Extracting representations of cognition across neuroimaging studies improves brain decoding
Texte intégral
Figure


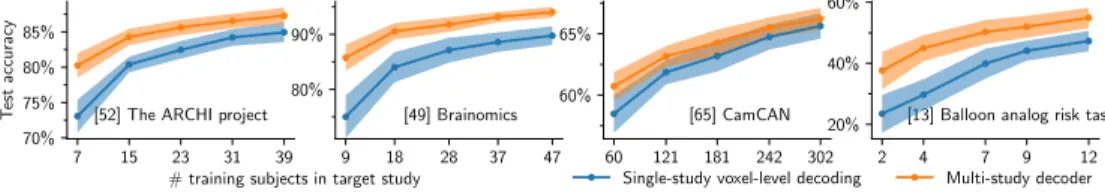



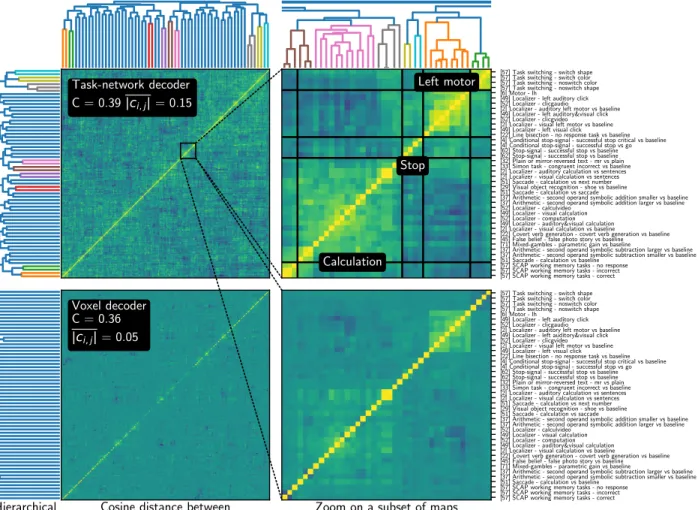
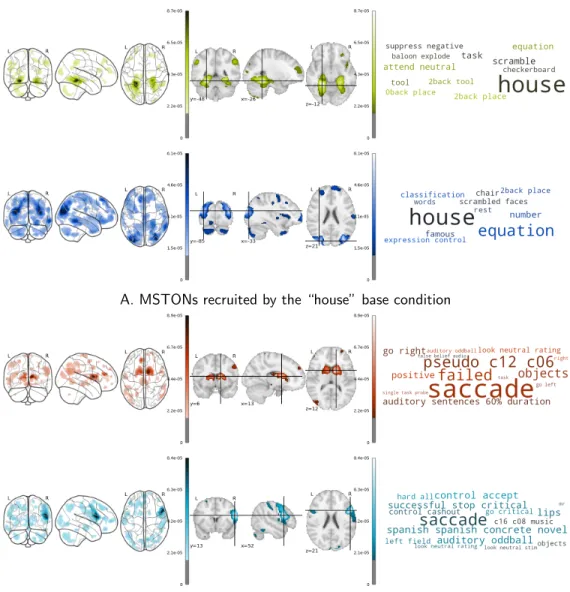

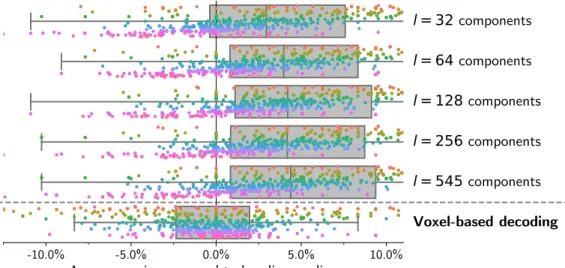
Documents relatifs
«Le mal dont nous souffrons tous c'est de vivre dans un monde qui n 'est ni à notre échelle, ni à notre image, où tout se fabrique à la chaîne.. Il est urgent
Cette étude que nous avons entreprise dans le cadre de cette thèse a pour objectif principal l’optimisation d’une part des paramètres inhérents aux deux méthodes de
ﻒﺳﻮﻳ ﺪﻟﻭ ﻰﻔﻄﺼﻣ. ﺔﻴﻌﻤﺎﺠﻟﺍ ﺔﻨﺴﻟﺍ : 2012 / 2013 ﰲ ﻱﺩﺮﺴﻟﺍ ﺀﺎﻨﺒﻟﺍ ﺔﻳﺭﺎﻤﻌﻣ ﺮﺼﻣ ﺓﺎﺘﻓ ﺔﻳﺍﻭﺭ ﺔﻴﺋﺎﻴﻤﻴﺳ ﺔﺑﺭﺎﻘﻣ ﻑﻭﺮﺻ ﺏﻮﻘﻌﻴﻟ.. ﺮـﻜﺷ ﺔـﻤﻠﻛ ﻡﺪﻘﺘﻧ ﻥﺃ ﻻﺇ ﻊﺿﺍﻮﺘﳌﺍ ﺚﺤﺒﻟﺍ ﺍﺬﻫ ﺎﻨﻴﺃ ﻥﺃ
In any case, the Zn-binding mode of hS100A8 is very similar to that of the four other S100 proteins for which crystal structures in the presence of zinc have been reported,
The intersystem crossing dynamics are affected by the excitation wavelengths, and a follow-up TR-PES study on 2-TU has shown a systematic decrease of time constants with
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
The lines of projection are parallel and intersect the view plane at an oblique angle, thereby providing a view of object depth.. The face of the object that is parallel
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB.
