CD4+CD25−mTGFβ+ T cells induced by nasal application of ovalbumin transfer tolerance in a therapeutic model of asthma
11
0
0
Texte intégral
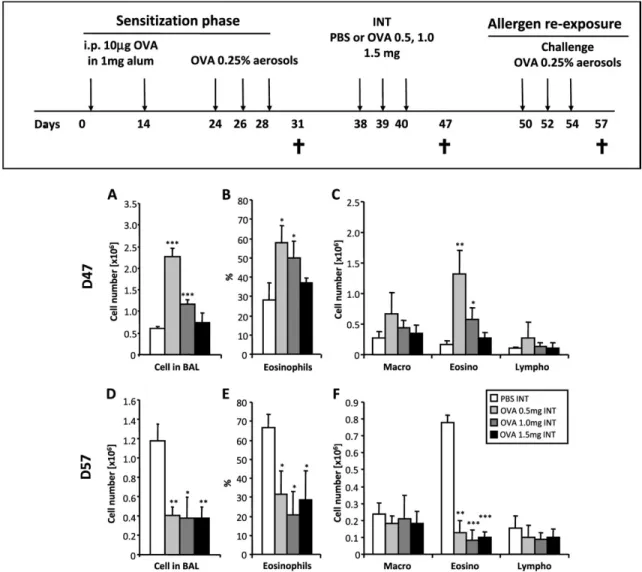
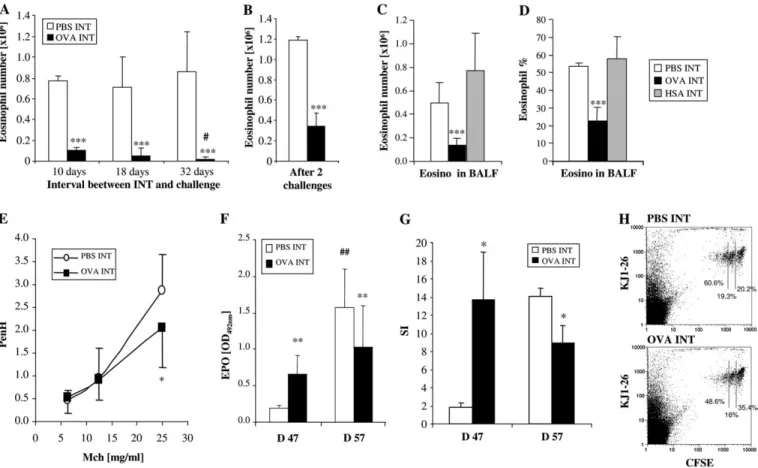
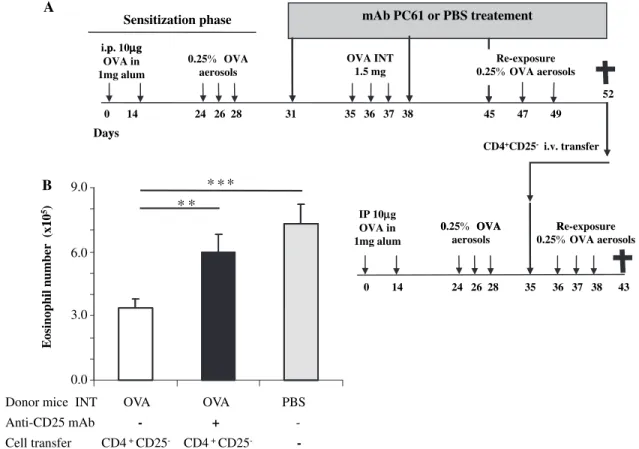
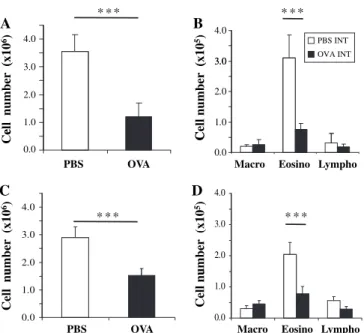
Figure


+3
Documents relatifs