Identification of reference genes for normalizing RNA expression in potato roots infected with cyst nematodes
Texte intégral
Figure
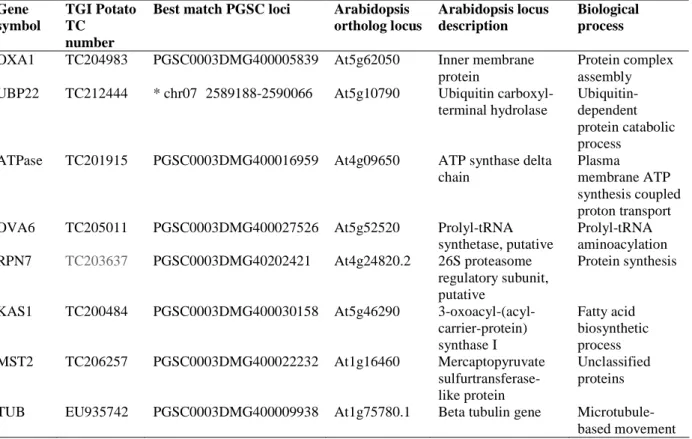

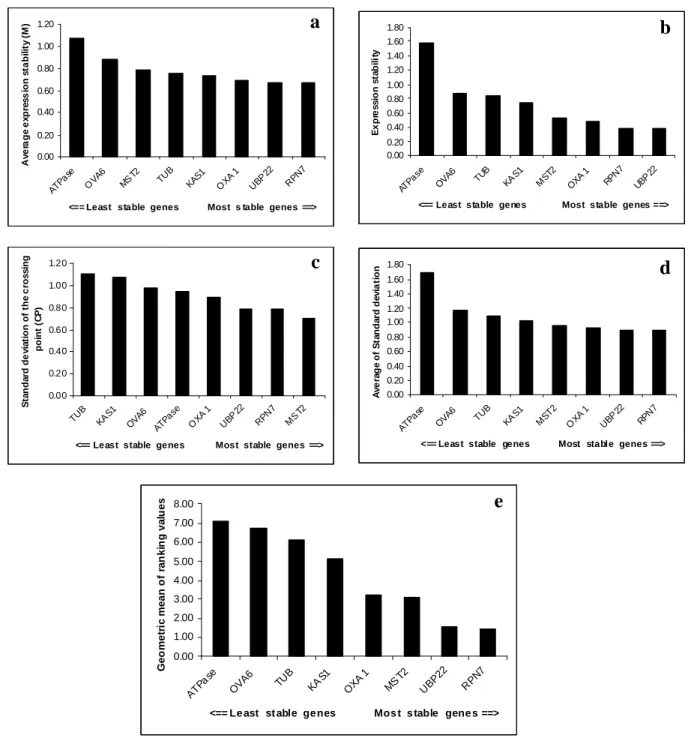
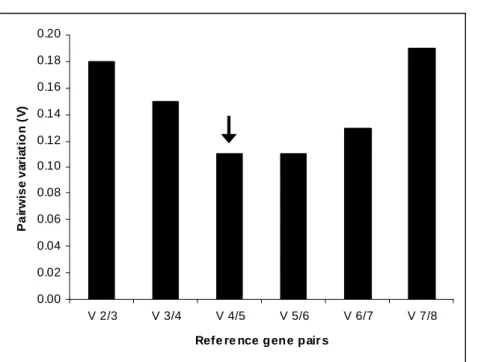
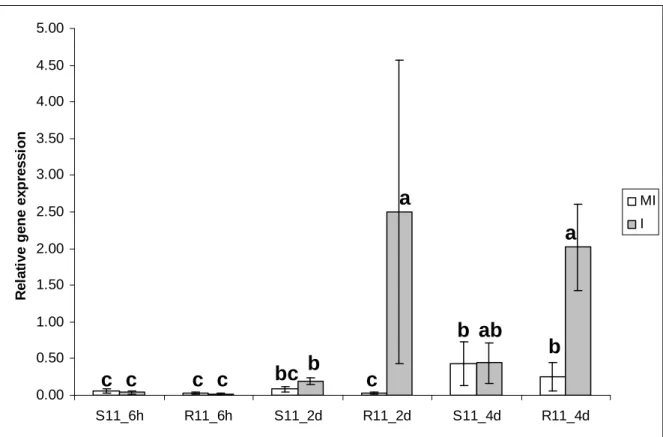
Documents relatifs
ficial effects of air pollution. Most of the 30 models available in that literature showed severe data and statistical problems. It seems apparent that the health
Abstract: The purpose of this study was to develop a method for identifying useful patterns in gene expression time-series data. We have developed a novel data mining approach that
Seven alternative methods for the analysis of gene expression data were compared for their empirical false discovery rates of di fferentially expressed genes.. The
Depuis la réapparition du tramway dans l’agglomération parisienne (ligne T1 entre Saint-Denis et Bobigny en 1992), s’est tissé un réseau incomplet de neuf lignes (105
The central problem of efficiently indexing a BOF im- age representation is that, to our knowledge, there exists no efficient approximate nearest neighbor search algorithm for
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
Le chapitre 4 portera sur une évaluation des performances des contrôleurs ONOS et ODL sur la découverte de la topologie dans un environnement avec un contrôleur unique puis dans le
Trouve un nom générique pour regrouper les noms particuliers de chaque liste... la gourmandise - la colère - le mensonge -
