ur{rvnRsrrÉ
DU
euÉnnc
À
mvrousKr
Rôle du
régime
deperturbations
sur la biodiversité
et lefonctionnement
desécosvstèmes
marins
Thèse présentée
dans le cadre du programme de doctorat en océanographie
en vue de
l'obtention
du grade de docteur ès sciencesPAR
O
Annie
SéguinComposition
dujury
:Emilien
Pelletier président
dujury,
Université
du Québec àRimouski
Phitippe
Archambault, directeur
de recherche,Université
du Québec àRimouski
Dominique Gravel, codirecteur
de recherche,Université
du Québec àRimouski
PauI Snelgrove,examinateur
externe,Memorial University
ofNewfoundland
LINIVERSITÉ DU
QUÉBECÀ
TuOUSKI
Service de la bibliothèque
Avertissement
La diffusion
de ce mémoire ou de cette thèse sefait
dans le respect des droits de sonauteur,
qui
a signé le formulaire <<Autorisation
de reproduire et dedffiser
unrapport,
unmémoire
ou
une
thèse >>.En
signant
ce
formulaire,
I'auteur
concèdeà
l'Université
du Québec à Rimouski une licencenon
exclusived'utilisation
et de publication de la totalitéou
d'une
partie importante de sontravail
de recherchepour
desfins
pédagogiques et noncommerciales. Plus précisément,
l'auteur
autoriseI'Université
du
Québecà
Rimouski
àreproduire, diffuser, prêter, distribuer ou vendre des copies de son travail de recherche à des
fins
non commerciales sur quelque support que ce soit,y
comprisl'Internet.
Cette licenceet
cette autorisation n'entraînent pas une renonciationde
la
part de l'auteur à
ses droitsmoraux
ni
à ses droits de propriété intellectuelle. Sauf entente contraire, I'auteur conserve la liberté de difftrser et de commercialiser ou non ce travail dontil
possède un exemplaire.<<
Je
crois
quel'humanité
est
dans le raison par la science. >>18e3)
1'avenir
de progrèsde
laREMERCIEMENTS
Que de chemin parcouru!
J'aifaitmon
entrée dans le labo dePhil àl'été2006
àtitre
de stagiaire d'été. Et oui,I
ans
de
collaboration entretoi
et
moi, Phil. De tout
ce
quetu m'as
apporté,c'est
de taconfiance dont
je
suis la plus reconnaissante. Dèsle
départ, encore étudiante au bacc,j'ai
été
en
charge de mon propreprojet,
qui
incluait
unesortie
enmer,
des incubationset
la maintenance d'espèces en bassin.Bon,
inutile
de rappeler quej'ai
échoué sur ce dernieraspect...
Plus
de
600
échinodermesont
trouvé mon
inexpérience disons
difficile
àsupporter! Tu m'as aussi donné carte blanche pour recruter mes stagiaires d'été
0'ai
choisi les meilleuresd'ailleurs, hein
Pauline et Marie-France!).Merci
pour
l'autonomie quej'ai
pu
développer,tu
as toujourscru
en mon potentiel. Grâce àtoi,
j'ai
maintenant de solides connaissances en plomberie et en structure de béton.Merci
dem'avoir
transmis ton intérêtpour
les stats et dem'avoir
offert
autant de latitude commeauxiliaire
d'enseignement. Jeretiendrai aussi toutes nos conversations, nos débats, dont un particulièrement mémorable, quelque
part
sur une route dela
Toscane. Quedire du Brunello
offen
pourmon
mariage,qui
vieillit
sagement dans mon sous-sol. Sois sûr quej'aurai
une pensée pourtoi
lorsqueje
I'ouwirai
dans quelques années!Dominique, tu as accepté de te
joindre
àmoi
àtitre
de codirecteur, rôle quetu
as suassumer à la perfection.
Tu
as été un mentorpour moi. Ton
esprit d'analyse etla
vitesse grandV
à laquelle fonctionnent tes neuronesm'auront
donnédu
fil
à retorde lors de nos discussions, maism'auront
assurément permis de repousser meslimites
et dem'ouwir
àd'autres facettes de
l'écologie.
Je suis reconnaissante de ta grande générosité, de ton temps et de ton écoute. Merci pour le coaching de rédaction. Ce sont des aptitudes qui me suivront toute ma vie.Je vous remercie tous les deux pour votre ouverture face à mes projets et obligations
familiales.
Jamaisje
ne me
suis
sentiejugée. Votre
support
a
eu
des
répercussions importantes et positives sur mon travail, mon quotidien et les personnes qui m'entourent.Merci
àÉmilien
Pelletierd'avoir
fait
parti de monjury
àtitre
de membre interne.Tu
avais
le
recul nécessaire pour me questionner et me suggérer des idées nouvelles, toujourspertinentes.
Tu
as aussi été très généreux de bienvouloir
partager les locaux dela
stationpour
la
réalisation
d'un
de
mes projets
et
même
de m'avoir
présentéeà
l'équipe
deDécouverte.
Paul, thank you
for
having accepted to be part of thejury
as external member.I
havea
great respectfor
your
work
andyour contribution
to
the
oceanography. Thankyou
for
being sucha
great leaderwithin
CHONe.Je veux évidemment remercier
le
Conseil de recherches en sciences naturelles et engénie (CRSNG) et le Fonds de recherche du Québec
-
Nature et technologies (FRQNT). Ils ontfinancélaquasi-totalité
de mes études, du bacc à ma dernière année de doc. Je remercieeuébec
Océanqui m'a
permis
de passer5
mois
àPise en
ltalie
dansle
laboratoire duprofesseur Lisandro
Benedetti-Cecchi.
J'ai
aussi
pu
bénéficier
d'une
expérienceenrichissante
en
siégeant
à
leur
conseil
d'administration.
Merci
à
CHONe
pour
le dynamisme du réseau, qui a encouragé les échanges entre étudiants et chercheurs de partout au Canada.Grazie
mille à
Lisandro
Benedetti-Cecchiet
particulièrementà
Elena
Maggi
quim'ont
accueillie à I'université de Pise, quiont
partagé avecmoi
leurs techniques de travailet
leur vision de
Ia
science.J'ai eu
la
chanced'échantillonner
en
apnéesur
un
site paradisiaque avec une super équipe. Un merciparticulier
à Rosarrrra, ma mamanitalienne.Grande bacione
ate
caraRosanna!A
presto! Je tiens à remercier Éric Harvey, pour la superéquipe que nous avons
formé.
Je collaborerais avectoi
à nouveau àtout
moment.Merci
aussià
Isabelle Gounandet
Timothée Poisotpour votre
importantecontribution à mon
travail
etpourm'avoir
aidé à apprivoiser (etje
pourrais même dire aimer!) lamodélisation.Xi
Merci
à Pauline Robert,pow
tontravail
sur le terrain, mais avant tout, pour être mameilleure amie depuis maintenant près de 6 ans.
Tu
es présente à toutes les grandes étapes de ma vie avec la même générosité. Marie-France,j'ai
eu énormément deplaisir
à travailleravec
toi.
Je garde en mémoire nos captures de crabes dansle
barachois deCarleton!
Tum'as
épatéepar ton
efficacité sur
le
terrain.
Merci à
Val,
pour
tes diners paninis,
tesdesserts loufoques et ta bonne humeur.
Tu
ajoutais dusoleil
àI'ISMER. Ton
amitié m'estprécieuse. Je
veux
remercier aussiAdeline
et
Virginie,
pour
nos discussionsde
bureau, pour votre soutien et nosmille
fous rires.Vous
allez me manquer. Sois prudenteVirginie
àI'autre bout
du monde!Cindy, merci
àtoi
dem'avoir fait
une petite place dans tavie
etdans
ta
maison.J'admirerai toujours
ton
senscritique
sanspareil.
Laure,
Lisa,
Heike,Mariève, Katrine, Jesi,
j'ai
eu tant deplaisir
à vous côtoyer, etj'ai
appris beaucoup de votreprofessionnalisme.
Merci
àMel,
Mylène et
Rénald, mes amisqui
étiezlà
àmon
arrivée dans le labo. Vous m'avez servi de modèles, de références. La barre étaithaute.Mel,
sacheque
je
veillerai
à ne pas être <trop
gentille >> dans lefutur! Merci
à Vanessa Pencalet pourle coup de main avec mes analyses de laboratoire.
Merci
papad'avoir
partagé avecmoi
ton
amour pourla
nature. Une part importante de mon intérêt pour l'écologie benthiquevient
de ces journées à attraper les écrevisses dansle
creek.Merci
maman,ma fan
numéroun, pour qui tout ce
queje
fais
estplus
quefantastique . Ça
fait
longtemps quej'ai
appris à être plus critique de mes accomplissementsque ce
quetu m'en
dis, mais
sachesque c'est
rassurantde
te
savoir
derrièremoi
peuimporte
cequi
peut arriver. Jete
dois ma passionpour l'eau. Le
sprintfinal
de rédactionn'avraitjamais
été possible sansta
générosité et tes fidèles services de gardiennage.Claude,
tu
asLA
sourced'inspiration. Je
sais queta
curiositépour
les
questionsscientifiques est supérieure à la mienne, et sans cette satanée maladie, tu serais à ma place,
le
doc
en pocheet
une
super carrière devanttoi.
Tu m'as
apprisla
persévéranceet
le courage. Tu m'as permis devoir
certaines embuches avec de la perspective, de comprendrexll
désagréable, mais ce n'est pas une raison valable pour perdre
le
sourire. Je suis choyée det'avoir
à mes côtés à Québec.Merci à mon mari, mon
stagiaire bénévole préféré, Francis.Toi, qui
aspris
desvacances pour
m'aider
sur le terrain, as compilé des données aux petites heures de la nuit,sans compter tous les textes que
je
t'ai fait
relire. Tu as toujours été 1à pour m'épauler et mesoutenir dans
ce
long
périple.
Merci d'avoir
rendu ma
vie
aussibelle
avecnos
<< deuxgarçons > et de nous avoir bâti un nid réconfortant, dans lequel
je
pouvais me réfugier dansles passes
plus
difficiles.
Jet'aime
etje
ne te remercieraijamais
assezpour
les sacrifices que tu as faits, de même que pour toutes ces sautes d'humeur que tu as dû endurer.Louis-Félix
et
Clément,
dur
de
croire que
je
veux
vous
remercier
pour I'aboutissementde mon
doctorat.Vous
avezplutôt
contribuéà
me
couperune
quantitéinnombrable d'heures de sommeil (Clément, on
fait
sesnuits
S.V.P.!)
et à m'empêcher detravailler
avecvos
rhumeset
autrespetits
microbes. Pourtant,merci pour
votre
amourinconditionnel
qui
me
donne envie demordre
àla vie
et d'entrevoir mon
avenir sur une note si positive.AVANT-PROPOS
Je me
suis
intéresséeà
l'écologie
des perturbations
en
constatantà
quel
point I'homme applique une pression énorme sur les écosystèmes marins. Les stocks de plusieurs espèces de poissons sont en chutelibre,
les eaux s'eutrophisentet s'acidifient,
l'oxygène diminue, sans parler dela taille
démesurée de ces(
continents de plastique >>. On ne peutassurément
pas
s'opposer
et
empêcher
le
développement économique
impactantdirectement
et
indirectement
le
milieu marin. Je me suis donc
questionnéeà
savoir comment on pouvait aumieux s'outiller
pour détecter et prédire efficacement les effets deces perturbations sur
la
biodiversité marine etle
fonctionnement des écosystèmesqui
lui
sont associés, afrn delimiter
les impacts négatifs qui peuvent en découler. Cette thèse s'estprincipalement basée
sur
la
comparaison d'approches.De
cette façon, j'espérais pouvoir proposerune ou
des approchesà privilégier, et ainsi
contribuer
à faire
progresser nos connaissances de la gestion et de la conservation des ressources marines.Cette thèse est
intitulée
<Rôle du
régime de perfurbations surla
biodiversitéet
lefonctionnement des écosystèmes marins >.
Elle fait
partie deI'axe
de recherche Effectsof
biodiversity
changesdue
to
anthropogenicactivities
on intertidal
ecosystem services ausein
du
réseau stratégique canadien CanadianHealthy
OceansNetwork (CHONe).
Lesprojets ont été financés par CHONe et le Conseil de recherches en sciences naturelles et en
génie (CRSNG).
Ils
s'insèrent
également dansle
programme
de
recherchedu
groupexiv
LTSTE DES PUBLICATIONS REALISEES DANS LE CADRE DE LA THESE
Séguin
A,
GravelD,
ArchambaultP
(2014)Effect of
disturbance regimeon
alpha andbeta diversity of rock pools.Diversity 6,
l-17
(chapitre 1)Séguin
A,
Maggi
E,
Benedetti-CecchiL,
GravelD,
Archambault,P
(A
soumettre)Role
of
disfurbance frequencyand intensity
on
the beta
diversity
of
benthic
rock
pool
communities. Sera soumis à Marine Ecology Progress Series (chapitre 2)Séguin,
A,
Harvey
E,
Archambault
P, NozaisC,
GravelD
(2014)
Body
size as apredictor
of
speciesloss effect
on
ecosystemfunctioning. Scientific
Reports,
4:4616(chapitre 3)
Harvey
E,
SéguinA,
Nozais
C,
Archambault
P,
Gravel
D
(20t3) Identity
effectsdominate
the
impactsof
multiple
species extinctions on thefunctioning
of
complex foodwebs. Ecology 94, 169-179 (annexe)
J'aitravaillé,
durant mon doctorat, à l'élaboration de l'étudeIdentity
fficts
dominate the impacts of multiple species extinctions on thefunctioning
of complexfood
webs , publiédans la revue scientifique Ecolog,,. Je n'ai pu inclure cet article entarrt que chapitre au sein de la thèse, n'étant pas la première auteure.
J'ai
contribué à sa réalisation à hauteur de40
à50 Yo, en
m'impliquant
autant au niveau de la planification, de la réalisation, des analyses et dela
rédaction. Cette étude est enlien
direct avec la problématique abordée dans ma thèse. C'est une éfude expérimentale réalisée en parallèle àla
fois
dansun
systèmemarin
et unsystème lacustre.
Elle
suggèreque
I'identité
fonctionnelle
seraitplus
importanteque
la richesse spécifiquepour
expliquer les variations
observées dansle
fonctionnement desécosystèmes.
Il
s'agissait dela
première étude expérimentale à testerI'effet
deI'identité
fonctionnelle au sein de systèmes multitrophiques.J'ai
donc décidé del'inclure
à la thèse, enf
insérant en annexe.LISTE DES PRESENTATIONS REALISEES DANS LE CADRE DE LA THESE
Séguin
A,
GounandI,
PoisotT,
GravelD,
Archambault P (2013) Impact de la perted'espèces connectrices des réseaux pélagiques
et
benthiquessur
le
fonctionnement desécosystèmes
marins.
Assemblée générale annuellede
Québec-Océan, Rivière-du-Loup,Canada 13-15 novembre 2013 (affrche;
prix
dela
meilleure affiche)
Séguin
A,
Gravel D, Archambault P (2012)Effet
de la fréquence de perturbations surla
variabilité
spatiale. Assemblée générale annuelle de Québec-Océan,Montréal,
Canada,7 -9 novembre (affiche ;
prix
dela
meilleure affiche)
Séguin
A,
Harvey É, Archambault P, Nozais C, GravelD
(2012) Use of body size asbiological
indicator of species loss effect. Assemblée annuelle de CHONe, Ottawa, Canada, 24-27 octobre (affiche)Séguin
A,
HarveyÉ,
Archambault P, GravelD
(2011).Effet
des extinctions sur lefonctionnement
d'un
systèmemultitrophique.
Assemblée générale annuellede
Québec-Océan, Lac-Delage, Canada, 17-18 novembre (affiche)Séguin
A,
Harvey É, Archambault P, GravelD
(2011)Body
size as impact predictorof
biodiversity
loss on ecosystemfunctioning.
World
Conference on Marine Biodiversity,Aberdeen, Écosse, Royaume-Uni, 26-30 septembre (oral)
Séguin
A,
HarveyÉ,
Archambault P, GravelD
(2011) Speciesidentity
and richnessefficiently
predict the impact
of
biodiversity
losson
ecosystemfunctioning
in
complex marinefood
webs. Assemblée annuelle de CHONe,Montréal,
Canada, 28-30awil
(oral;mention)
Séguin
A,
Harvey É, Archambaut P, GravelD
(2010) Et si être grosétaitbonpour
lasanté des écosystèmes ? Émde de
I'effet
detaille
surle
fonctionnement des écosystèmesmarins. Assemblée annuelle
de
Québec-Océan, Lac-Beauport, Canada,ll-12
novembre(affiche)
Séguin
A,
Archambault P (2010) Plus ça change plus c'estpareil:Diminution
de lavariabilité
spatialed'un
assemblage des suitesd'une
perfurbation chronique. Assembléeannuelle de
la
Société Canadienned'Écologie et d'Évolution,
Québec, Canada,9-I2
marxvl
Séguin
A,
Archambault
P
(2009) The
effect
of
disturbance frequencyon
spatialvariability
of
benthic community
as
a
managementtool.
European
Marine
Biology
Symposium, Liverpool, Royaume-Uni, 7
-ll
septembre (oral)Séguin
A,
Archambault P (2009) La fréquence favorise la perfoffinance. Colloque devulgarisation
scientifiqueLa
Biologie
danstous
cesétats,
Rimouski,
Canada,13
mars(oral)
Séguin
A,
Archambault P (2008)Did
disturbances increasespatialvaiability?
2008Assemblée
annuelle
de
Québec-Océan, Rivière-du-Loup,
Canada,
27 --28 novembre (affiche)Séguin Ao Archambault
P
(2008)In
situ
determinationof
the impact
of
press andpulse
disturbanceson
spatial
variability
of
a
St.
Lawrencerocky intertidal
ecosystem assemblage. Conference annuelledu
Conseil hrternationalpour
l'Exploration
dela
Mer,Halifax,
Canada, 22-26 septembre (affiche)nnsumn
Les
perturbationsnaturelles
et
anthropiquesjouent
un rôle
déterminant dans la structure des communautés naturelles. Sousl'effet
des changements climatiques, le régime de ces perfurbations est appelé à semodifier.
Les perturbations seront ainsi plus fréquenteset
de plus forte
intensité.
Il
est
essentield'être
en
mesurede
détecteret
de
prédire efficacementI'effet
des changements actuels et anticipés des régimes de perturbations sur la biodiversité, alors que le tauxd'extinction
des espèces serait présentement de 100 à 1000fois plus
élevé que lestaux
estimés dansle
registrefossile.
L'objectif
principal
de cettethèse était donc
d'estimer
expérimentalement les effets du régime de perturbations sur labiodiversité
et
le
fonctionnement des écosystèmesmarins.
La
chained'impact
entre le régime des perturbations, la biodiversité et le fonctionnement des écosystèmes a été étudiéeen
deux
temps.D'abord,
nous
avons testéI'effet
combiné (chapitre
1) et
indépendant(chapitre
2)
des régimes de perfurbations (fréquence et intensité) surla
biodiversité. Puis,alors
qu'il a
éIé suggéré quef
identité
fonctionnelle,une
composantede la
biodiversité,dominerait les effets
d'extinctions
sur le fonctionnement des écosystèmes, nous avons testéle potentiel
prédictif
de lataille
des espèces au sein de systèmes multitrophiques (chapitre3).
Nous
avons étudiéle rôle
des perturbations sur des communautés benthiques sessilesqui
colonisent les cuvettes marines en zone intertidale. Ces espèces sont reconnues commeétant
d'excellentes espèces sentinelles,qui
nous aident
à
détecter des changements debiodiversité
au
sein d'autres
systèmes,moins facilement
accessibleset plus
coûteux àéchantillonner.
Les
chapitresI
et
2
ont
été réalisés directement surle
terrain
au sein decuvettes marines naturelles. Le chapitre 3
a
été divisé en deux expérimentations, réalisé enparallèle dans des mésocosmes marins et lacustres.
Notre premier
objectif
étart devérifier
si rme augrnentation de I'hétérogénéité spatialede
la
structured'une
communauté sessile benthique (diversité bêta)était
indicatived'un
environnement perfurbé(chapitre
I
et
2).
L'effet
des perturbationssur
I'hétérogénéité spatiale s'est avéré beaucoup plus complexe. La réponse des communautés serait moduléepar le
régime (fréquenceet
intensité) associé àla
perfurbation. Selonqu'elle
survienne àfaible ou fort régime, une perturbation aurait des effets opposés sur l'hétérogénéité spatiale.
La forme de la relation, à savoir si I'hétérogénéité augmente, diminue, linéairement ou non,
adifféré
en fonction de la source de la perfurbation appliquée. La fréquence et I'intensité de perfurbations agiraient aussi indépendammentI'une de I'autre et
n'affecteraient pas lesmêmes composantes de
la
structure dela
communauté.La
fréquenced'une
perturbationinfluence
la
composition, alors queI'intensité
affecteplutôt
le recouvrement des espèces.xv1il
de la biodiversité autant, sinon plus importante que
la
richesse spécifique (diversité alpha) pour évaluerl'effet
des régimes de perturbations.Dans
le
chapitre3,
nous avons testéle
potentiel de
la taille
des espèces,un
trut
fonctionnel
facile
à mesurer,relié
aux tauxd'extinctions
et à plusieurs taux biologiques, à prédire les effets de la perte d'espècessrr
le
fonctionnement de systèmes multitrophiques. Nous avons montréun
effet
fort
dela taille
des espèces surle
fonctionnement. Cet effetserait toutefois
plus
important lorsque combiné
à
Ia
position trophique,
puisque conjointement, ces deux traits fonctionnels ont expliqué une large proportion de la variationobservée dans
le
fonctionnement des écosystèmes. La
taille fut
un meilleur
prédicteur deI'effet
d'extinctions
surles
fonctions reliéesà
la
consommation.Nos
résultats suggèrent également quel'étude
de séquencesd'extinction
aléatoire sous-estime les effets de la perte d;espèces, comparativement à une séquenced'extinctions
ordonnée en fonction dela
taille,
plus représentative des extinctions enmilieu
naturel.Cette
thèse proposedes
approchesà
privilégier
afin
de
détecteret
prédire
plusefficacement
I'effet
des perturbations. Les résultats obtenus se sont appuyés sur des plansexpérimentaux souvent
plus
réalistesque ce
qui
avait
été réaliséjusqu'à
présent. Nous suggérons, sur la base de nos observations,qu'il
serait plus facile qu'anticipé de prédire leseffets des perturbations
et
dela
perte debiodiversité
associée surle
fonctionnement desécosystèmes.
Mots
clés:
Perturbation,
biodiversité,
fonctionnement
de
l'écosystème, régime, cuvette marine, hétéro gé néité spatiale, extinction, c ommun auté benthiqueABSTRACT
Natural and
anthropogenic
disturbancesplay
a
k.y
role
in
structuring
naturalcommunities.
V/ith
climate change, the regime of these disturbances is expected to change.Disturbances
will
be more frequent and more intense.It
is essential to be ableto
detect andpredict
efficiently
the effectsof
current and anticipated changesof
disturbance regimes onbiodiversity,
while
the rateof
speciesextinction
isnow
100to
1000 times higher than theestimated rates
in
the fossil
record.
The main
objective
of
this
thesis was
to
estimateexperimentally the effects
of
disturbance regime onbiodiversity
and the marine ecosystemfunctioning. The
cascadeof
impacts between
disturbanceregimes,
biodiversity,
and ecosystem functioning was studied in two phases. First, we tested the combined (Chapter 1)and independent (Chapter
2)
effectsof
disturbance regimes (frequency andintensity)
onbiodiversity. Then,
while
suggestedthat
the
functional identity,
a
component
of
biodiversity,
dominates
extinction effects
on
ecosystemfunctioning,
we
tested
thepredictive
capacityof
speciesbody
sizein
multitrophic
systems (Chapter3). We
studiedthe role
of
disturbanceson benthic
sessilecommunities colonizing
rock pools.
These species are often considered as excellent sentinelsfor
estimating changesin
biodiversityin
other
systems, less accessibleand
more
expensiveto
sample. Chapters1
and2
wereconducte
d in
situ
in
marinerock pools
and Chapter3
wascaffied out
in
parallelboth in
marine and freshwater mesocosms.
Our
first
objective wasto
determine whether an increasein
the spatial heterogeneityof a
sessilebenthic community structure (beta diversity)
was
diagnostic
of
disturbedenvironments
(Chapter
I
and
2).
'We
found that the effect
of
disturbanceson
spatial heterogeneity was actually much more complex, since the regime (frequency and intensity)associated to the disturbance modulated the response
of
communities.A
disturbance couldhave opposite effects
on
spatial heterogeneity, dependingon
whetherit
occursat
low
orhigh
regimes.
The
form
of
the
relationship, whether
the
heterogeneityis
increasing,decreasing,
linearly
or not,
dependson the
sourceof
the
disturbance.The
disfurbancefrequency and
intensity
also act independently anddo
not
affectthe
same componentof
community
structure. Disturbance frequency influences
speciescomposition,
whereasintensity
instead
affects
speciescover.
Chapters
1
and
2
also
showed
that
spatialheterogeneity
is
a
biodiversity
measureas
much,
if
not more
important
than
speci esrichness (alpha diversity) in evaluating the effect of disturbance regimes.
In
Chapter3, we
testedthe potential
of
speciesbody
sizeto
predict the
effectsof
species loss on ecosystem functioning. This
functionaltrait
is easilymeasured and linked toXX
on the functioning. However, this effect was stronger when combined
with
trophic position. Together, thesetwo
functional traits explained a large proportionof
thevariation
observedin
functioning.
Species body size was a better predictorof
species loss effects on functionsdirectly
related
to
consumption.
Our
results also
suggestthat the
study
of
randommultispecies sequential
extinctions
underestimatesthe
effects
of
species
loss,
whenrotttpur.d to
non-random sequential extinctions, ordered bybody
size, more representativeof
extinction in natural ecosystems.This
thesis suggested approachesto
detect and predictthe
disturbance effects moreefficiently.
Results were basedon
morerealistic
experimental designs.Our
observations suggestthat
it
may be
easier than expectedto
predict
disturbance effects and associated species loss on ecosystem functioning.Keywords:
Disturbance,biodiversity,
ecosystemfunctioning, regime, rock
pool,TABLE
DESMATIERES
REMERCIEMENTS....oo...o...e ..o...o...o...r...'r..r..r...r..rr...
IX
AVANT-PROPOS ...r..r.r.ro....ro.to.rr.oor....r.r..oo...o..o...r....o...o...r...o...o.. XIIInÉSUVIÉ ...'...o...r.... ...o...oo..o...r....e ..o...o..oo...'... XVII
A[lSTRACT...o....r..r...o..r...o...oot...oo...o...o....o.... XIX
TABLE
DESMATrÈRES...o... ...,.r...
...r...,...XXr
LISTE
DESTABLEAUX
.o...o...o...r..e ...o...r..r..o...o...r...o..."XXV
LISTE
DESFIGURES
...o...o..o...e ...oe ...oo...e ....XXVII
INTRODUCTION
GÉNÉnnI,f...o..r..r...r...o..r...r...r...r.../
8IOUIVERSITE...o...r..o...o..r.r...r. .r..e..r..r...1
PnnfUnnATIONS ...o...o...ooo....e...o....'...eorr...r...2
RnCf nnn DE PERTURBATIONS ...o...o...,..r...o... 6
Dns REGIMES DE PERTURBATIONS EN CHANGEMENT...o....,..e...r...o...or..r.,...8
FoNcTTONNEMENT DES ECosysrEMES...o...,...,.o,.r...o...r...9
Mnrr,r,nuR PREDICTEUR : DIVERSTTE SPECIFIeUE
ou
FONCTIONNELLE?...14OBJECTIF PRINCIPAL
..o..oe ..r...r...r...r...r...o...o ...,.17OnfnCfIFS SPECIFIQUES...o....r...r...o....r...o...or...'...19
Chapitr€ 1...o...o...o..o.e...o...ôr..o.o...o...ô...r...rrr...1-9 Chapitre 2...o...o..ô....r...r...o...r....o.o..r..o...rrr...21 Chapitr€ 3...o...o...o.o.o.o..r...e .r...o....e .r... ...r23
xxii
CHAPITRE
1EFFET
DU nÉcTvTE DE
PERTURBATIONS
SURLA
DIVERS11É
ALpHA ET BETA
DESCUVETTES
MARINES
...o...o...25 RrSunnn aaa..o...o.aa...o....o.aa...r..aaaa...o.aaaaa..o..o...aaa...r...aa...o.o...o..aaaala..o...o...aaaa...25 A$STRACT...oo...o...o...r...o.ôo.o...o...o...o...t...ttttt27INrnonUCTION....e..o...o...o...o....o...r...r...o..ô...o...28 Ma.rnRrALS AND METHODS...r.o...o...o.ô...3L RnSUfrrS...r...o...o..e...r...r...o...r...r...o...35 DrSCUSSION...o...o..r...r...o..r...41
CHAPITRE
2EFFETS
INDEPENDANTS
DE
LA TNÉQUENCE
ET DE
L'INTENSITÉ
DE
PERTURBATION
SURLA
DIVERSITE BETA
DE
CUVETTES
MARINES ...r.o...o...r...o...t.o...o... ...ttttt 49RnSUVIE ...or...o..o...r...r...o..e...o...o... ... 49
AnSfn 4CT...ô...o...o...o...o...o...t.tt 51 M.q.rnRIALS AND METHODS...r...r...r..o...o...o... 54 Rnsu1rS...o...o...or.r.o...o..t....o...t..tt 58 DISCUSSION...ô...oo...o...o...r...e...o...61
CHAPITRE
3LA TAILLE
DES ESPECESCOMME
PREDICTEUR DE
L'EFFET
DE
LA
PERTE
DOESPECES SURLE FONCTIONNEMENT
DE
Lt8COSYSTEME...o...o..ô...toe...t...tttttt 69
RnSunm r.e..aaa...aa...or...aaa...r..o.aaa...o...aaaa...o.aaa...o...ô..o.aaaa...a4aaa...69
Bony
SIZE AS PREDICTOR oF spEcIEs Loss oN ECosYsrEM FUNCTIONING ...71AgSTRACT...r...o...o...o...r..o...o...'ll INrnOuUCTION....rr...o...o...r...e...e...oo...o...o...r...72 MATETALS AND METIIODS...r...o..r...o...o...oo'.ô...73 RnSulrS ..r...o.or...r...r...o...o...o...o..oto...o...ro.t...77
DrScussIoN...r...o...ô...o..e..o...o...o...o.o...o...78
XXiii
CONTnTBUTION DE LtETLJDE...o...o..o...r...,.r.r.rr...o...86 PnnSpnCTMS...r..o...o..o...e ...rr.o...o...r...r..o...92
CONCITUSION FINALE...ô.r.r..r....o...o...ooo.o.r..r...r...o...o...o...o...97
nÉnÉnnNCES
BrBLroGRAPHIeuES...o...rr.o.o..o...,..oe.o,.o...e ...99ANNEXE IDENTITY
EFFECTS
DOMINATE THE IMPACTS
OF
MULTIPLE
SPECIESEXTINCTIONS
ON
THE
FUNCTTONING OF
COMPLEX
FOOD
\ryEBS r...o...e ..'..e ...e ..ô...r....o...o..t...o...o..r... I 13AnSfnACT ...oo.rr..e ...r...o...o...o...o...o.o...o..o... 113
INTROUUCTION ...o..r...o...or...r...o...o....o...o.o...r...,...114
Mnrnmnl',s
AIID METHODS...o..o...o...,...r...o....,...r...o..o...o...L16RBSUITT S ...r.o...r...o..o...o..r...o....o...r..e ...ro...o. ...r....123
LISTE DES TABLEAUX
CHAPITRE
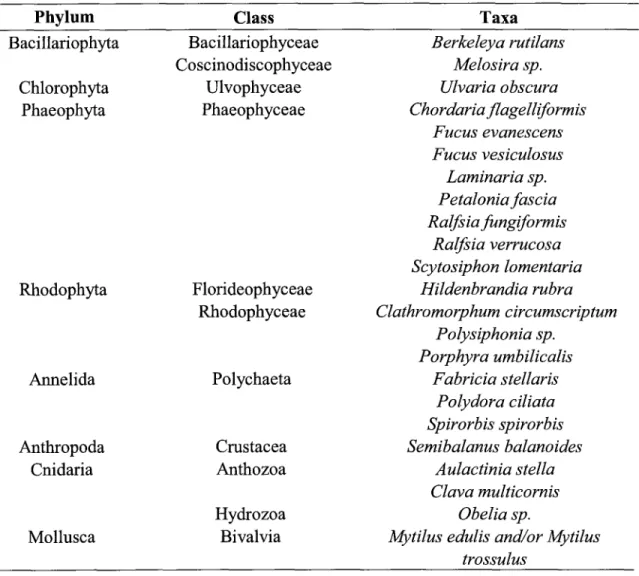
1Table
l:
List
of the 24 taxa identifiedwithin
the rockpools
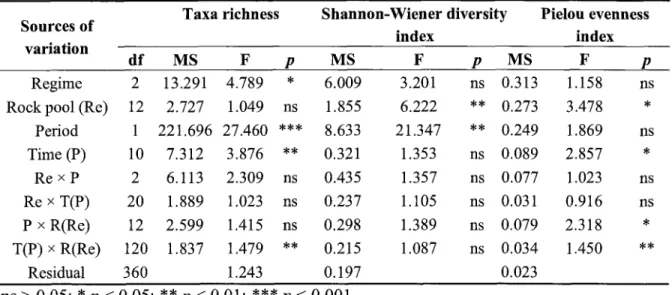
....36 Table 2: Univariate analysis performedwith
afour-way partially
hierarchicalANOVA
for
differencesin
taxa
richness, Shannon-W.ienerdiversity index
and Pielou
everulessindex
among regimes,rock
pools, periods and
times
through
nutrient
enrichment disturbances. Shannon-'Wiener diversityindex
and Pielou evenness index were powertransformed
(x2)
...37Table 3: Univariate analysis performed
with
afour-way
partrally hierarchicalANOVA
for
differencesin
taxa richness, Shannon-Wienerdiversity
index
and Pielou
evennessfollowing
drainingdisturbances..
...40CHAPITRE
2Table 4:
List
of the 38 sessile taxasampled
...59Table 5:
Univariate
analysis performedwith
an
asymmetricalANOVA for
differencesin
mean Euclidean distances (based on the percentage cover
of
benthic sessile taxa)in
controls versus disturbedrock
pools Disturbances were performedby
the additionof
boulders at two frequency (pulse, press) and intensity (low, high)
levels
...60Table
6: Univariate
analysis performedwith
an asymmetricalANOVA for
differencesin
mean Euclidean distances (based onthe
taxa compositionof
benthic sessile taxa)in
controls versus disturbed rock pools. Disturbances were performed by the additionof
XXVl
CHAPITRE
3Table 7: PCA output
for
(1) marine and (2) freshwatermesocosms..
...77 Table8:
Statistical resultsfor
the marine system. a) linear regression,b) ANCOVA with
trophic position
aspredictor
andc)
asymmetricaltwo-way
crossedANOVA for
themarine system
with
Number of taxa removed (no) and Sequence as main factors. Thevariability of
the factor
Sequence was thendivided
between 1) random versus non-randomextinction
sequence (R vsNR)
and2)
differences among random extinctions sequences (among random). P values in bold colrespond to significant results ... 80 Table 9: Statistical results for the freshwater system. a) linear regression, b)ANCOVA
with
trophic position
aspredictor
andc)
asymmetricaltwo-way
crossedANOVA for
thefreshwater system
with
Numberof
taxa removed(no)
and Sequence as main factors.The
variability of
the factor
Sequence was thendivided
between 1) random versusnon-random
extinction
sequence(R
vs NR)
and
2)
differences among
randomextinctions
sequences (amongrandom).
P
valuesin
bold
colrespondto
significantresults.
...81ANNEXE
Table 10:
Summarytable
of
the effect
of
functional
group
richnesson the
differentecosystem functions
studied...
..125
Table 1 1: Linear models partitioning the variance between linear group
richness,
nonlineargroup richness, identity effects and composition
effects.
...126
Table
12: Functional group contribution
to
ecosystemfunctions.
A
positive
coefficientindicates that
a
speciesis
contributingmore
to
the given function
than the average species,while
a
negativecoefficient
indicatesit
is
contributing
lessthan
average.Although global
identity
effect
is
significant
for
eachfunctions
(seetable
2),
boldLISTE
DESFIGURES
INTRODUCTION
Figure
1:
Représentationdu
rôle
des perfurbations dans l'assemblage dela
structure descommunautés
(tiré
deHilleRislambers et
a1.,2012). Les perturbations agissent àtitre
de
filtres
environnementaux, déterminant
I'importance
relative des
processusstochastiques
(dispersion, chance)
et
déterministes
(filtres
environnementaux et biotiques). Chaque lettre coffespond à une espècedifferente.
...3Figure
2
:
Changements
anticipés dans
la
probabilité d'occurrence
d'événements météorologiques extrêmes sousI'influence
des changements climatiques.Du
scénarioA
(ligne pointillée
;
condition
actuelle) au scénarioB
(ligne
pleine;
prédtction pour2050),la
valeur moyenne(xr
à xz) et la variabiliTé (écart-bpe ou largeur des courbes)des probabilités d'occurrences augmentent. (Tiré de Jentsch et a1.,
2007)....
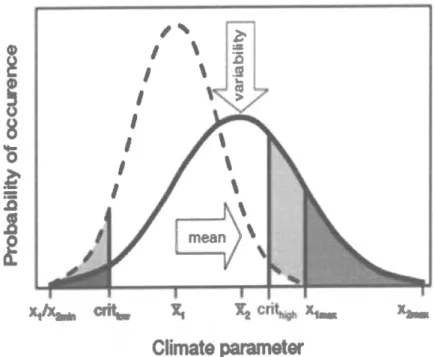
...8Figure
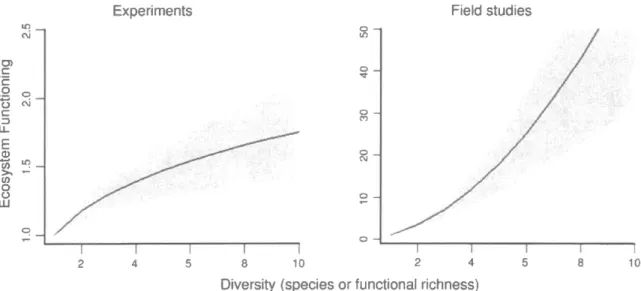
3:
Relation entre
la
biodiversité
et
le
fonctionnement des écosystèmesau
sein d'écosystèmes(i)
expérimentauxet
(ii)
naturels.Les
zones ombragées représententf
intervalle de confiance de 95% (tiré de Mora et al.2014)
...11Figure 4
:
Changementde
dominanceau
sein des
écosystèmesmarins
perfurbés. Les organismes de grandes tailles sont remplacés par des organismes de plus petites tailles(tiré de Pauly and
Chuenpagdee,Z}}Z)
...14Figure
5:
Schéma conceptuel
illustrant
la
chaine d'impacts entre
le
régime
desperturbations, la
biodiversité
etle
fonctionnement de l'écosystème. Les composantes en gris représentent les thèmes qui ne sont pas abordés dans cettethèse.
...18xxvlll
Figure
6
: Représentation graphique des hypothèses testées au chapitre2...
...22CHAPITRE
1Figure
7:
Non-metric multidimensional
scaling(nMDS)
ordinations.nMDS
were plottedfor
each regime
before and after the
beginning
of
disturbancesby
(a)
nutrient enrichment and(b)
draining.
High
:
black triangle;
Low
-
red square; Reference:
green
circle
"""'39
Figure 8: V/ithin-group dissimilarity
in
rock
pools disturbedby
nutrient
enrichment. The black vertical line represents the beginning of the disturbances. High-
nutrients added every diurnallow
tide;Low
-
nutrients added six times during the period, Reference:
no
nutrient added. Note that high within-group dissimilarityis
associatedwith
high betadiversity
"' 4I
Figure 9: Mean taxarichness (mean
+
SE) before and after the beginning of disturbancesby
draining. Before
-
black;After
-
cyan; High:
nutrients added every diurnallow
tide;Low
:
nutrients addedsix
times during
theperiod;
Reference:
llo
nutrient
added.Different
letters indicate statistically significant difference between regimes...42Figure 10:
Within-group
dissimilarity
in
rock pools
disturbedby
draining.
The
black
vertical line
representsthe
beginning
of
the
disturbances.High
:
draining
everydiurnal
low
tide;Low
:
draining six times during the period, Reference:
rto draining.Note that high within-group
dissimilarity
is associatedwith
high beta diversity ...44Figure
11:
Between-group dissimilarity
in
rock
pools
disturbedby
draining.
The
blackvertical line
representsthe
beginning
of
the
disfurbances.High
:
draining
everydiurnal
low
tide;Low
:
draining six times during the period; Reference:
rlo drainingXXIX
CHAPITRE
2Figure
12: Mean Euclidean distances (betadiversity;
+l-
SE)of
thetotal
percentage cover observedin
controls (white bars) andin
rock pools disturbed at pulse (grey bars) and press frequency regime(black
bars). Differences among regimes are shownfor
theentire community, then
for
the entire
communityin which
we
removedthe
canopy-forming
taxa,
the
erectedtaxa,
the turf-forming
taxa,
the
encrustingtaxa
and the sessileanimal
taxa. Greater mean Euclidean distancesrefer
to
greater beta diversity.Different
letters indicate statistically significant difference between regimes... ...62Figure
13:
Mean
Euclidean distances (betadiversity;
+l-
SE)
of
the
taxa
compositionobserved
in
controls (white bars) andin
rock pools disturbed at pulse (grey bars) and press frequency regime(black
bars). Differences among regimes are shownfor
theentire community, then
for
the entire
communityin
which we
removed thecanopy-forming
taxa,
the
erectedtaxa,
the turf-forming
taxa,
the
encrustingtaxa
and the sessileanimal
taxa. Greater mean Euclidean distances referto
greater beta diversity.Different
letters indicate statistically significant difference between regimes ... ...63CHAPITRE
3Figure
14: Schematic representationof
the experimental designfor
the marine ecosystem.Note that the experimental design
for
the freshwater ecosystem was identical, exceptfor
the numberof
species we removed(0,
1 ,2
and4)
and the numberof
independentcombinations of random sequences reproduced at each level
of
species richness (10in
the freshwater
ecosystem)....
...74Figure
15:
Effects
of
body size on
ecosystemfunctioning.
The
proxies
of
ecosystemfunctioning
illustrated
here are
a)
marine
multifunctioning,
b)
freshwaterphytoplankton biomass
and
c)
marine
encrusting algaenet production.
O
indicate grazers andC
indicateîoî-gtazers.
The y-axis refer to the effect size between valuesof
functioning
recorded
in
mesocomswith
removals
and
values
of
functioningxxx
Figure 16: Effects
of
taxaremoval on ecosystem functioning according to random and non-randomextinction
sequences. Theproxies
of
ecosystemfunctioning illustrated
hereare
a)
periphyton
dry
mass
b)
marine
multifunctioning
and
c)
freshwaterphytoplankton biomass. White boxplots indicate non-random extinction sequences and grey boxplots indicate random extinction sequences. The y-axis refer to the effect size
between values
of
functioning
recordedin
mesocomswith
removals and valuesof
functioning recorded in reference mesocosms (no removal)...
..-...82Figure
17:
Différentes
hypothèses quantaux
processus (stochastiquesvs
déterministes) dominantsqui
sous-tendent la relation entre le régime de perfurbations etla
diversitébêta
""""""""
93ANNEXE
Figure
18:
Simplified
schematic representationof
the
a)
marine and
b)
freshwaterecosystems
studied...
"""
119Figure
19: Responseof
a)
freshwater andb)
marine
ecosystemfunctions
to
functional
group richness. Boxplots indicated interquartile variance amongst richness treatments
with
black bold line indicated the median. Only significant functions are shown.....I24
Figure
20:
Relationships betweenfunctional
groupindividual
contribution and ecosystemmultifunctionality
for
a) freshwater and b) marine ecosystem(first
and second axesof
the
principal
component analysis-
PCl
andPC? respectively). Groups are shownin
INTRODUCTION GENERALE
BIoorvgRSITE
On a identifié
près
de
I,2
million
d'espècessur Terre. C'est
bien
peu
lorsqu'on estime quele
nombretotal
d'espèces peuplantla
planètes'élèverait
àL8,7
millions.
La grandemajorité d'entre
elles resterait donc toujoursà
être répertoriées.En
milieu
marin uniquement, 91% des espècesn'auraient
toujours pas été découvertes, cequi
représente plusde2 millions
d'espèces(Mora
etal.,2}ll).
C'est très
souvent
en
ces termes, c'est-à-dire
en
nombre
d'espèces,que
lacommunauté
scientifique
discute de
biodiversité.
En
réalité,
la
signification
du
motbiodiversité est beaucoup plus large. Elle réfère à la diversité du vivant et
inclut
la diversité des espèces,la
diversité des gènes etla
diversité destraits
fonctionnels (Cardinale et al., 2012). Notons que les traits fonctionnels sont des traits morphologiques, physiologiques ou phénologiques propres à I'espèce (p.ex.taille, rôle
trophique),qui influencent
sa réponse aux conditions environnementales (Hooper et a1.,2005). La biodiversité peut être décrite en termesde
richesse(nombre d'unités vivantes), mais
égalementen
terme
d'équitabilité (dominance)et
d'hétérogénéité(dissimilarité)
entre les unités vivantes (Magurran, 2004;Hooper et a1.,2005; Cardinale et al., 2012).
En
milieu
naturel,
Ia
diversité des
espècesse
décline
en
differents
niveauxd'organisation.
D'abord,
ofl
appelle population
un
ensembled'individus d'une
même espèce dansun
environnement donné.Une
communauté estformée
lorsqueplus
d'une population d'espèces differentes s'associe, cohabite et interagit entre elles. Iæs interactionssont dites intraspécifiques
si
elles
surviennent entre deuxindividus ou
plus à
I'intérieur
d'une
même
population, alors que
les
interactions
sont
interspécifiques lorsqu'ellessurviennent
entre
au
moins
deux
espèces.Il
existe plusieurs types
d'interactionsinterspécifiques.
Les
interactions
élémentaires
directes incluent
le
mutualisme,
Ia compétition et la prédation(Morin,
2011). Les communautés interagissent également avecun
ensemblede
facteurs abiotiques,tels que
la
lumière,
la
température,Ia
pression, les sédiments et les nutriments.Un
écosystème englobe I'ensemble des interactions biotiqueset abiotiques dans un environnement donné (Thurman et
Trujillo,2004;
Morin,
20ll).
Au
sein des
écosystèmes,la
structure des communautés(composition,
abondancerelative,
biomasseou
recouwement des espèces)n'est
pas
statique dansl'espace
et
le temps. Cettevariation
émerge dela
combinaison de plusieurs mécanismes d'assemblagesdes communautés
(Gravel
et
a1.,2011).La
composition des communautés seraiten fait
déterminée
parun
ensemble de processus stochastiqueset
déterministesqui
surviennentsimultanément dans
un
système(Lepori
et
Malmqvist,
2009;
Chaseet
Myers
2011). Brièvement,tel
que résumépar Chase et Myers (2011), les théories stochastiques suggèrentque
les
chancesde
colonisation,
les
extinctions
aléatoires
et
la
dérive
écologique détermineraient largement les patrons de biodiversité(Hubbell,
2001), alors que les théories déterministes soulèventf
importance des processus locaux, à la base du modèle de niche,tels
que
les
interactions biotiques, abiotiques
et
les
compromis
interspécifiques.L'importance
relative des
processus
stochastiqueset
déterministes dépendrait
desperturbations,
qui
selonleur
sévérité, agiraient àtitre
defiltres
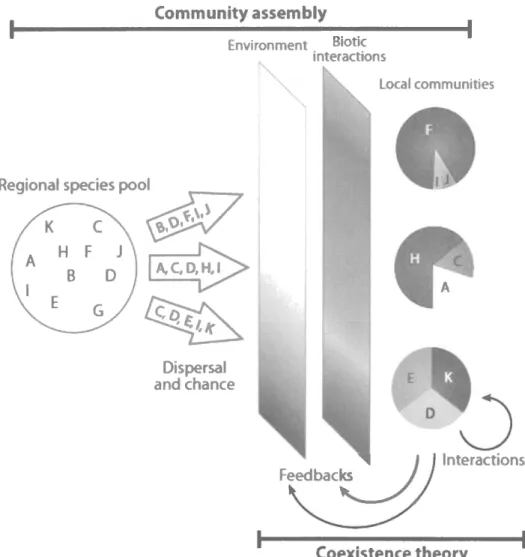
environnementaux(Fig.
1).Les processus stochastiques tendraient à dominer au sein d'environnements non perturbés,
alors que les
processus déterministes prévaudraienten
milieux
perfurbés (Chase, 2007; Lepori andMalmqvist,
2009). Les perturbations, partie intégrante des écosystèmes naturels, sont donc une source majeure d'hétérogénéité spatiale et temporelle dansla
structure et la dynamique des communautés naturelles (Sousa, 1984).PeRTuRBATIONS
plusieurs
définitions du
terme perturbation
ont
été proposéesdans
la
littérature(Grime,
1977;Bender,
1984; Sousa,1984;
White
etPickett,
1985).Ici,
une perturbationréfère
à
(
tout
événement,défini
dans
le
temps,
qui
peut altérer
la
structure
d'un
écosystème,d'une
communauté
ou
d'une population
et
modifier
les
ressources, ladisponibilité du
substratou
encore I'environnement physique D(White et Pickett,
1985).Une
perturbation
peut être biologique (p.e*.
prédation)
ou
physique
(p.ex.
tempête),naturelle
(p.ex.
inondation,
glace)
ou
anthropique
(p.ex. rejet d'égout,
destruction
de I'habitat, surpêche, introduction d' espèces invasives).Community
assembly
Environment Biotic interactions Local communities Dispersal and chance Interactions Regional species pool^N
(l
',F,')4>
vq\
'
Coexistencetheory
Figure
l:
Représentation durôle
des perturbations dans l'assemblage dela
structure descommunautés
(tiré
deHilleRislambers
et
al., 2012). Les perturbations agissent àtitre
defiltres
environnementaux, déterminantI'importance relative
des processus stochastiques(dispersion, chance) et déterministes
(filtres
environnementaux et biotiques). Chaque lettre correspond à une espèce différente.A HF
)4
Les perturbations affectent les communautés autant positivement que négativement.
D'une
part, les
perturbationspeuvent favoriser
la
coexistence des espècesau
sein
descommunautés
par
divers mécanismes.Par
exemple, une perturbationpar
prédation peutprévenir
la
monopolisation des
ressources environnementalespar une
seuleet
uniqueespèce
(paine 1966;
I97L;
Dayton,
L97l).
La
coexistenceest
aussi possible
via
uncompromis interspécifique colonisation-compétition.
D'après
I'hypothèse
de
la perturbation intermédiaire, un niveau modéré de perturbations optimiserait ce compromis,maximisant
la
diversité
(Grime,
1973;Connell,
1978).À
la
basede
cette
hypothèse,Connell (1978) stipule qu'à trop faible fréquence ou intensité de perturbations, les meilleurs
compétiteurs
du milieu
supplantentles
espèces arD( capacitésde
compétition
moindre.L'effet
se
traduit
alors
par
un
systèmeà
l'équilibre, peu diversifié.
À
I'opposé2 uneperturbation
trop
fréquenteetlou
intense laisse plusieurs espaces dénudés, n'avantageant que les espèces colonisatrices,qui
ont
un
meilleur taux de
croissance intrinsèque et unecapacite
de
dispersion
supérieure.Une
fréquence
ou
une
intensité
de
perturbationsintermédiaires favoriseraient
alors
la
coexistenceentre
les
espècescolonisatrices
et compétitrices, en perturbant la hiérarchie de compétition, augmentant le taux de mortalité etlaissant des
espacesdisponibles
pour
le
recrutementdes
espècesmoins
compétitives (Svensson et a1., 2007).Iæs perturbations peuvent aussi
modifier
négativement les paramètres physiques deI'environnement,
ne
permettant
qu'à
certaines espècesparticulières
de
surviwe.
Parexemple,
le
paradigme proposé
par
Pearsonet
Rosenberg(1978) décrit
les
étapessuccessives de
la
distribution
d'espèces benthiquesvivant
dans des sédiments meubles deplus en plus riches en
matière
organique.D'abord
diversifiées,
les
communautés setransforment peu à peu, pour
n'être
représentées que par quelques espèces seulement, très abondantes et capables de tolérerun
enrichissement important en matière organique. Lesperturbations anthropiques seraient,
de
plus, en
grandepartie
responsablesde
la
perted'espèces observée actuellement, dont
le taux d'extinction
seraitde
100à
1000fois
plus élevé que ceux estimés dans le registre fossile(Pimm
etal.,
1995). Une évaluation efficacede
I'effet
de la perte de biodiversité suite à l'occurrence de perturbations est importante sion veut, ultimement, pouvoir en minimiser les impacts négatifs associés.
L'évaluation
des effets des perfurbations surla
biodiversité est, laplupart
du temps, éfudiée en comparantle
nombre d'espèces contenues dansun
échantillon, c'est-à-dire ladiversité
alpha(Whittaker,
1960), entre sites perturbéset
sites témoins(non
perturbés).Cette approche soulève deux problèmes fondamentaux
(Underwood,
1994).D'abord,
lenombre
moyen
d'espèces dansune
communautévarie
naturellement dansle
temps. Ce<<bruit
de
fond> rend
souvent impossible
la
détection des perturbations
anthropiquesuniquement
sur
la
base
du
nombre
d'espèces observées. Deuxièmement,les
patrons temporels de la richesse spécifiqued'une
communauté varient énormémentd'un
endroit àI'autre.
Statistiquement,il y
aLLradonc
interaction entre
les
variations
temporelles et spatialesdu
nombre
d'espèces dansla
communauté,ce qui
augmented'autant
plus
ladifficulté
à déceler la présence de perturbations àpartir
des patrons de richesse spécifique.Il
n'existe aucun raisonnement scientifique de considérer seulement unediminution
dans lenombre d'organismes comme
effet
néfaste
des
perturbations
sur
la
biodiversité (Underwood,l99l).
Rappelons que la biodiversité peut non seulement être décrite en termede nombre d'espèces, mais également en terme
d'équitabilité et
d'hétérogénéité entre les espèces(Magunan,2004;Hooper
eta1.,2005; Cardinale et al., 2012).En
réalité,
les
perturbations affecteraient autant I'hétérogénéité
que
le
nombre d'espèces lui-même (Underwood,I99l;
1992; 1994; Chapman etal.,
1995; Bertocciet
al.,2005).
Une des
approchesprivilégiées afrn d'estimer
l'impact de
perturbations
surl'hétérogénéité consiste à comparer
la
diversité bêta entre communautés perturbées et non perturbées. Anderson et al. (2011) différencient deux types de diversité bêta. Premièrement,la
diversité bêta peut référer
à
un
changementdirectionnel dans
la
structure
de
la communauté d'une unité d'échantillonnage à 1'autre le longd'un
gradient spatial, temporelou
environnemental.La
diversité
bêtapeut
aussiréférer à une notion de variation,
nondirectionnelle,
de
la
structure
de
la
communauté
parmi
un
ensemble
d'unités d'échantillonnage.C'est
cette
définition
qui
sera retenueici.
Une
différence dans
ladiversité bêta d'une communauté peut être attribuable à
i)
unemodification
de l'abondance totale des individus à un endroit donné,ii)
unemodification
du nombre total d'espèces,iii)
un
changement dans leratio
variance/moyenne pour des espèces particulièresou
encore àiv)
un
changementdans
I'identité des
espèces présentesau
sein d'une
cofirmunauté(Warwick
et Clarke,L993;
Chapman et al. 1995).Les tests basés sur les changements de l'hétérogénéité spatiale et temporelle seraient
plus
sensibles etplus
appropriéspour
estimer les impacts environnementaux que les tests baséssur
des
comparaisonsde
moyennes
(p.
ex.
diversité alpha)
ou
de
centroïdes(Lindegarth
et
aI. 2000). Les variationsde
la
structured'une
communauté révèlent aussibeaucoup d'informations (Benedetti-Cecchi,2003). Elles peuvent
fournir
des indices quantagx
processusqui
structurent
les
écosystèmesà
plusieurs
échelles
spatiales. Ellespermettraient aussi
de
prédire des
changementsd'état
à venir
dansla
dynamique desécosystèmes sous
f
influence des perturbations, alors que les communautés tendraient vers uneplus
grande hétérogénéité spatiale à l'approched'un point
de transition (Fraterrigo etRusak, 2008; Scheffer et a1.,2009).
RTCNNS DE PERTURBATIONS
Une
perturbationpeut être décrite,
en
fonction
de
sesdifftrentes
caractéristiques appelées régimes de perturbations. Ony
précisera alors la magnitude (intensité et sévérité) de même que les composantes temporelle (p. ex., fréquence) et spatiale (p. ex.,taille
d'unezoîe
perturbée)de
cette perturbation.
La
définition de
chacun des
termespeut
êtreretrouvée dans