une distribution gaussienne. Lorsque ce n’était pas le cas, des transformations logarithme et racine quatrième ont été testées pour corriger les écarts à la normalité. La transformation racine quatrième a été retenue car elle a permis de se rapprocher au mieux d’une distribution gaussienne.
Les données ont été vérifiées afin d’écarter les valeurs aberrantes. Les valeurs s’écartant de la moyenne de plus de trois écarts-types sont considérées comme erronées et ne sont pas prises en compte par la suite.
3.3.2. Modèle linéaire
Plusieurs effets environnementaux tels que le sexe, l’âge, la taille de la portée, le mode d’allaitement ou les lots peuvent avoir un impact sur les caractères mesurés. Pour chacun d’eux, les animaux sont répartis en plusieurs niveaux. Grâce à une procédure FREQ de SAS®,
le nombre d’animaux a pu être calculé pour chaque niveau d’effet. Si certains niveaux comptent trop peu d’animaux (moins de 5), on les regroupe afin d’avoir des effectifs suffisants.
Avant d’effectuer l’analyse QTL, on souhaite corriger le modèle pour les effets environnementaux. Pour cela on cherche à identifier ceux qui ont un effet significatif sur les caractères mesurés. Les effets environnementaux ont été testés à l’aide d’une procédure GLM de SAS® dans le cas où ils étaient considérés comme fixes. La procédure GLM est basée sur le modèle suivant :
y = Xβ + ɛ
où y représente le phénotype, X la matrice relative aux effets fixes, β le vecteur des effets fixes et le vecteur des résidus du modèle.ɛ
Une procédure MIXED de SAS® a permis d’identifier les effets environnementaux significatifs dans le cas où l’effet père était considéré comme aléatoire. Le modèle mixed peut s’écrire selon l’équation suivante :
y = Xβ + Z + ɣ ɛ
avec y le phénotype, X la matrice relative aux effets fixes, β le vecteur des effets fixes, Z la matrice relative aux effets aléatoires, le vecteur des effets aléatoires et le vecteur desɣ ɛ résidus du modèle.
Ces procédures utilisent un test de Fisher pour tester la significativité des paramètres.
3.3.3. Qualité des données génotypiques
Il est aussi important de contrôler la qualité des données génotypiques. L’ADN peut être de mauvaise qualité, ce qui est à l’origine d’un grand nombre de données manquantes. Il faut donc identifier les SNP qui présentent un fort taux de données manquantes. Il faut également éliminer les individus pour lequel le taux de succès (call rate) est trop faible car on peut alors supposer que leur génotype n’est pas d’assez bonne qualité. On ne tient pas non plus compte des marqueurs pour lesquels la fréquence de l’allèle mineur (MAF) est trop faible. Dans notre étude, nous avons exclu les SNP pour lesquels la MAF était inférieure à 5% et le call rate inférieur à 90%.
Après ce contrôle, 780 SNP étaient disponibles pour la race Martinik Black Belly, 774 pour la population Romane INRA, 806 pour la population Romane OS et 815 pour la race Blanc du Massif Central. Les données génotypiques de la race Manech Tête Rousse ont été triées avant le début de mon stage, 39 874 SNP étaient disponibles.
3.3.4. Recherche des QTL
La détection des QTL est basée sur l’utilisation d’associations non aléatoires entre les loci des marqueurs et des QTL. On parle de déséquilibre de liaison. Si un marqueur apparaît comme significatif à la suite de l’analyse, cela signifie qu’il existe un QTL proche du marqueur en déséquilibre de liaison avec lui.
Avec le logiciel Muller, l’analyse est réalisée SNP par SNP. La méthode utilisée permet de contrôler le risque de première espèce sur l’ensemble de l’analyse en tenant compte de la relation entre les tests réalisés en chaque marqueur. La présence d’un QTL est ainsi testée en différentes positions du génome. Pour chaque SNP, le logiciel calcule un seuil de rejet pour l’hypothèse d’absence de QTL. Le GWAS est basé sur le modèle suivant proposé par le logiciel BLUPF90 (programme BLUPF90 de Ignacy Misztal et collaborateurs, University of Georgia):
y = 1µ + Xb + Zu + e
dans lequel y représente le vecteur des performances, µ l’effet moyen général à la population, b l’effet de l’allèle, X le vecteur des génotypes au SNP testé, Z la matrice d’incidence des performances sur les valeurs polygéniques, e la résiduelle et où V(u)=Aσ2u, A étant la matrice de parenté.
Le programme Muller utilise un fichier contenant les génotypes, un fichier pour les phénotypes, un fichier pour le pédigrée des animaux et un fichier carte indiquant la répartition des marqueurs sur les chromosomes étudiés. Pour chaque fichier phénotype, un fichier le décrivant est créé (annexe 1). On indique aussi l’héritabilité pour les caractères mesurés qui est de 0,3 dans notre cas.
3.3.5. Comparaison des résultats Emmax et Muller chez les Romanes de la Sapinière
Pour la population de race Romane de la Sapinière, les données avaient été précédemment analysées à l’aide de la méthode Emmax. Le modèle mis en œuvre par cette méthode estime l’effet des SNP en s’appuyant sur une matrice de parenté basée sur l’information génotypique. Dans le cadre de mon stage, j’ai ré-analysé ces données avec la méthode Muller qui utilise une matrice de parenté basée sur le pédigrée des animaux.
Les p-values obtenues avec les deux méthodes ont été comparées pour chaque SNP pour les moyennes des OPG en première et deuxième infestation (fropg1 et fropg2). Les SNP significatifs à 5% et pour lesquels la MAF était supérieure à 10% avec les deux méthodes ont été sélectionnés.
3.4. Sélection des lignées divergentes
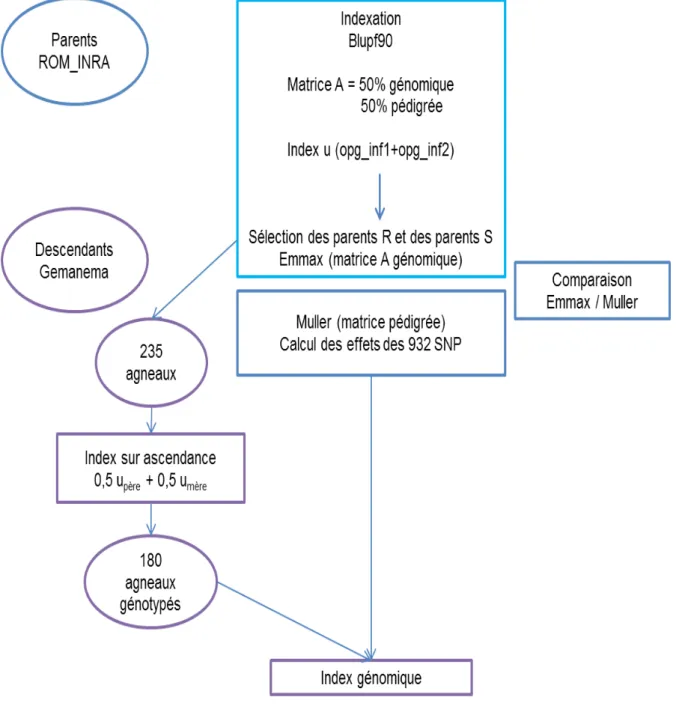
Un index u basé à 50% sur l’information génomique et à 50% sur le pédigrée a permis de sélectionner 6 mâles et 77 femelles extrêmes pour leur résistance au parasitisme parmi la population Romane de la Sapinière. Des accouplements entre les animaux résistants d’une part et les sensibles d’autre part ont été réalisés avant le début de mon stage. Deux cent trente-cinq agneaux sont issus de ces accouplements (99 issus des accouplements entre animaux sensibles et 136 issus des accouplements entre animaux résistants).
Lors de mon stage, nous avons sélectionné 180 agneaux parmi les 235 nés. Pour cela, un index sur ascendance a été calculé pour ces agneaux en attribuant à chacun d’eux la moitié de la valeur de l’index u de son père et la moitié de la valeur de u de sa mère. Ces agneaux ont été génotypés pour les 932 SNP.
Puis un index génomique a été calculé en utilisant comme population de référence leurs parents, phénotypés et génotypés pour les OPG en première et deuxième infestation. Les SNP ayant un effet significatif chez les parents avec la méthode de Muller sont standardisés (exprimés en écart-type phénotypique) et utilisés pour prédire la valeur génomique des descendants. L’index génomique est ensuite calculé pour chaque agneau en attribuant les effets des SNP standardisés calculés chez les parents aux agneaux en fonction de leur génotype. Ces index ont permis de sélectionner les agneaux les plus extrêmes pour le projet Gemanema (figure 4). Ce projet a pour objectif d’évaluer plusieurs caractères (analyse du comportement, métagénome par exemple) chez des lignées extrêmes sélectionnées pour leur résistance au parasitisme.
Figure 4 : Etapes suivies pour la sélection des lignées divergentes dans le cadre du projet Gemanema. Les cadres bleus correspondent aux étapes concernant la génération des parents (population Romane INRA). Le cadre bleu ciel contient les étapes qui ont été réalisées avant le début de mon stage. Les cadres violets représentent les étapes suivies pour la sélection de leurs descendants.
Partie 4 : Résultats
Les populations sont désignées dans les tableaux présentés dans cette partie par les abréviations suivantes :
- MTR pour la Manech Tête Rousse
- ROM_INRA pour la population de race Romane issue d’un troupeau du domaine de la Sapinière
- ROM_OS pour la population Romane issue d’un troupeau de l’organisme de sélection - BMC pour la race Blanc du Massif Central
4.1. Distribution des données
Les OPG présentent des écarts à la normale et des distributions asymétriques du fait d’un nombre important de valeurs nulles (figure 5). La transformation racine quatrième a été appliquée afin de les corriger (figure5). Des écarts à la normale ont également été observés pour la concentration en pepsinogène. Ils ont été corrigés de la même manière. Pour la population Romane de la Sapinière, les données brutes de concentration en pepsinogène pour les deuxièmes prélèvements en infestation 1 et 2 (peps2 et peps4) présentaient une distribution proche d’une distribution gaussienne. Aucune transformation n’a été appliquée à ces données. Figure 5 : Graphiques représentant la déviation à la normalité des OPG de la population Romane de Bourges à 24 jours après la première infestation avant (graphique de gauche) et après la transformation (graphique de droite).
Pour l’hématocrite, les données brutes présentent une distribution très proche d’une distribution gaussienne (figure6). Aucune transformation n’a été nécessaire.
Figure 6 : Graphique représentant la déviation à la normalité de l’hématocrite de la population Romane de Bourges à 24 jours après la première infestation.
Plus de deux cents individus ont été pris en compte dans l’analyse pour les races Romane (INRA de Bourges) et Manech Tête Rousse avec respectivement 265 et 205 animaux. Pour les autres races, les données d’une centaine d’individus étaient disponibles : 129 pour la
race Romane (OS) et 104 pour la race Blanc du Massif Central et 79 individus pour la race Martinik Black Belly.
Après avoir éliminé les valeurs aberrantes, le nombre d’animaux conservés par race est de 194 pour la Manech Tête Rousse, 265 pour la Romane (INRA), 102 pour la Romane (OS), 70 pour la Martinik Black Belly et 102 pour la Blanc du Massif Central.
Le tableau 4 présente les principales caractéristiques des données collectées chez les animaux de race Romane (INRA). Ces résultats sont décrits pour les autres races en annexe 2.
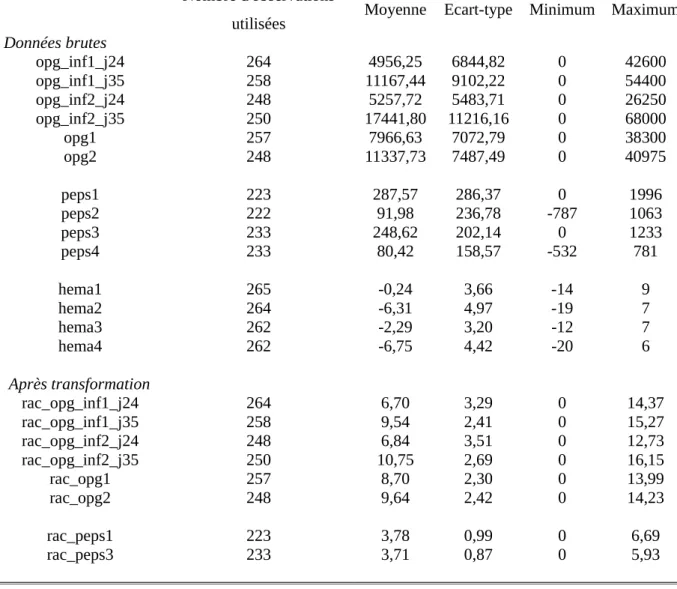
Tableau 4 : Statistiques descriptives pour les OPG, le pepsinogène et l’hématocrite avant et après transformation pour la race Romane (INRA). Le tableau présente le nombre d’animaux utilisés pour les calculs, la moyenne, l’écart-type et les valeurs extrêmes pour chaque caractère mesuré.
Romane - INRA de Bourges
Nombre d'observations
utilisées Moyenne Ecart-type Minimum Maximum
Données brutes opg_inf1_j24 264 4956,25 6844,82 0 42600 opg_inf1_j35 258 11167,44 9102,22 0 54400 opg_inf2_j24 248 5257,72 5483,71 0 26250 opg_inf2_j35 250 17441,80 11216,16 0 68000 opg1 257 7966,63 7072,79 0 38300 opg2 248 11337,73 7487,49 0 40975 peps1 223 287,57 286,37 0 1996 peps2 222 91,98 236,78 -787 1063 peps3 233 248,62 202,14 0 1233 peps4 233 80,42 158,57 -532 781 hema1 265 -0,24 3,66 -14 9 hema2 264 -6,31 4,97 -19 7 hema3 262 -2,29 3,20 -12 7 hema4 262 -6,75 4,42 -20 6 Après transformation rac_opg_inf1_j24 264 6,70 3,29 0 14,37 rac_opg_inf1_j35 258 9,54 2,41 0 15,27 rac_opg_inf2_j24 248 6,84 3,51 0 12,73 rac_opg_inf2_j35 250 10,75 2,69 0 16,15 rac_opg1 257 8,70 2,30 0 13,99 rac_opg2 248 9,64 2,42 0 14,23 rac_peps1 223 3,78 0,99 0 6,69 rac_peps3 233 3,71 0,87 0 5,93
Chez les races Romane (INRA) et Martinik Black Belly, le nombre d’œufs dans les fèces augmente entre J24 et J35 lors des deux infestations. Les moyennes calculées pour la variation de l’hématocrite sont, de manière générale, négatives chez toutes les races alors qu’elles sont plutôt positives pour les variations de concentration en pepsinogène. Ces observations correspondent bien à ce qui a pu être décrit dans la bibliographie pour ces caractères. La race Martinik Black Belly présente deux à trois fois moins d’œufs dans les fèces que la race Romane (INRA) en première et deuxième infestations (rac_opg1 et rac_opg2). Les mesures d’hématocrite sont en moyenne plus élevées pour la race Martinik Black Belly. Là encore, nos données sont en accord avec les résultats obtenus pour ces races par de précédentes études. Cela nous permet de penser que les données phénotypiques collectées sont fiables. Pour la race Manech Tête Rousse, quatre protocoles d’infestation différents ont été suivis. L’utilisation de différentes doses a eu un impact très fort sur les caractères mesurés. Les moyennes et écarts-types ont donc été calculés pour chaque protocole. La figure 7 présente les distributions obtenues en première infestation pour chaque protocole. Des différences importantes entre les moyennes et les écarts-types calculés pour chaque protocole peuvent être notées.
Figure 7 : Distribution observée pour les OPG en première infestation chez la race Manech Tête Rousse (protocole 1 en haut à gauche, protocole 2 en haut à droite, protocole 3 en bas à gauche et protocole 4 en bas à droite.
4.2. Corrélations phénotypiques entre les caractères.
Les corrélations phénotypiques entre les caractères d’infestation sont présentées dans le tableau 5 pour la race Romane (INRA).
Nous avons obtenu de bons coefficients de corrélations phénotypiques entre les OPG en première et deuxième infestation (0,39) et entre les mesures de l’hématocrite hema1 et hema2 pour la première infestation (0,39) et hema3 et hema4 en deuxième infestation (0,48).
Les corrélations entre les OPG et le pepsinogène sont généralement positives. Elles sont au contraire plutôt négatives entre les OPG et l’hématocrite et entre le pepsinogène et l’hématocrite. Cependant les coefficients de corrélation obtenus sont plutôt faibles.
Les mêmes tendances ont été observées chez les autres races (annexe 3).
Tableau 5 : Corrélations phénotypiques estimées entre les caractères pour la race Romane (INRA de Bourges). Opg inf1 Opg inf2 rac_ peps1 peps2 rac_
peps3 peps4 hema1 hema2 hema3 hema4 fropg1 -0,39 *** NS NS NS NS -0,17 ** -0,61 *** -0,13 * -0,26 *** fropg2 - NS NS NS NS NS -0,21 *** NS -0,5 *** rac_ peps1 -0,27 *** 0,17 * NS NS NS NS 0,15 * peps2 - NS 0,29 *** NS NS NS NS rac_ peps3 -0,24 *** NS NS NS NS peps4 - NS NS NS NS hema1 - 0,39 *** NS NS hema2 - NS 0,24 *** hema3 - 0,48 *** hema4 -4.3. Effets environnementaux
Selon la population étudiée, différents effets fixes ont été testés afin de mesurer l’impact des effets non génétiques et/ou environnementaux sur les caractères mesurés et, si besoin, corriger le modèle utilisé pour la recherche des QTL.
Plusieurs paramètres étaient connus pour les races Romane et Martinik Black Belly infestées au domaine de la Sapinière. La taille de la portée, le sexe et les lots formés ont été testés chez les deux races. Le mode d’allaitement n’a pas pu être testé chez la Martinik Black
Belly car seulement quatre animaux sur 79 ont été allaités artificiellement. Chez la race Romane, le mode d’allaitement a eu un effet significatif sur les trois caractères mesurés lors de la première infestation (p-value de 0,0007 pour la moyenne des OPG en première infestation). Le sexe a eu un effet significatif en deuxième infestation chez les deux races (p-value<0,0001 pour la moyenne des OPG en deuxième infestation chez les deux races). Les lots d’infestation ont également eu un effet significatif en première et deuxième infestation pour les deux races.
Les données de la race Romane de l’organisme de sélection ont été collectées en 2011 et 2012. L’effet année a été considéré car les conditions d’infection (virulence de la souche notamment) et climatiques ainsi que l’alimentation ont pu varier d’une année à l’autre et avoir un effet sur les OPG et les mesures d’hématocrite. L’élevage d’origine a aussi été testé. Les analyses ont révélé un effet significatif de l’année en deuxième infestation pour les OPG (p-value de 0,01) mais pas de l’élevage d’origine.
Les béliers phénotypés de race Blanc du Massif Central sont issus de deux élevages. Ce paramètre a eu un effet significatif sur les OPG et l’hématocrite en deuxième infestation.
Les données phénotypiques de la race Manech Tête Rousse ont été collectées en 2008, 2009 et 2011 après infection par H.contortus. En 2008, deux protocoles d’infestation ont été testés : un protocole avec une dose de 5 000 L3 et un protocole avec une dose de 7500 L3. En 2009, Les béliers ont été infestés avec des doses de 3 500 L3 en première infestation et 5 000 L3 en deuxième infestation. En 2011, la dose était de 3 000 L3 en première et deuxième infestation. Des béliers d’âges différents ont été infestés au cours des trois protocoles. En plus du protocole d’infection incluant un effet année de mesure, l’année de naissance (millésime) a donc été testée. Les analyses ont montré un effet significatif du protocole sur les OPG et l’hématocrite en première et deuxième infestation (p-value<0,0001 pour les OPG en première infestation et l’hématocrite en première et deuxième infestation, p-value de 0,0041 pour les OPG en deuxième infestation). En revanche, l’âge des béliers n’a pas eu d’effet significatif sur ces deux caractères.
Les effets environnementaux pris en compte pour la correction des données des différents caractères mesurés sont présentés dans le tableau 6.
Tableau 6 : Effets environnementaux ayant un impact significatif sur les caractères mesurés (OPG, pepsinogène et hématocrite).
Race Infestation Caractère Effets environnementaux
MBB
1
OPG Père (J24), sexe (J35)
Pepsinogène Père, lot d'infestation
Hématocrite /
2
OPG Sexe
Pepsinogène Père, lot d'infestation
ROM_ INRA
1
OPG Mode d'allaitement, lot d'infestation Pepsinogène Mode d'allaitement, lot d'infestation
Hématocrite Mode d'allaitement
2
OPG Sexe, taille de la portée, lot d'infestation
Pepsinogène Lot d'infestation
Hématocrite Lot d'infestation, sexe
ROM_OS
1 OPG /
Hématocrite /
2 OPG Père, année
Hématocrite /
BMC 1 OPG, hématocrite
2 Elevage d’origine
MTR 1 OPG Père, protocole d'infestation
Hématocrite Protocole d'infestation
2 OPG Père, protocole d'infestation
Hématocrite Protocole d'infestation
En plus de ces effets, l’effet du père a également été inclus dans l’analyse comme effet aléatoire. La prise en compte d’un modèle mixte permet de tester les effets fixes dans un modèle incluant un effet génétique père afin de mieux approcher les modèles QTL utilisés par la suite.
Les pères des individus de race Romane infestés au domaine expérimental de la Sapinière n’étaient pas connus. Vingt-six pères ont pu être identifiés grâce à l’information moléculaire avant mon stage. Cependant certains pères étant plein-frères, il n’a pas été possible de retrouver le père de tous les animaux. Finalement, les pères de 216 animaux sur les 274 animaux génotypés sont connus. Toutefois le pédigrée étant partiel, l’effet père n’a pas pu être mesuré sur cette population.
Chez la race Blanc du Massif Central, le père n’a pas eu d’effet significatif sur les OPG et l’hématocrite. L’effet du père n’a été significatif sur l’hématocrite chez aucune des races. Le père a eu un effet significatif sur les mesures réalisées à J24 en première et deuxième infestation, avec un effet plus fort en première infestation chez la Martinik Black Belly. Enfin, les analyses ont montré un effet significatif du père sur les OPG en première et deuxième infestation selon les races. On note en particulier l’effet hautement significatif du père sur les OPG en deuxième infestation chez la Manech Tête Rousse (p-value<0,0006).
4.4. Recherche de QTL
4.4.1. Analyses réalisées avec le logiciel Muller
Dans un premier temps, les données ont été analysées grâce au logiciel Muller. Il permet d’exécuter des BLUP et ainsi obtenir les tests statistiques pour chaque SNP. Puis les seuils de rejet de l’hypothèse d’absence de QTL sont calculés sur chaque groupe de liaison. A la fin de l’analyse, un fichier solution indique pour chaque SNP le résultat du test, la p-value et l’effet du SNP.
Les résultats sont également présentés de sous la forme de deux graphiques :
- un quantile-quantile plot (QQ plot) qui représente la distribution des p-values observées par rapport aux p-values attendues
- un Manhattan plot représentant les p-values en fonction de la position du SNP.
Les figures 8 et 9 présentent un exemple de Manhattan plot et QQplot.
Figure 8 : Manhattan plot obtenu pour le chromosome 5 pour les OPG en première infestation chez la race Romane (INRA). Chaque point représente le résultat du test pour un SNP. La position des SNP est indiquée sur l’axe des abscisses. L’axe des ordonnées indique la significativité du test (-log10 (p-value)). Les points situés au-dessus de la ligne la plus basse correspondant au seuil de Muller à 10% sont considérés comme étant des SNP significatifs.