HAL Id: hal-02785641
https://hal.inrae.fr/hal-02785641
Submitted on 4 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Effet de variables locales et contextuelles sur
l’assemblage des communautés végétales en prairies
permanentes
Gabrielle Benquey
To cite this version:
Gabrielle Benquey. Effet de variables locales et contextuelles sur l’assemblage des communautés végé-tales en prairies permanentes. Biodiversité et Ecologie. 2017. �hal-02785641�
1
2016-2017
MASTER FAGE
Biologie et Ecologie pour la Forêt,
l’Agronomie et l’Environnement
Spécialité
FONCTIONNEMENT ET GESTION DES
ECOSYSTEMES
EFFET DE VARIABLES LOCALES ET CONTEXTUELLES
SUR L’ASSEMBLAGE DES COMMUNAUTES VEGETALES
EN PRAIRIES PERMANENTES
GABRIELLE BENQUEY
Mémoire de stage, soutenu à Nancy le 05/09/2017
Julien, Pottier Chargé de Recherche
Bruno, Ferry Enseignant chercheur
INRA Centre de Recherches Auvergne Rhône Alpes,
5 chemin de Beaulieu,
63000 Clermont-Ferrand.
2 Introduction
Selon Diaz et al. (2006), la biodiversité peut être considérée comme : « le nombre,
l'abondance, la composition, la distribution spatiale et les interactions des génotypes, populations, espèces, types et traits fonctionnels, et des unités de paysages dans un système donné. ». Selon l’évaluation des écosystèmes pour le millénaire (Millennium Ecosystem Assessment, 2005) la biodiversité fournit des services écosystémiques essentiels au bien-être humain, incluant des services de soutien (ex. formation du sol), d'approvisionnement (ex. production alimentaire), de régulation (ex. pollinisation) et culturels (ex. bénéfices récréatifs ou spirituels).
Pour les écosystèmes prairiaux en particulier, l'augmentation de la biodiversité permettrait d’améliorer la pollinisation (Orford et al., 2016), d’augmenter la production de fourrage (Fischer et al., 2008), de réduire les pertes de production en cas de sécheresse (Tilman et Downing, 1994), de diminuer le taux d'infection par les pathogènes fongiques foliaires (Fischer et al., 2008), de limiter les invasions de plantes exotiques (Fischer et al., 2008) et d'améliorer le cycle des nutriments (Fischer et al., 2008). La valeur financière des services apportés par les prairies a été estimée à environ 232$ par hectare et par an (Costanza et al., 1997). Enfin, la prairie permanente représente en 2015 près de 33% de la surface agricole utilisée en France (Eurostat, sans date), bien que ses
surfaces aient connues des évolutions importantes par le passé (Xiao et al., 2015). Dans ce contexte, l'enjeu de préservation de la biodiversité des prairies permanentes en France est de taille, aussi est-il nécessaire de comprendre par quoi et comment le niveau de biodiversité est modulé afin de mettre en place des pratiques agricoles durables permettant de garantir son maintien.
Dans cette étude, nous nous intéressons à la diversité taxonomique et fonctionnelle des prairies permanentes de la région Auvergne dans le cadre d’un projet de recherche (FLORIS). La diversité taxonomique d'une communauté correspond au nombre et aux proportions des espèces qui la composent. La diversité fonctionnelle rend, quant à elle, compte des valeurs de traits fonctionnels caractérisant les individus des espèces présentes. Une des définitions de la diversité fonctionnelle, largement utilisée, est : « la valeur et l'étendue des traits des espèces et organismes qui influencent le fonctionnement d'un écosystème » (Tilman, 2001) cité par Laureto et al., (2015). Le terme de trait fonctionnel sera par la suite défini comme une caractéristique mesurable à l’échelle d'une plante individuelle qui affecte indirectement les performances de cette dernière via ses effets sur sa croissance, sa reproduction, et sa survie (Violle et al., 2007). Si la biodiversité apporte des services écosystémiques c'est principalement du fait de ces caractéristiques fonctionnelles (distribution des valeurs de traits) des organismes composant les écosystèmes (Díaz et al., 2006). Si certaines études ont mis en évidence un lien entre le nombre d’espèces et la diversité fonctionnelle (Cornwell et al., 2006) cette corrélation n'est pas systématiquement observée dans des communautés naturelles ou semi-naturelles. Le contexte environnemental agit en effet comme un filtre promouvant des espèces présentant des caractéristiques fonctionnelles leur permettant de se maintenir dans les conditions environnementales locales (Diaz et al., 1998). Dans ce contexte, l'ajout de nouvelles espèces dans la communauté n'a pas nécessairement pour effet d'augmenter l'étendue des valeurs de trait fonctionnel (Schmid et al., 2002). Il est ainsi nécessaire de prendre en compte conjointement les dimensions taxonomique et fonctionnelle de la diversité pour comprendre les mécanismes d’assemblage des communautés (Münkemüller et al., 2012).
La productivité (quantité de biomasse produite par unité de surface en un temps donné) et les perturbations (destruction totale ou partielle de la biomasse au cours d'une période de temps
délimitée) représentent des facteurs pouvant influencer le niveau de diversité taxonomique et/ou fonctionnelle d'une communauté végétale prairiale (Louault et al., 2017; Loranger et al., 2016 ; Socher et al., 2013 ; Laliberté et al., 2013). C'est en modulant ces facteurs que les pratiques agricoles constituent des leviers d'action potentiellement efficaces pour accroître ou maintenir la biodiversité des prairies permanentes. La productivité des prairies peut ainsi être affectée par le niveau de fertilisation et le pâturage (Frank et al., 2002), et les perturbations sont induites par la fauche et le pâturage. Notre objet d'étude est la prairie permanente gérée. Nous cherchons à quantifier l'impact du niveau des perturbations et de productivité sur la diversité taxonomique et
3 fonctionnelle des communautés végétales prairiales. Diverses études ont présenté des hypothèses et résultats concernant les variations de la diversité taxonomique et fonctionnelle en prairie le long des gradients de perturbation et de productivité.
Le DEM propose des mécanismes et des patrons de richesse spécifique relative
Huston en 1979 propose le Modèle d’Équilibre Dynamique (DEM) qui a pour objectif de prédire un ensemble de patrons de richesse spécifique relative d’une communauté en réponse aux gradients concomitants de productivité et de perturbations. Le modèle est nommé modèle d'équilibre dynamique car pour éviter le phénomène d’exclusion compétitive et ainsi maintenir un niveau maximal de coexistence entre espèces, il est nécessaire que le système subisse des perturbations répétées dans le temps. Ces perturbations doivent être à la fois assez importantes pour limiter la densité des individus et freiner l'exclusion compétitive, mais pas trop importante afin que les populations puissent se maintenir malgré la destruction de la biomasse associée au régime de perturbations. Les mécanismes qui sous-tendent le DEM sont les suivants :
Dans un système où les ressources sont limitées, trois principaux mécanismes aboutissent à l'exclusion de certaines espèces, et ainsi à la diminution de richesse spécifique à l’échelle de la communauté : i) la mortalité dépendante de la densité d'individus (corrélation positive avec la productivité) : lorsque la densité est forte, les ressources disponibles doivent être partagées entre les individus de différentes espèces. Les parts des ressources sont inégalement réparties entre les espèces du fait de différences de stratégies d’acquisition des ressources. Ainsi, les espèces ayant les taux de croissance les moins importants finissent par avoir de moins en moins de ressources et être exclues par des espèces plus compétitives. Plus la densité est forte, plus la compétition est intense. L'extinction d'espèces peut également être due à la mortalité non dépendante de la compétition pour l'acquisition des ressources : elle est aussi liée ii) à la quantité de ressources disponibles non modulée par la présence de compétition (ex : les ressources captées par un individu peuvent être limitées par la pauvreté du sol en lui-même) et iii) aux perturbations lorsque ces dernières ont une intensité trop importante et/ou sont répétées trop fréquemment. Les espèces montrant un développement trop lent ou une reproduction trop faible pour compenser les pertes d’individus engendrés par une forte fréquence/intensité des perturbations ne pourront se maintenir.
La productivité étant liée positivement à la vitesse à laquelle les populations croissent et à la densité, elle détermine l’intensité de la compétition. Le DEM prédit que cet effet de la productivité peut être contrebalancé par l’effet du régime des perturbations qui à l’inverse, a pour effet de diminuer la densité d'individus par la diminution de biomasse qu’il occasionne (Huston, 1979). Cependant, le régime des perturbations, s’il est trop important, peut mener à l’exclusion d’espèces qui ne produisent pas assez de biomasse pour croître et se reproduire entre les perturbations afin d'achever leur cycle de vie. Le DEM prédit qu'un tel régime de perturbations peut être contrebalancé par une forte productivité qui aurait pour effet d'augmenter la vitesse de croissance des populations et leur permettre de se maintenir entre les perturbations.
Les conséquences de ces mécanismes sur la richesse spécifique sont résumées dans la figure 1. Lorsque le niveau de productivité est élevé et que le niveau de perturbation est faible, la richesse spécifique est supposée minimale. Le niveau de diversité peut augmenter soit en diminuant le niveau de productivité soit en augmentant le niveau de perturbations. Quelques études ont été
spécifiquement menées dans le but de tester empiriquement la justesse de ces prédictions du DEM. Certaines, notamment traitant d'écosystèmes prairiaux (Laliberté et al., 2013) et marins (Worm et al., 2002) ont permis de confirmer certaines prédictions du DEM tandis que d’autres, notamment
traitant de communautés d’algues, d’invertébrés (Svensson et al., 2007), de protistes et de bactéries (Scholes et al., 2005) n’ont pas pu valider les prédictions de ce modèle.
4 Figure 1 : Modèle de l’équilibre dynamique proposé par Huston (1979)
Réponses attendues de l’équitabilité
Le DEM se focalise sur un indicateur de diversité taxonomique, la richesse spécifique relative, or d’autres sont plus couramment employés pour décrire les communautés, tel que l'équitabilité (J). L'équitabilité n'est pas dépendante du nombre d'espèce mais des abondances relatives de ces dernières. Cependant, des résultats issus de simulations mathématiques permettent de soutenir la capacité du DEM à prédire des valeurs d'équitabilité, à l’exception des prédictions à des niveaux importants de perturbations (Svensson et al. 2012). Les mécanismes invoqués dans les variations d'équitabilité selon les valeurs de perturbations et de productivité sont les mêmes que pour le DEM mise à part dans le cas où les perturbations sont fortes. En effet, dans ce cas de figure, l'équitabilité se maintient à un fort niveau et ce pour n'importe quelle valeur de productivité. Il est alors supposé que les niveaux élevés de perturbations laissent des zones de substrat libres permettant l'établissement d’espèces spécialistes des milieux perturbés (qui sont capables d’une colonisation rapide du milieu ouvert) qui coexistent alors avec les espèces plus compétitrices présentes.
En prairie permanente gérées pour la production de fourrages, des résultats empiriques ont pu mettre en évidence des patrons correspondant à ceux prédits par le DEM. Ainsi, lorsque le niveau de production est intermédiaire et que le niveau de perturbation (utilisation de l’herbe) diminue jusqu'à une absence de perturbation (ex. abandon du pâturage), la richesse spécifique diminue (Louault et al., 2017, fig. 1b).
Réponses attendues de différentes métriques de diversité fonctionnelle
Le DEM proposé par Huston (1979) ne propose pas de prédictions sur la diversité fonctionnelle. Récemment, Loranger et al. (2016) ont proposé une extension des prédictions du DEM pour la diversité fonctionnelle en s'appuyant sur des connaissances acquises par ailleurs sur les conséquences de la compétition sur les traits fonctionnel d'une communauté. Nous nous
appuyons sur les mécanismes proposés par Loranger et al. (2016) pour poser nos hypothèses sur les variations attendues de diversité fonctionnelle en fonction des gradients de productivité et de perturbation. Loranger et al. (2016) considèrent que le volume fonctionnel d'une espèce peut
5 constituer une mesure simplifiée de l’espace de la niche qu'elle occupe (également appelé niche fonctionnelle, (Violle et Jiang, 2009)). Le parallèle niche-volume fonctionnel proposé dans cette étude nous apparait pertinent car des théories existent sur le lien entre les caractéristiques de la niche d’une communauté et le nombre d’espèces pouvant s’y maintenir. Dans cette étude, le volume fonctionnel occupé par l’ensemble des espèces d’une communauté (aussi appelé richesse
fonctionnelle FRic) est utilisé comme un descripteur de la diversité fonctionnelle et est estimé dans un espace fonctionnel qui comporte autant de dimensions que de traits étudiés. A l'intérieur de ce volume se trouve un certain nombre d'espèces traduisant une « densité » appelée « species
packing », ce qui constitue un second descripteur de la diversité fonctionnelle de la communauté. Le niveau de species packing est élevé lorsque la majorité des espèces est regroupée dans une zone restreinte du volume fonctionnel de la communauté, et faible lorsque les espèces sont réparties équitablement dans l’ensemble de ce volume. Les auteurs suggèrent que les variations du volume fonctionnel occupé seraient proches de celles prédites dans le DEM pour la richesse spécifique relative. Les auteurs suggèrent que deux mécanismes associés à la compétition, mais aux effets opposés seraient potentiellement à l’œuvre dans le cadre du DEM étendu aux patrons de diversité fonctionnelle : la limite de similarité et la dominance compétitive. La limite de similarité est basée sur le phénomène d’exclusion compétitive survenant lorsque la quantité des ressources -
généralement souterraines telles que l'eau ou les nutriments - est limitante. L'exclusion compétitive est supposée affecter préférentiellement des espèces dont les stratégies d'acquisition des ressources sont très proches, par exemple, présentant une même profondeur d'enracinement, utilisant la même forme d'azote au même moment. Ceci favoriserait une plus grande amplitude des valeurs de traits fonctionnels associés à l'acquisition de certaines ressources, ainsi qu'une équitabilité plus grande de distribution de ces traits dans l'espace, c'est-à-dire une tendance des valeurs de traits fonctionnel à diverger et une réduction du species packing. L’autre mécanisme est la dominance compétitive. Ce mécanisme est davantage associé à une compétition asymétrique, où les plus grands individus obtiennent une part disproportionnée des ressources par rapport à leur taille ce qui aboutit à l’exclusion des plus petits individus (Weiner, 1990). C’est généralement le cas pour la ressource lumineuse pour laquelle les espèces les plus hautes et dont le port est recouvrant interceptent la très grande majorité de cette ressource. Il en résulte une dominance d'espèces compétitives dans
l’acquisition de la ressource lumineuse ayant toutes une même stratégie (hauteur élevée) ce qui réduit théoriquement l'espace fonctionnel occupé. Il faut noter que de petites espèces ayant des SLA plus élevées peuvent aussi survivre dans ces conditions et coexister avec les espèces dominantes, mais ces petites espèces produisent comparativement moins de biomasse. Dans ce cas, peu d’espèces coexistent dans un petit espace fonctionnel et les valeurs de traits fonctionnels ont tendance à converger et le species-packing est à son maximum.
Replaçons-nous dans le cadre du DEM pour des niveaux de productivité faible, intermédiaire et élevé.
Lorsque le niveau de disponibilité des ressources est faible (Fig. 1a), seules les espèces tolérantes au stress peuvent se maintenir (Grime 1977, Table 1) et au fur et à mesure que l’intensité/fréquence des perturbations augmente, le nombre d’espèces tolérantes au stress pouvant aussi résister à une intensité croissante de perturbations diminue, et avec lui le volume de l’espace fonctionnel. Lorsque le niveau de disponibilité des ressources est à un niveau intermédiaire (Fig. 1b) et que le niveau de perturbation est bas, le phénomène de limite de similarité est à l’œuvre car une biomasse assez importante doit être approvisionnée (puisque peu consommée ou exportée) et les ressources puisées doivent être assez importantes. L'espace fonctionnel est alors assez grand. Lorsque le niveau des perturbations augmente, la biomasse aérienne est moins importante ce qui est aussi le cas des ressources nécessaires pour la maintenir. Le phénomène de limite de similarité n'est plus dominant, mais l'espace fonctionnel ne diminue pas pour autant. Certaines niches laissées à l'abandon sont en effet colonisées par de nouvelles espèces et le volume fonctionnel occupé par la communauté est alors supposé plus grand. Dans ce cas, les grandes amplitudes des valeurs de trait ne sont plus une nécessité pour survivre comme dans le cas de la limite de similarité, mais le résultat d'un surplus qui profite à de nouvelles espèces. Ainsi les espèces aussi bien proches qu'éloignées fonctionnellement peuvent cohabiter.
6 Enfin, lorsque le niveau des perturbations est fort, le volume fonctionnel est plus bas qu'en condition de perturbation faible car seules les espèces rudérales (possédant des caractéristiques de tolérance aux fortes perturbations) peuvent se maintenir. Les niches occupées sont restreintes où les conditions de sol ne sont pas trop pauvres pour maintenir les espèces rudérales (Grime 1977, Table 1), ce qui contribue aussi à la diminution de l'espace fonctionnel. Lorsque le niveau de disponibilité des ressources est élevé, la ressource limitante est alors fréquemment la lumière, dont l’acquisition est unidirectionnelle, c’est-à-dire que l’espèce occupant l’espace de la canopée peut s’accaparer la majeure partie de cette ressource. Dans le cas où il n’y a pas de perturbation la compétition pour cette ressource est potentiellement maximale et l’espace fonctionnel se réduit puisqu’une unique stratégie maximisant à l’obtention de cette ressource est favorisée. La compétition pour la lumière diminue avec l’augmentation du régime des perturbations, induisant ainsi une augmentation du volume fonctionnel car les espèces de grandes tailles, de petites tailles supportant l'ombre ou ne la supportant pas peuvent coexister.
Une étude apporte une confirmation d’une des situations décrites ci-dessus par des résultats empiriques. Louault et al. (2017) montrent qu’une diminution du régime des perturbations jusqu’à abandon total du pâturage, en prairie productive s’accompagne d’une diminution de la diversité fonctionnelle des valeurs de LDMC, SLA et hauteur reproductive (associé à des valeurs moyennes de SLA faibles et des hauteurs reproductives et de LDMC élevées à l'échelle de la communauté). Ce phénomène de filtrage des espèces hautes avec un LDMC fort en conditions peu perturbées a déjà été observée dans de précédentes études et pourrait s’expliquer par l’importance de la compétition pour la ressource lumineuse en absence de perturbations ((Janeček et al., 2013).
Intervention d’autres facteurs et mécanismes
Le DEM proposé par Huston (1979) se propose de prédire des valeurs de richesse spécifique relative et non absolue, c'est-à-dire qu'il prédit le nombre d’espèces qui coexistent en un lieu donné par rapport au nombre d’espèces présentes dans le pool d’espèces local (Huston, 2014). Le pool d’espèces local correspond à l’ensemble des espèces qui peuvent potentiellement coloniser et se maintenir dans les conditions physico-chimiques locales (Zobel, 1997). Ce pool inclue donc les espèces visibles dans la communauté locale mais également présentes sous forme de propagules seulement dans la banque de graines par exemple. Or il semble que la richesse spécifique de nombreuses communautés soient contrainte par la taille du pool d’espèces. L'étude de Myers et Harms (2009) montrent ainsi que l'ajout de graines dans une communauté, correspondant à l’augmentation artificielle du pool, a pour effet d'augmenter la richesse spécifique dans différents écosystèmes terrestres, y compris en prairies. Une étude menée par Butaye et al. (2001), dans un contexte paysager forestier fragmenté, montre que le paysage constitue une contrainte de dispersion des espèces (le paysage est en partie responsable du fait que le pool local constitue une portion réduite du pool présent à une échelle plus importante). Cette même étude montre qu'il existe aussi un effet des contraintes environnementales (pH, lumière, humidité du sol, disponibilité en
nutriments) sur la constitution du pool local, cette dernière contrainte étant cependant moins restrictive que la première. L’influence de l’hétérogénéité du contexte paysager est également connue pour affecter le pool local des espèces adventices en grandes cultures (Gaba et al., 2010). La température a par ailleurs des effets sur la disponibilité en eau (les site les plus chauds en été sont aussi plus secs que les sites plus froid en été), sur la longueur de saison de végétation (la saison de végétation des sites les plus chaud est plus longue et inversement pour les sites les plus froids) et sur la disponibilité en nutriments (les sites ayant les été les plus chauds ont aussi une disponibilité en nutriments plus importante (plus grande teneur du sol en carbone organique total, Gutiérrez-Girón et Gavilán, 2013). En conséquence, sur les sites les plus froids, les plantes présentent des
caractéristiques particulières. Les feuilles ont un LDMC plus élevé, comme attendu en réponse à une faible disponibilité en nutriment (Gross et al., 2007) les tailles de feuilles sont plus petites, ce qui confère des avantages sur le plan de la conservation des ressources (Niinemets et al., 2007), et favorise en partie les espèces de petite taille car le temps de développement des petites feuille est
7 plus court (Moles et Westoby, 2000). La température est donc un facteur contraignant des valeurs de traits fonctionnels des espèces. Le climat pourrait agir sur le pool d'espèces en tant que filtre
environnemental, l’environnement contraignant alors les valeurs de traits permettant aux organismes de vivre dans des conditions abiotiques particulières, affectant alors potentiellement la constitution du pool local. Ainsi, Chalmandrier et al. (2017) ont mis en évidence que ce filtrage
environnemental apparait plus marqué dans les conditions les plus contraignantes, notamment à des altitudes plus élevées auxquelles la durée de saison de croissance de la végétation est plus réduite. Les contraintes s’exerçant sur le pool d’espèces peuvent donc interférer avec les mécanismes locaux à l’œuvre dans le DEM au point de masquer leurs effets sur la diversité taxonomique et
fonctionnelle des communautés (Lessard et al., 2012).
Dans le cadre de cette étude sur la diversité des prairies permanentes gérées, nous testons ici deux hypothèses complémentaires :
1- Les niveaux de diversité taxonomique et fonctionnelle des prairies permanentes auvergnates varient en fonction de leur niveau de productivité et du niveau d’utilisation de l’herbe selon les prédictions du DEM et ses extensions à la diversité fonctionnelle par Loranger et al 2016 et à l’équitabilité par Svensson et al. 2012
2- Les diversités taxonomiques et fonctionnelles des prairies permanentes auvergnates sont influencées par les conditions pédoclimatiques et le contexte paysager qui les caractérisent.
Matériel et méthodes
Nous étudions 18 prairies permanentes réparties sur l’ensemble du Massif Central âgées d’au moins 20 ans. Leur localisation est présentée en annexe (fig. A1). J’ai pu participé à l’ensemble de la campagne de terrain.
1) Design d’observation 1-1) Sélection des sites
Les conditions climatiques, les caractéristiques pédologiques et le contexte paysager peuvent influencer la diversité locale via un phénomène de limitation du pool d’espèces, notamment du fait d’un filtrage environnemental abiotique ou de contraintes de dispersion des graines. Afin de discriminer l'effet de ces facteurs des effets locaux du régime de perturbations et de la productivité du site sur le niveau de diversité, nous avons sélectionné 18 prairies permanentes sur la base de deux gradients croisés décrivant d'une part le pédo-climat et d'autre part le contexte paysager. Concernant le gradient pédo-climatique, un indicateur synthétique a permis de distinguer des prairies d'altitude (plus froides et présentant une importante quantité de carbone et d'azote total dans le sol) des prairies de plaine présentant des caractéristiques relativement opposées. Concernant le contexte paysager, nous nous sommes intéressé à la proportion de prairies entourant chaque site dans un rayon de 3 km. Une étude préliminaire a montré que cette variable est très corrélée à la connectivité fonctionnelle (i.e. la probabilité du site à échanger des espèces prairiales avec le paysage qui l’entoure en tenant compte des capacités de dispersion des espèces). Nous avons ainsi discriminé des contextes dominés par la prairie permanente d'autres contextes où les surfaces de grandes cultures ou de forêts étaient dominants. Les prairies sélectionnées intègrent par ailleurs l’éventail des principales pratiques de gestion des prairies dans le Massif Central, notamment différents régimes de fertilisation et d’utilisation de l'herbe, à l'exception des prairies d'estives qui n’ont pas été retenues. En prairies permanentes, les pratiques agricoles associées à l’utilisation de l’herbe sont souvent peu variables d’une année sur l’autre. En d’autres termes, les prairies pâturées, fauchées ou à gestion mixte présentent généralement ce type de gestion depuis une période assez longue. Le chargement animal et le régime de fertilisation peuvent sensiblement varier entre années, du fait des conditions météorologiques et de la quantité d’herbe produite. Nous assumons cependant qu’un régime moyen de gestion existe pour chaque parcelle (en d'autres termes que les variations de perturbation et de
8 productivité entre les années sont négligeables à l’échelle de plusieurs décennies). Une discussion avec chaque exploitant(e) nous a permis une première caractérisation des pratiques qui sera complétée par une enquête approfondie à l'automne dans le cadre du projet FLORIS. 8 prairies étaient fauchées, 4 prairies étaient pâturées lors de courte période (une prairie ayant un chargement animal intermédiaire et 3 prairies avec un chargement animal fort) et 6 prairies étaient pâturées pendant de longues période (3 prairies ayant eu un chargement faible, 2 ayant eu un chargement intermédiaire et 1 ayant eu un fort chargement).
1-2) Dispositif d’étude
Sur chacun des 18 sites un exclos carré de 10 x 10 m a été positionné sur une zone dont la composition floristique apparaissait représentative de la parcelle agricole. La zone à l'intérieur de l'exclos, délimitée et entourée d'une clôture électrifiée en cas de pâturage, était ainsi indemne de toute perturbation liée aux pratiques agricoles réalisées sur la parcelle lors de la campagne de terrain. Les différents éléments du dispositif sont présentés en annexe 2. Quatre visites ont été réalisées sur chaque site au cours de la saison de végétation afin d'avoir une représentation de la dynamique de la biomasse produite par la communauté ainsi que de la dynamique d'utilisation de l'herbe à l'extérieur de l'exclos. Ces visites se sont échelonnées entre le début de la saison de végétation et le pic de biomasse. La somme des degrés jours de croissance (GDD) représentent la somme des degrés Celsius moyens par jour dont la température était supérieure à 0 depuis le 1er février (les GDD sont exprimés en °j). Cette méthode de calcul est utilisée par les chambres d’agricultures auvergnates pour caractériser l’avancement de la saison de végétation. Le début théorique de la saison de végétation se situe aux alentours de 400 °j et le pic de biomasse se situe aux alentours de 1200 °j en moyenne dans les prairies du Massif Central. Du fait d’une fin d’hiver particulièrement chaude nous n’avons pu débuter nos observations qu’à partir de 500 °j. Les visites se sont échelonnées comme suit :
- 1ere visite (V1) en moyenne à 510 °j (min : 430 ; max : 560 ; ET : 37) ; - 2eme visite (V2) à 670 °j (min : 530 ; max : 780 ET : 70),
- 3eme visite (V3) à 990 °j (min : 880 max : 1080, ET : 60), - 4eme visite (V4) à 1240 °j (min : 1120 max : 1350, ET : 70). Les GDD sont été calculées sur la base de la formule suivante:
𝐺𝐷𝐷𝑗 = ∑ max (𝑇𝑏𝑎𝑠𝑒; 𝑇𝑚𝑎𝑥𝑖 − 𝑇𝑚𝑖𝑛𝑖
2 − 𝑇𝑏𝑎𝑠𝑒)
𝑗
𝑖=32
Avec j compris entre 32 et 365 (on considère les jours julien à partir du 1er fevrier), Tmax et Tmin les températures maximale et minimale observées au jour j, et Tbase ici 0°C la température
minimale à partir de laquelle la végétation se développe (McMaster & Wilhelm, 1997). 1-3) Relevé botanique et des traits fonctionnels
Des relevés botaniques ont été effectués dans huit quadrats de 1m² (Qbot : Quadrat
botanique). Chaque quadrat est divisé en 100 mailles de 10 cm². Après tirage aléatoire, dix mailles ont été étudiées dans chaque quadrat. Les relevés ont été conduits lors de la visite 3 de manière exhaustive dans les dix mailles des huit Qbot, soit 80 relevés par site d'étude. Cette méthode a permis de renseigner la présence des espèces de chaque maille, et ainsi la fréquence d’occurrence de chaque espèce à l’échelle du site, en se basant sur sa proportion de présence par rapport à l'ensemble des occurrences de l’ensemble des espèces sur l’ensemble des 80 mailles étudiées. Dans la suite du texte, le terme d'abondance relative sera utilisé, plutôt que celui de fréquence d'occurrence relative. Les relevés botaniques dans les mailles ont été complétés par la liste des espèces présentes dans chaque quadrat (mais à l’extérieur des mailles marquées) et dans l'ensemble de l'exclos. Une faible valeur a été attribuée à ces espèces complémentaires afin de les inclure par la suite dans le calcul de
9 la richesse spécifique tout en limitant leur poids dans le calcul d'indicateurs pondérés par les fréquences relatives, tout particulièrement l'équitabilité taxonomique. Cela a permis d'obtenir respectivement la liste des espèces présentes par quadrat et par site au cours de la visite 3.
Plusieurs traits fonctionnels ont été mesurés afin de caractériser fonctionnellement les communautés prairiales, parmi lesquels les traits proposés par Westoby (1998) définissant le schéma LHS (Leaf-Height-Seed), à savoir la surface spécifique foliaire (Specific Leaf Area : SLA ; exprimé en mm²/mg) correspondant au rapport de la surface foliaire sur la masse sèche de la feuille, la hauteur de canopée à maturité (HC ; exprimé en mm), et la masse des graines (MG ; exprimé en mg). Nous avons également étudié la teneur en matière sèche des feuilles (Leaf Dry Mater Content : LDMC ; exprimé en mg/g) correspondant au ratio entre la masse sèche et la masse fraîche de la feuille. L'ensemble des traits précédemment cités ne sont pas corrélés significativement dans notre analyse. Ces traits fonctionnels sont renseignés pour l'ensemble des espèces représentant au moins 80% des fréquences d’occurrence relatives cumulées pour chaque site séparément. Cette limite a été choisie comme un compromis entre la qualité de la caractérisation fonctionnelle des communautés et la faisabilité des relevés (Pakeman et Quested, 2007). Les espèces sélectionnées font l'objet d'une collecte à l’intérieur de l’exclos au cours de la visite 3. Le traitement des échantillons a suivi les protocoles standards (Perez-Harguindeguy et al., 2013). Les individus sont collectés, autant que possible, de manière homogène dans l'exclos et au stade végétatif. Dix individus par espèce sont sélectionnés pour mesurer le SLA et le LDMC. Les mesures de hauteur végétative sont réalisées sur 20 individus matures par espèce, et ce durant les visites 3 ou 4, afin d’obtenir les valeurs maximales pour chaque espèce, dépendant de la précocité et de la vitesse de développement des espèces. La hauteur de canopée est celle qui est atteinte par l’extrémité haute de l'organe photosynthétiquement actif le plus haut. Pour l'ensemble de ces traits, la valeur moyenne par site sera utilisée dans les analyses présentées par la suite. Par ailleurs, la masse des graines moyenne de chaque espèce a également été renseignée via des données issues de la base de données LEDA (Kleyer et al., 2008), provenant des mesures effectuées par l'Université de Sheffield dans des conditions similaires pour toutes les espèces. Ce choix d'utiliser ces valeurs provenant de bases de données s'explique par la faible variabilité intraspécifique de ce trait, comparativement à sa variabilité interspécifique (Kazakou et al., 2014).
1-4) Quantification des variables environnementales
La productivité des prairies a été mesurée sur la base de la quantité de biomasse aérienne produite. Quatre zones de tonte de 50 cm x 50 cm à l'intérieur de l'exclos ont permis de mesurer la biomasse produite en absence de perturbations à chaque visite (voir schéma explicatif). Les tontes ont été réalisées à 5 cm au-dessus du sol. Les biomasses récoltées ont été séchées à l'étuve à 60°C pendant trois jours. Sur la base des biomasses sèches aériennes prélevées à chaque visite, il a été possible d'estimer une mesure de productivité, c.à.d. une quantité de biomasse produite par unité de surface durant une période de temps donnée. Dans notre cas, la productivité a été quantifiée comme la quantité de biomasse aérienne sèche produite dans l'exclos entre deux visites successives (V1-V2, V2-V3 et V3-V4). La moyenne des valeurs de productivité obtenues a été considérée comme un indicateur synthétique de productivité de la prairie au cours de la saison de végétation en absence de perturbation. Deux métriques, exprimées en termes de biomasse produite par m² pour 100°j (notée « P100GDD ») et
pour 15 jours (notée « P15j ») ont été comparées lors des analyses statistiques. La somme des GDD à
l’année basée sur les normales saisonnières de températures (NSGDD avec Tbase =5°C) est la variable
climatique utilisée dans nos modèles. Cette variable permet de caractériser la durée de saison de végétation, comme utilisé dans de précédentes études (Chalmandrier et al., 2017). La proportion de prairie autour des prairies étudiées (PP) est utilisée comme indicateur du contexte paysager.
1-5) Quantification des perturbations
Le régime des perturbations dans la parcelle lors de l’année en cours a été quantifié via un Taux d'Utilisation de l'Herbe moyen « TUH». Le taux d'utilisation de l'herbe correspond à une mesure de la proportion de la biomasse produite qui a été retirée du fait des perturbations (exportée via une
10 fauche, un ensilage, consommée du fait du pâturage). Nous ne disposons pas d'informations sur la biomasse présente juste avant et après chaque événement de perturbation, aussi nous ne pouvons pas calculer le TUH exact. Pour approcher au mieux la valeur du TUH, nous assumons que le gain de biomasse entre deux visites à l'extérieur de l'exclos – en conditions perturbées – est le même que celui à l'intérieur de l'exclos et calculons le TUH suivant la formule :
TUHViVi+1 = (Bint Vi+ 1 - Bint Vi + Bext Vi - Bext Vi+ 1) / Bint Vi + 1 - Bint Vi + Bext Vi
où i correspond au numéro de visite ; Bext Vi correspond à la biomasse récoltée à l'extérieur de l'exclos
en visite i ; Bint Vi correspond à la biomasse récoltée à l'intérieur de l'exclos en visite i. Le numérateur
est une quantification de la quantité de biomasse retirée par les perturbations entre deux visites et le dénominateur est la quantité de biomasse disponible entre deux visites (et qui pourrait être retirée par les perturbations).
Afin d'illustrer l'indicateur, supposons qu'entre les visites 1 et 2, une fauche ait été réalisée, peu après la visite 1. A l'intérieur de l'exclos, en absence de perturbation, la pousse de l'herbe a été continue et la biomasse mesurée BintV2 correspond à la biomasse présente en visite 1 BintV1 ainsi que
la pousse entre la visite 1 et la visite 2 (BintV2 - BintV1). A l'extérieur de l'exclos, en cas de perturbation,
la biomasse mesurée BextV2 est le reste de la biomasse après perturbation tenant compte de la pousse,
c'est-à-dire le résultat de la biomasse présente en visite 1 à l'extérieur de l'exclos BextV1 (celle-ci
pouvant être différente de BintV1), de la pousse potentielle estimée comme la biomasse produite à
l'intérieur de l'exclos (BintV2 - BintV1) en retirant ce qui a été prélevée du fait de la perturbation. Une
moyenne des taux d'utilisation de l'herbe pour les trois périodes a été calculée pour donner le taux d'utilisation d'herbe moyen (TUH). Le TUH est un taux compris entre 0 (lorsque qu'aucune biomasse produite n'a pas été consommée) et 1 (lorsque toute la biomasse produite a été consommée). Il arrive que ce taux soit parfois légèrement négatif car la biomasse produite hors exclos peut parfois être légèrement supérieure comparée à l'intérieur, ceci étant lié à l’hétérogénéité de la production au sein des parcelles. Dans ce cas le taux est ramené à 0.
2) Indicateurs de diversité 2-1) Diversité taxonomique
La richesse spécifique (S) correspond au nombre d'espèces présentes dans une communauté. Elle est estimée sur la base du nombre d'espèces observées à l'échelle du site de 10m x 10m. L'équitabilité constitue une mesure du degré de régularité de la distribution des abondances relatives des espèces dans la communauté. L'équitabilité (notée J) est le rapport de l'entropie de Shannon (H) sur l'entropie de Shannon lorsque l'équitabilité est maximale (Hmax):
où S correspond au nombre d'espèces dans la communauté ; ni correspond au nombre d'individus de
la ième espèce et N correspond au nombre d'individus total de la communauté. Lorsqu’un nombre
réduit d'espèces dominent la communauté, H tend vers 0 (et J tend vers 0), lorsque les espèces sont réparties équitablement, H tend vers ln(S) (et J tend vers 1).
11 2-2) Diversité fonctionnelle
Les métriques de diversité fonctionnelle ont été calculées pour chaque communauté, tenant compte soit de l'ensemble des traits (métrique multi-dimensionnelle à 4 dimensions, FRic, Qmulti,
mNNDmulti), soit en considérant chaque trait séparément (métrique uni-dimensionnelle, exemple pour
le SLA, RangeSLA, QSLA, mNNDSLA, CWMSLA. Dans la suite de ce paragraphe, nous décrivons la
méthode de calcul de chaque métrique.
Le FRic (Functional Richness) (Villéger et al., 2008) est une mesure du volume occupé par une communauté dans un espace multidimensionnel, c'est-à-dire le plus petit volume intégrant l'ensemble des espèces dans l'espace fonctionnel multidimensionnel considéré. Cette analyse s'appuie sur une analyse en coordonnées principales (PCoA) à 4 dimensions. Les espèces sont distribuées dans l'espace fonctionnel de la PCoA sur la base d'une matrice de distance Euclidienne. Les espèces sont ainsi espacées les unes des autres sur la base de leurs dissimilarités fonctionnelles. Afin de comparer les différentes communautés entre elles et de donner le même poids aux différents traits, les valeurs de traits associées à chaque espèce dans chaque communauté sont au préalable standardisées par l’étendue des valeurs de traits de l'ensemble des espèces observées dans toutes les communautés selon la formule : (valeur - min) / (max - min) (min-max scaling). Chaque valeur de trait est alors comprise entre 0 et 1.
Le Range correspond à l'étendue des valeurs de chaque trait, sans standardisation (Villéger et al., 2008) :
Rangetc = Val maxtc-Val mintc
où Rangetc correspond à l'étendue des valeurs du trait t dans la communauté c ; Val max correspond
à la Valeur maximale du trait ; Val min correspond à la Valeur minimale du trait
L'entropie quadratique de Rao (Q) (Rao, 1982) représente la distance fonctionnelle moyenne entre deux individus tirés au hasard avec remplacement dans la communauté. Cette métrique nous permet de rendre compte du degré de divergence des traits fonctionnels (voir Bernard verdier et al 2012). La distance fonctionnelle pondérée par l'abondance relative de chaque espèce est calculée pour chaque paire d'espèces. Cette métrique permet de tester les phénomènes de limite de similarité et de dominance compétitive en supposant que les exclusions compétitives peuvent avoir lieu entre les espèces aussi bien proche qu'éloignées fonctionnellement. La pondération par l'abondance permet de tenir compte de la dominance relative des espèces. La formule est :
où s correspond au nombre d'espèces représentant au moins 80% de fréquences d’occurrence relatives cumulées pour chaque site séparément. pi et pj correspondent respectivement à l'abondance relative
de l’espèce i et de l'espèce j dans la communauté ; dij correspond à une matrice de distance fonctionnelle entre chaque paire d’espèce i et j. La distance fonctionnelle entre chaque paire d’espèces est calculée comme la distance Euclidienne dans l’espace à 4 dimensions considéré. Avant de calculer l'entropie quadratique de Rao multi-dimensionnelle, les traits fonctionnels sont standardisés de la même manière que pour la métrique FRic. Pour chaque trait considéré séparément, aucune standardisation n'a été réalisée.
Le mNND (mean Nearest Neighbour Distance) permet de tester les phénomènes de limite de similarité (Stubbs et Wilson (2004) ont aussi utilisé cette métrique pour tester ce même phénomène) et de dominance compétitive. Le mNND est la distance fonctionnelle moyenne pondérée par l'abondance entre les plus proches voisins dans l'espace fonctionnel. La distance fonctionnelle entre chaque paire d’espèces est calculée comme la distance Euclidienne dans l’espace à 4 dimensions considéré. La pondération par l'abondance permet de tenir compte de la dominance relative des espèces. Cette métrique a été calculée dans l'espace à 4 dimensions (mNNDmulti) et aussi pour chaque
trait sur une dimension (mNNDSLA par exemple). Pour le mNND calculé dans l'espace à 4 dimensions
une standardisation par l'étendue est effectuée de la même manière que pour le FRic. Pour le mNND calculé pour chaque trait séparément, la standardisation par l'étendue n'a pas été effectuée.
12 Le CWM est la moyenne pondérée des valeurs d'un trait fonctionnel à l'échelle d'une communauté. Le CWM est calculé comme suit :
où pi est l'abondance relative de l'espèce i, s correspond au nombre d'espèces représentant au moins 80% des fréquences d’occurrence relatives cumulées pour chaque site séparément et traiti correspond
à la valeur moyenne du trait pour l'espèce i 3)Analyses statistiques
3-1) Analyses préliminaire sur les données brutes
La normalité des données brutes a été vérifiée via un test de Shapiro (pour les indicateurs Prod et TUH) et une inspection visuelle (pour les valeurs de traits). Une transformation ln (log naturel) a été appliquée pour la variable masse des graines (MG) dont la distribution des valeurs était largement asymétrique. Nous avons testé le niveau de corrélation entre les traits fonctionnels considérés. Aucun trait n'était fortement corrélé aux autres dans notre cas d'étude.
3-2) Sélection de modèles 3-2-1) Modèles généraux
Quinze modèles différents nous permettent de tester nos hypothèses. Nous présentons ci-dessous les deux modèles généraux intégrant l'ensemble des variables explicatives du niveau de diversité.
Le modèle général correspondant à l’hypothèse 1 s’écrit comme suit: Div = b + a1 Prod + a2 TUH + a3 Prod² + a4 TUH² + a5 Prod * TUH
(éq. 1)
où Div correspond à une métrique de diversité (S, J, FRic, mNND, Q, CWM), TUH au taux d'utilisation de l'herbe et Prod à une métrique de productivité (soit P100GDD, soit P15j).
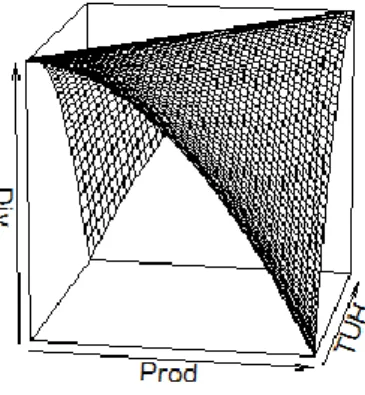
Ce modèle intègre un effet quadratique de Prod et TUH ainsi qu’une interaction entre ces facteurs tel que formulé dans le DEM. La figure 2 illustre la forme générale de cette équation inspirée des attendus théoriques de Svensson et al (2012).
Figure 2 : Illustration en 3 dimensions des attendus théoriques de Svensson et al. (2012). TUH : Taux d’utilisation de l’herbe ; Prod : Métrique de productivité (P100GDD ou P15j).
Afin de tester l’importance du contexte pédoclimatique et du paysage (hypothèse 2) le modèle général suivant a été proposé :
13 Div = b + a1 Prod + a2 TUH+ a3 NSGDD + a4 PP + a5 Prod² + a6 TUH² + a7 Prod * TUH
(éq. 2)
avec NSGDD la somme des GDD annuels basé sur les normales saisonnières et PP la proportion de
prairies dans un rayon de 3km autour du site considéré. Ici, nous supposons que les variables explicatives de climat et de paysage ont un effet additif dans le modèle proposé. A partir des équations 1 et 2 découlent 13 autres équations issues de différentes combinaisons des différents termes en veillant à ce que l’effet des variables TUH et Prod ne soit jamais sous leur forme linéaire seule. 3-2-2) Première étape de sélection : la méthode AIC
Nous utilisons la méthode basée sur l'Akaike Information Criterion (AIC) et décrite par Burnham et Anderson (2002) pour effectuer une première sélection des modèles expliquant le mieux les métriques taxonomiques et fonctionnelles. Cette méthode permet de classer les modèles selon le critère d'Akaike permettant un compromis entre le pouvoir explicatif du modèle et le nombre de variables explicatives le composant. Les meilleurs modèles sur la base de l'AIC ne sont donc pas systématiquement ceux comportant le plus grand nombre de variables explicatives. Les modèles étant considérés comme les meilleurs ont les valeurs AIC les plus faibles. Pour chaque indicateur de diversité, une série d'étapes ont été conduites afin d'identifier le meilleur modèle incluant le modèle nul compris. A chaque modèle m testé est associée une valeur AIC. Cette valeur est tout d'abord comparée à la valeur AIC du modèle nul. Le modèle nul a pour équation : y=M. Où y est l'axe des valeurs de la variable à expliquer, M correspond à la moyenne estimée de la variable à expliquer. En calculant le deltaAIC0 = AIC0 – AICm nous n'avons par la suite conservé que les modèles présentant une valeur AIC significativement inférieure au modèle nul (c’est à dire ayant un delta AIC0 positif mais aussi ayant un pouvoir explicatif significativement différent du modèle nul (inspection de la p-value)). S’il s’avérait que le meilleur modèle selon le critère AIC était aussi significativement différent du modèle nul, alors nous nous le conservions pour la suite des étapes de sélection de modèle. Dans le tableau 1, pour chaque variable explicative, seul les deltaAIC0 les plus forts sont notés. Le classement des différents modèles pouvant aussi s’effectuer via le critère Akaike weight (notée AW). Le critère Akaike weight permet d'effectuer un dernier filtrage permettant d'obtenir un groupe de modèle, par variable à expliquer, contenant le meilleur modèle avec une probabilité de 0,95. En effet la valeur AW représente la probabilité que le modèle qu'elle décrit soit le meilleur modèle parmi l'ensemble des modèles testés.
L'analyse AIC informe également sur l'importance relative des différentes variables explicatives (notée Ivar). L'importance d'une variable explicative correspond à la somme des AW des
modèles incluant cette variable parmi les 15 modèles testés. En d’autres termes, plus une variable est présente dans un grand nombre de modèles considérés, plus il est possible de supposer que cette variable est probablement une variable explicative importante. Dans le tableau 1, les colonnes 5 à 8 donnent les importances relatives des différentes variables seulement pour les modèles significativement explicatifs. Les importances relatives sont exprimées en pourcentage de la somme des importances toutes variables confondues afin d’être plus aisément comparables entre elles. 3-2-3) Seconde étape de sélection : choix du meilleur modèle de régression et test des prérequis
Les modèles résultant de la première phase de sélection doivent respecter plusieurs conditions pour être considérés pertinents, à savoir des modèles permettant de prédire des valeurs avec un degré d'incertitude réduit. Pour ce faire, chaque modèle doit respecter trois prérequis : i) la distribution des résidus ne doit pas s'écarter significativement de la loi normale, ii) la variance des résidus doit être homogène et iii) aucun point ne doit influencer fortement à lui seul les prédictions du modèle. i) Pour tester la normalité des résidus du modèle, un test de Shapiro a été effectué sur les valeurs des résidus de chaque modèle. Tous les modèles respectaient ce prérequis. ii) Pour tester l'homogénéité de la variance, l'inspection visuelle d'un graphique représentant la relation entre les valeurs des résidus et les valeurs prédites par le modèle a été utilisée. Des transformations ont été réalisées, lorsque
14 nécessaire, afin d'améliorer l'homogénéité des variances. Cela n'a pas toujours permis d'obtenir un résultat visuellement adéquat. Les modèles dont la transformation des données n’a pas permis d’améliorer significativement l’homogénéité des variances sont indiqués lors de la présentation des résultats. Nous avons donc pris des précautions concernant l'interprétation de ces modèles. iii) Les points influents sont des points qui, lorsqu'ils sont retirés, chacun séparément, modifient de manière disproportionnée les capacités de prédiction du modèle. Ce sont des points isolés qui tirent fortement le modèle vers eux, le modèle prédit minimisant la somme des écarts au carré des points aux valeurs prédites. De tels points ont été retirés afin d’obtenir un modèle plus représentatif de la majorité des points considérés. Nous avons considéré qu'un point était influent lorsque sa valeur calculée sur la base de la distance de Cook était supérieure ou égale à 0.5. Lorsqu’un point en dehors de cette limite a été retiré, ou qu'une transformation a été effectuée afin de rendre les variances des résidus homogènes, les prérequis ont été testés à nouveau jusqu'à obtenir un modèle respectant les prérequis. L'ensemble des modèles respectant les prérequis sont alors disponibles pour passer la dernière étape de sélection: un test permettant d'évaluer la significativité de chaque variable explicative retenue. Nous avons considéré une variable comme significative lorsqu'il y avait moins de 5 % de probabilité que l'effet de cette variable dans le modèle soit lié à l'effet du hasard. Le niveau de significativité évalué est celui de l'effet net de chaque variable du modèle, c'est-à-dire l'effet de cette variable après prise en compte de la variabilité expliquée par les autres variables du modèle. Pour les modèles comptant plus d'une variable, la(les) variable(s) non explicatives étaient retirées une à une en débutant par la variable présentant la p-value la plus élevée. Dans ce cas de figure, les prérequis ont été testés à chaque nouvelle étape de retrait d'une variable explicative.
4) Packages utilisés :
J’ai utilisé le logiciel R et ses packages pour calculer FRic et Q (package « FD »), mNND (package « lefse »), pour appliquer la procédure basée sur les AIC des modèles (package « AICcmodavg »), pour calculer la distance de Cook et la significativité des variables explicatives des modèles (package « stats »).
Résultats :
Les valeurs de TUH montrent que les prairies étudiées se répartissent le long d’un gradient allant de prairies à faible et à fort régime de perturbation, la valeur moyenne de TUH est de 0.43 (min = 0.24, max = 0.73, ET =0.12, figure 3). La productivité P100GDD montre des valeurs indiquant que les
prairies sont réparties sur un gradient de productivité assez important également : P100GDD est en
moyenne de 73g/m²/100GDD (min = 36 , max = 113 , ET =22, figure 3). La productivité P15j est en
moyenne de 123g/m²/15j (min = 55 , max = 211, ET = 42). Les variables de productivité (P100GDD et
P15j) étant très corrélées (corrélation de Pearson, r =0.91, p-value<0.001), seuls les résultats basés sur
l'indicateur P100GDD ont été présentés dans le texte principal.
Analyse AIC
Concernant les variables explicatives supposées affecter le niveau de diversité dans le cadre du DEM, nous observons tout d'abord que l'indicateur du régime des perturbations TUH n'est présent dans aucun modèle supérieur au modèle nul quel que soit l'indicateur de diversité considéré (Tableau 1). De plus, aucun modèle n'explique mieux que le modèle nul les variations de richesse spécifique et d'équitabilité, principaux attendus théorique du DEM. Bien qu'un modèle expliquant les variations de FRic soient significativement différent du modèle nul, il apparait qu'après transformation (afin de respecter les prérequis sur les résidus des modèles), aucune variable explicative ne soit finalement significative pour ce modèle, les variables étant alors toutes marginalement significatives.
Concernant les autres métriques fonctionnelles, nous observons qu'aucun modèle n'apparait meilleur que le modèle nul lorsqu’on se base sur la métrique mNND, que ce soit dans l'espace à 4 dimensions
15 ou pour chaque trait fonctionnel considéré séparément. Cette observation est aussi valable pour l'entropie quadratique de Rao calculée dans l'espace à 4 dimensions. Un ensemble de modèles a cependant été sélectionné lors de l’analyse AIC et les modèles finalement retenu à l’étape de sélection suivante sont présentés ci-dessous.
Modèles sélectionnés
Le tableau 2 résume les différents modèles jugés pertinents. L'étendue des valeurs de masse de graines est négativement corrélée au niveau de productivité P100GDD (modèle n°4 ; adj.R² =0.51 ,
Figure 3), tandis que le niveau de perturbation n'affecte pas la diversité. Nous observons que les étendues des valeurs de SLA sont corrélées négativement avec les NSGDD (modèle n°1 ; adj.R² =0.21
) et les étendues des valeurs de LDMC sont corrélées négativement avec la PP (modèle n°6 ; adj.R² = 0.20).
L'entropie quadratique de Rao apparait expliquée par les variables NSGDD et PP, non considérées dans
le cadre du DEM. QSLA est négativement corrélée à NSGDD (modèle n°2 ; adj.R² = 0.43), Qln(SM) est
corrélée positivement avec NSGDD (modèle n°5 ; adj.R² = 0.28), et enfin QLDMC est corrélée
négativement à PP (modèle n°7 ; adj.R² = 0.19). Pour ce dernier modèle, la variance des résidus augmente clairement avec les valeurs prédites, rendant l'interprétation des résultats délicate.
Variations du CWM
Le CWMSLA est expliqué par le niveau de productivité P100GDD (relation positive) et la durée
de saison de végétation NSGDD (relation négative) (modèle n°3 ; adj.R² = 0.53), mais semble
indépendante du niveau de perturbation (Figure 4). Cependant la variance des résidus augmente avec les valeurs prédites, suggérant que les sites présentant un CWMSLA sont plus difficiles à prédire. Si
aucune métrique associée à la hauteur de canopée n’est apparue expliquée significativement par les variables explicatives retenues, le CWMCH apparait cependant positivement associé de manière
marginalement significativement au niveau de productivité (p-value = 0.07). Discussion
Absence de mise en évidence du DEM et de ses extensions
Le DEM et son extension afin de prédire le niveau d'équitabilité taxonomique n'a pas été mis en évidence dans cette étude. Par ailleurs l'extension du DEM pour la prédiction du niveau de diversité fonctionnelle proposé par Loranger et al. (2016) n’a également pas été largement confirmée. Nous observons uniquement que l'étendue des valeurs prises par la masse de graines d’une communauté prairiale est d'autant plus faible que la productivité est forte, suggérant une convergence fonctionnelle. Ces résultats ont également été observés dans une autre étude soulignant que l'étendue des valeurs de masse des graines diminue avec la profondeur des sols, c’est-à-dire. dans les milieux où les ressources disponibles sont plus importantes (Bernard-Verdier et al., 2012). Cependant, plusieurs mécanismes peuvent conduire à une convergence fonctionnelle, parmi lesquels la dominance compétitive, mais également l’effet d’un filtrage environnemental d’origine abiotique (Perronne et al., 2017). L’étude mise en place permet cependant de confirmer certains attendus d’écologie fonctionnelle. A titre d’exemple, la valeur moyenne de SLA d’une communauté (CWMSLA) apparait positivement corrélée à la productivité des prairies étudiées. Ceci pourrait
s’expliquer par la corrélation connue entre le SLA et le RGR (taux de croissance par unité de masse à l’échelle individuelle, Hunt & Cornelissen, 1997; Poorter et al., 2003). Le CWMSLA est également
16
VAE Équations des modèles AW
S J 1.2 0.34 39 2.1 43 15 3.8 0.67 3 3.4 72 21 0.16 4.6 0.69 2.8 2.6 74 21 0.18 5.7 0.3 42 0.96 32 25 0.19 0.15 0.11 0.086 0.081 0.071 3.7 0.57 63 1 17 19 0.15 2.1 0.5 55 1.5 26 17 4.2 0.74 3.1 3 78 16 2.1 0.5 22 4.8 16 57 2 0.52 9.1 2.9 30 58
Tableau 1 : Sélection des modèles selon le critère AIC. Mesure de productivité : P100DDEG AIC0 - AICmin IP100DDEG (%) ITUH (%) INSDDEG (%) IP P (%) 0.16 (n.s.) 0.40 (n.s.) FRic Div = b + a1 * P100GDD + a2 * NSGDD + a3 * P100GDD² Qmulti -2.6 (n.s.) mNNDmulti -1.4 (n.s.)
RangeSLA Div = b + a1 * NSGDD
Div = b + a1 * NSGDD + a2 * PP QSLA Div = b + a1 * NSGDD Div = b + a1 * NSGDD+ a2 * PP mNNDSLA 1.1 (n.s.) CWMSLA Div = b + a1 * P100GDD + a2 * NSGDD + a3 * P100GDD² Div = b + a1 * P100GDD + a2 * P100GDD² Div = b + a1 * P100GDD + a2 * PP + a3 * P100GDD² Div = b + a1 * PP Div = b + a1 * NSGDD + a2 * PP Div = b + a1 * P100GDD + a2 * NSGDD + a3 * PP + a4 * P100GDD² Div = b + a1 * NSGDD RangeCH 1.2 (n.s.) QCH -0.92 (n.s.) mNNDCH -2.5 (n.s.) CWMCH Div = b + a1 *P100GDD + a2 * P100GDD² Div = b + a1 * P100GDD + a2 * NSGDD + a3 * P100GDD² RangelnSM Div = b + a1 * P100GDD + a2 * P100GDD² QlnSM Div = b + a1 * NSGDD mNNDlnSM -0.087 (n.s.) CWMlnSM -2.8 (n.s.) RangeLDMC Div = b + a1 * PP QLDMC Div = b + a1 * PP mNNDLDMC 1.1 (n.s.) CWMLDMC 0.71 (n.s.)
AIC0 - AICmin : deltaAIC0 du modèle ayant la valeur AIC la plus basse. (n.s.) : modèle non significativement plus explicatif que le modèle nul au seuil de 5% ; AW : Akaike Weights ;
VAE : Variables À Expliquer ; IVar : Importance de la variable Var dans l'explication de la variable à expliquer. Où Var = P100GDD, TUH, NSGDD ou PP.
17 Figure 3 : Variation de la biomasse à l’intérieur et à l'extérieur de l'exclos pour les 18 sites. Bint = biomasse récoltée à l'intérieur de l'exclos ; ext = biomasse récoltée à l'extérieur de l'exclos. Les abréviations correspondent aux différents sites échantillonnés
400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours ARD Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours BEG Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours ChMAR Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours DON Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours DOU Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours ESC Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours FAV Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours HER Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours ISL Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours ISS Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours JUN Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours LAM Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours LIM Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours MAR Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours MOL Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours OYE Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours SER Bint Bext 400 800 1200 0 200 400 600 800 bi om as se (g /m ²)
somme degrés jours YGR
Bint Bext
18 Figure 4 : Illustration des modèles issus de
la seconde étape de sélection. TUH : Taux d’utilisation de l’herbe ; 3 Classes de TUH quand n = 18 :
+ : (0.24 ; 0.40] , n= 11 ; ++ : (0.40 ; 0.57], n= 4 ; +++ : (0.57;0.73], n = 3)
18 Tableau 2 : Modèles issus de la seconde étape de sélection. Mesure de productivité : P100GDD
Influence du climat et du contexte paysager
Le climat et le contexte paysager expliquent – bien que dans des proportions modestes – les variations de certaines métriques de diversité fonctionnelles, comparativement au niveau de
perturbation et de productivité qui constituent des facteurs locaux. Ainsi, l'étendue prise par les valeurs de SLA est d'autant plus faible et sa convergence fonctionnelle d'autant plus forte que la durée de saison de végétation est importante. Ceci signifie qu’en absence de contrainte de croissance liée à la température et à la période potentielle de croissance, les valeurs de SLA entre espèces apparaissent plus similaires. Nous savons par ailleurs que la valeur moyenne de SLA tend à être plus faible lorsque la durée de saison de végétation est plus élevée. Si l'on considère que le SLA traduit de manière prédominante des variations de RGR, alors nous pouvons dire que moins les communautés sont contraintes par la durée de végétation, plus les taux de croissances des espèces abondantes sont réduits et associés à des valeurs plus faibles, ce qui est cohérent avec la stratégie classique des espèces compétitrices, à croissance plus lente mais accumulation plus importante de biomasse afin notamment d’atteindre des hauteurs plus importantes (Sun et Frelich, 2011). Certains traits, tels que la masse des graines, semblent présenter un résultat opposé au SLA. Nous mettons en effet en évidence une divergence fonctionnelle le long du gradient d’augmentation de la durée de saison de végétation. Ces relations contraires sont fréquemment observées entre traits végétatifs (ex. le SLA) et traits de régénération (Swenson et Enquist, 2009). Enfin, concernant le LDMC, il apparait qu’une
augmentation de la proportion de prairies dans le paysage avoisinant augmente la convergence fonctionnelle pour ce trait, suggérant un filtrage environnemental, bien qu’il soit ici difficile d’en proposer une explication.
Ces résultats tendent à supporter l’idée que le contexte pédo-climatique et - dans une moindre mesure - paysager sont les principaux déterminants de la structure des communautés prairiales auvergnates, indépendamment de la productivité des sites et du régime d’utilisation de l’herbe. Une part importante des variations de diversité reste non expliquée. Ceci suggèrent également l’importance forte de facteurs non pris en compte ici comme l’histoire des sites ou l’importance potentielle de mécanismes neutres de structuration des communautés prairiales (Hubbell, 2001). Il faut cependant considérer certaines assertions du DEM potentiellement peu réalistes et les limites méthodologiques de notre étude.
La temporalité des effets de la productivité et du régime de perturbation sur la diversité
La stabilité du régime des perturbations et de la productivité d’une année sur l’autre est une hypothèse forte du DEM (Huston 1979). Dans notre cas, nous avons supposé que les prairies étaient
n° Équations des modèles b a1 a2
1 4.6 (***) 0.21 2 6.9 (***) 0.43 3 33 (***) 0.53 4 6.7 (***) 0.51 5 0.28 6 201 (***) -80 (*) 0.20 7 3296 (***) -2006 (*) 0.19 adj. R² ln (RangeSLA) = a1 * NSGDD + b -5.5 * 10-4(*) ln (QSLA) = a1 * NSGDD+ b -1.3 * 10-3 (**) CWMSLA = a1 * P100GDD + a2 * NSGDD + b 1.2 * 10-1 (**) -5.5 * 10-3 (*) Rangeln (SM) = a1* P100GDD + b -3.1 * 10-2 (***) Qln (SM) = a1 * NSGDD + b 5.6 * 10-2 (n.s.) 8.5 * 10-4 (*) RangeLDMC = a1 * PP + b QLDMC = a1 * PP + b
n° : numéro identifiant ; adj.R² : R² ajusté ; PP : proportion de prairies environnantes ;
NSGDD : somme des degrés jours de croissance ; Codes correspondants aux niveaux des p.value : *** : 0<P < 0.001 ; ** : 0.001<P < 0.01 ; * :0.01<P < 0.05 ; n.s. : 0.05<P<1
19 gérées de la même manière depuis des années. Cependant, nous savons pour quelques sites que les conditions météorologiques exceptionnelles de ce printemps 2017 ont poussé certains agriculteurs à adapter leurs pratiques et que malgré une certaine stabilité des modes de gestion en prairie permanente les exploitants sont parfois amenés - comme cette année - à les moduler. De même, la productivité des prairies varie fortement sous l’effet des variations climatiques interannuelles. Or, il a été montré que les modifications des régimes de gestions mettent plusieurs années à produire des effets et que la production et la diversité des prairies montrent des variations interannuelles fortes malgré un régime de gestion strictement constant (Louault et al., 2017). Ceci suggère que les variations environnementales interannuelles s’ajoutent aux effets moyens des niveaux de production et de perturbation au point probable de les confondre.
Richesse relative et richesse absolue.
Les prédictions du DEM sont des prédictions de richesse spécifique relative au pool local d’espèces et le fait d'utiliser la richesse spécifique absolue dans nos modèles constitue une raison potentielle du fait que nous n'observions pas les attendus théoriques du DEM (Huston, 2014). En effet, supposons que le DEM soit pertinent pour notre cas d’étude et prenons l'exemple de deux prairies ayant des pools locaux de différentes tailles. Une prairie présentant un pool local réduit pourra avoir atteint une valeur de richesse spécifique relativement faible constituant cependant son maximum, tandis qu’une prairie présentant un pool local large pourra être considérée comme plus riche sur la base d’une mesure de richesse absolue, mais sans pour autant avoir atteint son maximum de richesse tenant compte du pool local. L'utilisation de la richesse spécifique absolue pour étudier l'effet des perturbations et du niveau de productivité sur le niveau de coexistence des espèces dans différents contextes biogéographiques pourrait ainsi induire de fausses conclusions. Il faut noter que les études qui ont pu mettre en évidence une partie des attendus du DEM (ex. Laliberté et al., 2012 ; Louault et al., 2017) étaient des expérimentations mise en place sur un seul et même site d’étude (un pool d’espèces commun). Il apparait ainsi nécessaire dans les études multi-sites de caractériser explicitement le pool d’espèces (Butaye et al.,2001 ; Lessard et al 2011).
Limitations méthodologiques à la mise en évidence des diverses forces modulant le niveau de diversité
Parmi les choix expérimentaux réalisés, le faible nombre de sites étudiés a pu être un frein à la mise en évidence du DEM et de ses extensions, compte tenu notamment du grand nombre de facteurs considérés.
La pertinence des métriques employées pour tester les hypothèses et des mesures effectuées sont aussi des points ayant potentiellement freiné la mise en évidence du DE :
- Le taux d'utilisation de l'herbe n’a eu aucun pouvoir explicatif dans les modèles sélectionnés. Le calcul du TUH est une estimation du taux d'utilisation de l'herbe, ne prenant pas compte le phénomène de surcompensation lors de la repousse dans les zones fauchées ou pâturées. Par ailleurs, nous savons par que la nature de la perturbation (pâturage bovin, ovin ou équin et fauche) et pas seulement sa sévérité interagit avec la productivité pour piloter la diversité spécifique (Socher et al., 2013).
- La durée de saison de végétation dans notre étude, à l’instar de Chalmandrier et al. (2017) prend en compte les degrés jours, sans intégrer les précipitations. Cependant, des contraintes de sécheresse potentielle contraignant la croissance des plantes auraient pu être intégrées à cet indicateur, comme proposé par Loranger et al. (2016) afin de rendre compte du nombre de jours favorable à la pousse des plantes (c'est-à-dire pour des conditions de température et de pluviométrie donnée).
- Les hypothèses que nous avons faites ne supposaient pas par exemple d'interactions entre les variables locales (perturbation et productivité) et régionale (climat et paysage). Si une telle interaction existe, il est probable qu'elle ait été responsable de notre incapacité à capter les variations attendues en fonction des prédictions du DEM. Pour des raisons de puissance statistique associée à la faible taille de notre échantillon de prairies, nous n’avons pu tester cette interaction.