THESE
en vue de l’obtention du
DOCTORAT DE L’UNIVERSITE DE TOULOUSE délivré par l’Université Toulouse III - Paul Sabatier
Discipline : Hydrobiologie présentée et soutenue par Stéphanie BOULÊTREAU le 28 septembre 2007
Titre :
Déterminisme des fonctions d’accrétion et de détachement du biofilm
phototrophe en milieu naturel : études expérimentale et numérique des
facteurs de contrôle de la biomasse en rivière.
Directeurs de thèse :
Sabine SAUVAGE et José-Miguel SÁNCHEZ-PÉREZ
JURY
J.-L. ROLS, Professeur, Université Toulouse III , Président
A. ELOSEGI, Professeur, Université de Bilbao, Espagne , Examinateur
F. GARABETIAN, Professeur, Université de Bordeaux I , Examinateur
P. MARMONIER, Professeur, Université de Rennes , Examinateur
M. POULIN, Ingénieur de Recherche, Ecole des Mines Paris , Rapporteur
S. SABATER, Professeur, Université de Girona, Espagne , Rapporteur
J.-M. SÁNCHEZ-PÉREZ, Directeur de Recherche, CNRS , Directeur de thèse
Je voudrais tout d’abord remercier MM. Jean-Luc ROLS (Professeur, Université Paul Sabatier) et Eric CHAUVET (Directeur de Recherche, CNRS), respectivement directeurs du LEH (Laboratoire d’Ecologie des Hydrosystèmes) et d’EcoLab (Laboratoire d’écologie fonctionnelle), pour m’avoir successivement accueillie au sein de leur laboratoire.
Je tiens à remercier mes directeurs de thèse, Mme Sabine SAUVAGE (Ingénieur de Recherche, CNRS) et M. José-Miguel SANCHEZ-PEREZ (Directeur de Recherche, CNRS), qui m’ont offert l’opportunité de réaliser cette thèse et se sont rendus très disponibles au cours de ce travail.
Je tiens à remercier les personnes qui ont accepté de juger ce travail : M. Sergi SABATER (Professeur, Université de Girona, Espagne), M. Michel POULIN (Ingénieur de Recherche, Ecole des Mines de Paris), M. Arturo ELOSEGI (Professeur, Université de Bilbao, Espagne), M. Frédéric GARABETIAN (Professeur, Université de Bordeaux) et M. Pierre MARMONIER (Professeur, Université de Rennes).
Je remercie aussi particulièrement M. Arturo ELOSEGI pour m’avoir accueillie au sein de son équipe du Département de Biologie Végétale et d’Ecologie de l’Université de Bilbao pendant mon séjour ATUPS et pour avoir aussi efficacement fait avancer ce travail.
Je remercie également le personnel de l’Institut de Mécanique des Fluides de Toulouse qui a participé à la mise en place et à la réalisation financière et technique de l’expérience en canal de laboratoire : MM. Alexandre BEER, Yvan BERCOVITZ, Grégory DHOYE, Noël DOLEZ, Olivier EIFF, Serge FONT et Frédéric MOULIN.
Je tiens à remercier Mme Claude MUR et M. Daniel DALGER pour leur professionnalisme et leur contribution inconditionnelle à l’ensemble des analyses chimiques de ce travail. Merci beaucoup à M. Yvan NICAISE pour sa disponibilité et son investissement dans le traitement des échantillons de biologie moléculaire. Merci à M. Mehdi SELLALI pour son implication lors de l’expérience en canal.
beaucoup apprécié de partager les discussions scientifiques et philosophiques post prandiales. A mes collègues gaulois de début ou de fin de thèse, Emilie, Pierre, Sandrine, Arthur, Malvina, les Freds, Sammy, Sylvain, Daniel, Yvan, Jean-Louis, Jo, Armelle, Anthony, Cristina et Julien qui ont contribué à la bonne ambiance au laboratoire.
Un merci tout particulier au Professeur « Garabet » pour s’être investi autant, m’avoir conseillée, fait confiance et tellement appris tout au long de ces années et ce, malgré mon lourd passé d’ingénieur.
Une pensée amicale pour Amaia et Laurent, mes vrais faux amis toulousains du Pays Basque (le vrai !) et de l’Ardèche, que j’ai rencontrés et appris à mieux connaître pendant ces années. Une pensée aussi pleine de repentance pour les termites de Laurent.
Liste des abréviations ………
1
Liste des figures ………. 2
Liste des tableaux ………..
6
Avant-propos ……….
7
Chapitre 1 : Contexte général et problématique ...……….
10
1 – Les biofilms épilithiques : de la rivière à l’agrégat ...………
11
1.1 – Définitions ……….. 11
1.2 – Conditions de développement ……… 13
1.3 – Rôle fonctionnel dans les écosystèmes lotiques ……… 15
1.4 – Dynamique temporelle de la biomasse ……….. 16
1.5 – Intérêt pour la bioindication ………... 19
2 – Les modèles mathématiques : de la description à la prédiction ………
21
2.1 – Définitions ……….. 21
2.2 – Intérêts pour l’écologie ……….. 22
2.3 – Démarche de modélisation ………. 23
2.4 – Limites et contraintes d’application ………... 26
3 – Objectifs de l’étude ………...
28
3.1 – Problématique ……… 28
3.2 – Questions et démarche ………... 30
Chapitre 2 : Méthodologie générale ………
32
1 – Expérimentation numérique ………..
33
1.1 – Présentation des séries temporelles ………... 34
1.1.1 – Séries temporelles de l’Agüera ………... 34
Sites d’études………. 34
Echantillonnage……… 36
Données disponibles ……… 36
Données disponibles ……… 39
1.2 – Revue des modèles existants ……….. 40
1.3 – Description du modèle ………... 43
1.3.1 – Formulation initiale ………. 43
1.3.2 – Développement………... 44
1.4 – Résolution numérique ……… 45
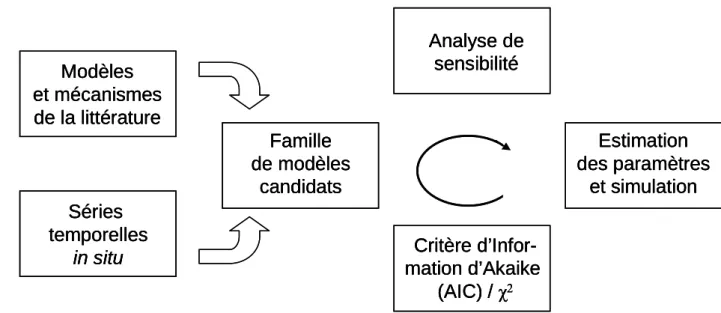
1.5 – Sélection de modèle et identification des paramètres ……… 45
1.5.1 – Critère d’Information d’Akaike (AIC) ……… 46
Identification des paramètres sans contrainte ………... 46
Identification des paramètres avec contrainte……… 47
Interprétation des paramètres……… 47
1.5.2 – Critère du χ2 ………. 48
2 – Expérimentations in situ ………
49
2.1 – Expérimentation en milieu contrôlé ………... 49
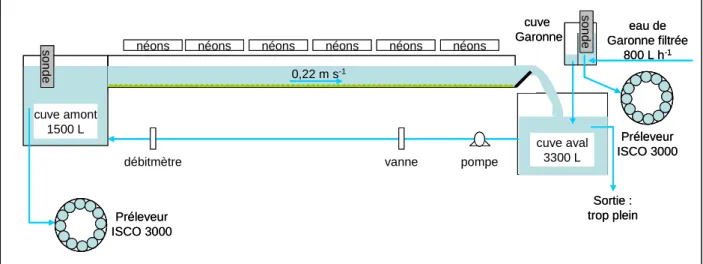
2.1.1 – Dispositif expérimental ………... 49
Caractéristiques du montage hydraulique ……….. 49
Conditions nutritives et physico-chimiques………. 50
Caractéristiques de l’écoulement ………. 51
Conditions d’éclairement……… 51
Caractéristiques des substrats ………... 52
2.1.2 – Déroulement de l’expérience………... 53
Phase d’ensemencement ………. 53
Echantillonnage et conditionnement du biofilm………. 54
2.1.3 – Analyses ………... 55 Chlorophylle a ……….. 55 Métabolisme………… ……….. 55 MSSC et MS ………. 57 Composition algale ………... 57 Composition bactérienne………. 58 Analyse de l’eau ………... 58
MSSC et MS ……….. 61 Chlorophylle a ……….. 61 Dénombrements algaux ……….. 61 Invertébrés ……… 61 Analyses de l’eau ………. 61 Débits ………. 62 Rayonnement solaire ………... 62
Chapitre 3 : Importance des perturbations hydrodynamiques dans la
dynamique spatio-temporelle du biofilm épilithique : vers un modèle
de prédiction ………..
63
1 – Contexte et objectifs ………..
64
2 – Principaux résultats et discussion ………..
66
3 – Conclusion ……….
69
4 – Publication ………. 70
Chapitre 4
:
Importance du détachement autogène dans la
dynamique spatio-temporelle du biofilm épilithique
: le modèle
comme outil de recherche ……….
88
1 – Contexte et objectifs ………..
89
2 – Principaux résultats et discussion ………..
91
3 – Conclusion ……….
93
4 – Publication ………. 94
Chapitre 5 : Dynamique de la biomasse en écoulement contrôlé : vers
une évolution autogène de l’agrégat ? ……….
106
1 – Contexte et objectifs ………..
107
5 – Publication ………. 120
Chapitre 6 : Comparaison interannuelle de la dynamique de la
biomasse épilithique en milieu naturel ………
152
1 – Contexte et objectifs ………..
153
2 – Résultats et discussion ………...
154
2.1 – Caractéristiques de la série temporelle de 2004-05-06 ………….. 154
2.2 – Comparaison interannuelle des patrons de biomasse ………. 161
2.3 – Comparaison interannuelle des conditions environnementales …. 163 2.4 – Comparaison taxonomique des assemblages algaux ……….. 164
3 – Conclusion et perspectives ………
167
Conclusion générale ………..
171
AI ou IA : Autotrophic index ou index d’autotrophie AIC : Akaike information criterion
APHA : American public health association
AR : Accretion rate
BIC : Bayesian information criterion
COD : Carbone organique dissous
DBL : Diffusive boundary layer
DGGE : Denaturing gradient gel electrophoresis
DIREN : Direction régionale de l’environnement EPS : Exopolymeric substances
EPU : Eucaryotes photosynthétiques unicellulaires
Fr : Nombre de Froude
GPP : Gross Primary Production
HPLC : High performance liquid chromatography
IC ou CI : Inorganic content ou contenu inorganique IMFT : Institut de mécanique des fluides de Toulouse
MES : Matière en suspension
MS : Matière sèche
MSSC : Matière sèche sans cendre
NPP : Net Primary Production
NTU : Nephelometric turbidity unit
OTU : Operational taxonomic unit
PB : Peak of biomass
QMJ : Débit moyen journalier
RCC : River Continuum Concept
PAR : Photosynthetically active radiation
PCR : Polymeric chain reaction
RSS : Residual sum of squares
R : Respiration
SC : Schwarz criterion
Figure 1.1. Observations du biofilm épilithique à l’échelle microscopique (a), du
galet (b) et du tronçon de rivière (c) ………... 12
Figure 1.2. Courbe théorique de croissance (d’après Biggs 1996) ………... 17
Figure 1.3. Représentation schématique de la démarche de modélisation (d’après
Jorgensen 1983) ………... 24
Figure 1.4. Spectre de la complexité d’un modèle (adapté de Snowling & Kramer
2001). La flèche représente la gamme de complexité possible pour un modèle, de plus en plus simple (à gauche) et de plus en plus complexe (à droite) ………... 27
Figure 1.5. Schéma illustrant la relation entre complexité – incertitude – sensibilité
d’un modèle. Plus la complexité d’un modèle augmente, plus sa sensibilité augmente et plus son incertitude diminue. Il existe un modèle de complexité intermédiaire qui optimise la relation complexité – incertitude – sensibilité (d’après Snowling &
Kramer 2001) ………... 28
Figure 2.1. Schématisation de la démarche d’expérimentation numérique mise en
oeuvre pour obtenir un modèle de dynamique de la biomasse épilithique ………. 33
Figure 2.2. Vues des sites amont (site 2, à gauche) et aval (site 9, à droite) de
l’Agüera ………... 34
Figure 2.3. Débit moyen journalier (m3 s-1) mesuré dans le site 9 (aval) lors des périodes d’échantillonnage de 1990-91 (a), 1992-93 (b) et 2001-02 (c) ………... 35
Figure 2.4. Vues prises en rive droite des sites amont (Aouach, à gauche) et aval
Figure 2.6. Photographie et schéma de l’agencement des substrats dans le canal ...….. 53
Figure 2.7. Schéma du dispositif de mesure de la production et de la respiration
(schéma modifié de J. Meillon) et photographie de la chambre d’incubation des
substrats ……….. 56
Figure 4.1. Illustration d’un galet érodé par le détachement autogène ……….. 91
Figure 5.1. Photographies des substrats colonisés après 4 semaines (a), 10 semaines
(b), 12 semaines (c) et 14 semaines (d) de croissance. Le diamètre d’un substrat est de
4 cm ………. 112
Figure 5.2. Evolutions temporelles de (a) la dérive moyenne du biofilm exprimée en
mg MSSC L-1 (g MSSC m-2)-1, (b) le contenu chlorophyllien et (c) le contenu inorganique (en % de cendres). L’évolution de la biomasse chlorophyllienne (moyenne ± ES) en mg chlorophylle a m-2 est reportée sur chaque graphique sur l’axe
des ordonnées de droite (cercles gris) ………... 116
Figure 5.3. Résultats des simulations en chlorophylle a (colonne de gauche) et MSSC
(colonne de droite). Les graphiques comparent les évolutions de la biomasse (moyenne ± ES) observée (carrés noirs) et de la biomasse simulée selon l’hypothèse 1 d’« altération physiologique » (ligne noire) ou l’hypothèse 2 d’ « abrasion » (ligne grise). Les tableaux indiquent la signification de la variable de forçage Sed(t) du terme de perte ainsi que les valeurs des paramètres et de l’ajustement de la combinaison optimale ………... 119
Figure 6.1. Evolutions temporelles de la biomasse épilithique (moyenne ± ES)
exprimées en g MSSC m-2 (a, a’), g MS m-2 (b, b’) et mg chl a m-2 (c, c’) des séries temporelles de 2004-05-06 (colonne de gauche) et de 2001-02 (colonne de droite) à l’Aouach (site amont, Garonne). Noter que les échelles des axes des ordonnées sont
global journalier (b, b’), de la température de l’eau (c, c’), de la concentration en phosphore (d, d’) et de la biomasse épilithique (e, e’) des séries temporelles de 2004-05-06 (colonne de gauche) et de 2001-02 (colonne de droite) à l’Aouach (site amont, Garonne). Noter que les échelles d’une série à l’autre sont identiques, excepté pour la
biomasse (e, e’) ………... 157
Figure 6.3. Densités totales, des collecteurs filtreurs, des collecteurs assembleurs, des
brouteurs, des prédateurs et des déchiqueteurs lors des prélèvements des 1, 13 et 26
juillet 2005 ...………... 158
Figure 6.4. Evolutions temporelles du contenu chlorophyllien du biofilm en % (a) et
du contenu intracellulaire en chlorophylle a en mg chl a (ind.)-1 (b). La biomasse
épilithique (en g MSSC m-2) est reportée sur le second axe des ordonnées du graphique (a). Le rayonnement global journalier (en J cm-2) est reporté sur le second
axe des ordonnées du graphique (b) ……….... 160
Figure 6.5. Comparaison entre la biomasse épilithique observée (moyenne ± ES) et la
biomasse épilithique simulée obtenue en appliquant la paramétrisation optimale du modèle à trois paramètres confronté à la série temporelle de 2001-02 : µmax,0 = 1 d-1,
kinv,B = 3.8 et cdet = 0.0002 d-1………... 161
Figure 6.6. Evolutions temporelles de l’abondance relative (%) des biovolumes des
principaux groupes algaux (a, a’) et du nombre de taxa (b, b’) lors des séries temporelles de 2004-05-06 (colonne de gauche) et de 2001-02 (colonne de droite). R : Rhodophycées ; C : Cyanophycées ; VF : algues vertes filamenteuses ; VNF : algues vertes non filamenteuses ; DF : diatomées filamenteuses ; DC : diatomées coloniales ; DUL : diatomées unicellulaires libres et DUF : diatomées unicellulaires fixées. La
biomasse épilithique en g MSSC m-2 est reportée sur les graphiques (a) et (a’) ……… 166
Figure 6.7. Distribution des années en fonction de la valeur du couple durée moyenne
et amplitude moyenne des périodes de basses eaux. Le couple est défini à partir des indices hydrologiques décrits par Richter et al. (1998). La durée de la période de
période de basses eaux correspond au minimum de l’amplitude des QMJ au cours de
périodes de 90 jours ……… 167
Figure 6.8. Evolution temporelle du débit moyen journalier QMJ (m3 s-1) entre
Tableau 2.1. Synthèse comparative des caractéristiques des modèles de périphyton de
la littérature ………... 42
Tableau 3.1. Tableau récapitulatif des résultats de la sélection du « meilleur » modèle
dans les 13 situations testées (1ère colonne). Les situations 90-9 et 92-7 n’ont pas été testées. La présence d’une croix dans une colonne indique que le processus de la colonne est activé. Exemples : le meilleur modèle de la situation 90-2 est constitué d’un terme de croissance, limitée par l’épaisseur et d’un terme de détachement hydrodynamique continu ; le meilleur modèle de la situation 01-9 est constitué d’un terme de croissance et d’un terme de détachement hydrodynamique catastrophique (activé lorsque le débit Q est supérieur au débit critique Qcr) ……… 67
Tableau 6.1. Caractéristiques structurelles globales du biofilm épilithique lors des
séries temporelles de 2004-05-06 et de 2001-02 ……….... 162
Tableau 6.2. Caractéristiques environnementales moyennes des prélèvements lors
des séries temporelles de 2004-05-06 et de 2001-02. M.E.S. : Matières en Suspension ; C.O.D. : Carbone Organique Dissous ; Vf : vitesse au fond ; Hf : hauteur
La pierre glissante qui nous fait prendre un bain involontaire dans le ruisseau que nous voulions traverser, le réservoir d’eau de la cafetière expresso constamment visqueux, les dents déjà râpeuses peu après leur brossage sont autant de situations que nous devons à la croissance des microorganismes qui colonisent toutes sortes de surfaces, généralement sous la forme de communautés appelées biofilms. Ces derniers sont de tailles très variables, de la microcolonie de quelques millimètres à l’agrégat de plusieurs centimètres d’épaisseur dans lesquels les microorganismes sont englobés dans une matrice visqueuse. La forme d’existence offerte par le biofilm est particulièrement avantageuse puisqu’elle assure notamment une protection vis-à-vis des brouteurs et des biocides, une meilleure résistance aux forces physiques comme le courant ou aux situations de stress comme le dessèchement et facilite les interactions métaboliques.
Dans les ruisseaux ou rivières, le biofilm épilithique se développe à l’interface entre la colonne d’eau et les galets ou rochers. Il s’agit d’un assemblage constitué de microorganismes appartenant à des groupes aussi différents que les bactéries, les protozoaires, les algues, les invertébrés, etc. En tant que producteurs de biomasse et décomposeurs, les biofilms épilithiques constituent un maillon vital du réseau trophique. Ils participent au fonctionnement hydro-écologique de la rivière (Battin et al. 2003), en particulier au niveau des transformations de l’azote (Teissier et al. 2007) ou du fonctionnement des communautés benthiques (Feminella & Hawkins 1995). Dans les secteurs de cours d’eau où ils représentent la seule biomasse microbienne, le bilan de matière est conditionné par la production, l’arrachement et la dégradation de la matière organique issue de ce biofilm. Le fonctionnement biogéochimique de ces secteurs est souvent directement associé à la dynamique spatiale et temporelle du biofilm épilithique et aux mécanismes qui le structurent.
Le développement du biofilm épilithique en rivière est une conséquence de conditions hydrodynamiques (vitesse du courant) et géomorphologiques (faciès dominants, hauteur d’eau, stabilité des galets) particulières. Certains tronçons de rivières (zones torrentielle et semi-torrentielle) ne sont effectivement propices qu’au développement d’une communauté fixée car les communautés libres sont « lessivées » par des temps de transfert très courts. C’est le cas de la Garonne sur une partie de son cours entre sa source et sa confluence avec le Tarn, en amont et en aval de l’agglomération toulousaine. Celle-ci est effectivement caractérisée par de faibles concentrations en chlorophylle a dans la colonne d’eau : comprises entre 2 et 20 µg L-1, les concentrations maximales sont rencontrées dans les zones de retenues artificielles (Eulin 1997) et, dans les autres stations, la concentration est le plus souvent inférieure à 10 µg L-1. Il a par ailleurs été démontré que cette chlorophylle a est
principalement d’origine benthique (Améziane et al. 2003). La Garonne constitue à ce titre un modèle d’étude du biofilm épilithique.
La grande variabilité des communautés microbiennes qui composent les biofilms, et tout particulièrement les biofilms épilithiques, et la multitude des processus biologiques et physico-chimiques qui s’y déroulent induisent une grande complexité structurelle. Cette complexité rend leur étude expérimentale particulièrement difficile. Les modèles mathématiques constituent alors des instruments d’intérêt pour exprimer de façon quantitative la vision que nous possédons de ces systèmes et d’en vérifier la pertinence par comparaison avec des résultats expérimentaux. Ces modèles sont aussi un outil de changement d’échelle permettant de transférer les connaissances acquises au niveau de l’agrégat au fonctionnement de l’écosystème.
C’est dans ce cadre général que nous allons nous intéresser aux mécanismes qui structurent la dynamique spatio-temporelle du biofilm épilithique en milieu naturel avec pour angle d’attaque principal la contribution de l’outil modélisation dans la connaissance de ces mécanismes. Pour aborder cette question, ce manuscrit se structure en 6 chapitres :
• Le chapitre 1, qui s’appuie sur une base bibliographique, précise le contexte général autour des mots-clés « biofilms épilithiques » et « modèles mathématiques » avant de définir la problématique et les questions scientifiques auxquelles ce mémoire répond. Le biofilm épilithique est présenté du point de vue de sa structure, de sa dynamique en milieu naturel et de son rôle au sein des écosystèmes aquatiques. Les modèles mathématiques sont appréhendés du point de vue de la démarche de modélisation, des intérêts et limites de leur utilisation en écologie.
• Le chapitre 2 détaille l’ensemble des méthodologies relatives à la démarche de l’expérimentation numérique ainsi que les dispositifs expérimentaux, protocoles d’échantillonnage et analyses effectués.
• Les chapitres suivants présentent et discutent les résultats de ce mémoire : les chapitres 3 et 4 sont consacrés à la mise en œuvre de l’expérimentation numérique au service de la connaissance des processus qui contrôlent l’évolution de la biomasse épilithique en milieu naturel. Les chapitres 5 et 6 analysent et discutent les résultats de deux expérimentations complémentaires ayant pour objectifs de préciser, en milieu contrôlé (canal de laboratoire) certains processus du modèle et d’explorer la variabilité interannuelle de la dynamique de biomasse épilithique (in situ).
Chapitre 1
Contexte général et problématique
1 – Les biofilms épilithiques : de la rivière à l’agrégat
1.1 – Définitions
Dans les secteurs de rivière ou de fleuve à forte hydrodynamique (torrentiels et semi torrentiels), le temps de résidence des masses d’eau très court est incompatible avec la croissance d’une biomasse microbienne en suspension à l’origine de processus planctoniques (Reynolds et al. 1994). Les interfaces liquide-solide comme les supports rocheux (galets, blocs, roche mère ou matériau de construction), les sédiments ou les macrophytes constituent alors des supports favorables à l’installation et au développement des microorganismes. Ces microorganismes forment une couche visqueuse de quelques micromètres à quelques centimètres d’épaisseur et de couleur brunâtre ou verdâtre.
Associés à des particules détritiques et enchevêtrés dans une matrice d’exopolymères (EPS, exopolymeric substances), ces microorganismes adhérant à une surface submergée ou soumise à un environnement aqueux forment un film biologique appelé biofilm (Costerton et al. 1987 ; Lock 1993) (Figure 1.1.a). La formation de biofilms concerne tous les milieux aquatiques naturels (aquifères, lacs, rivières, mers) ainsi que les tissus vivants (tissus épithéliaux, dents, racines…), les biomatériaux médicaux (ustensiles médicaux, prothèses…), les dispositifs industriels et sanitaires (climatiseurs, réseaux de distribution d’eau potable, coques de navire…). Leur développement est souvent associé à de lourds enjeux sociétaux : dégradation des installations portuaires, bio-salissures des coques de navire, contamination des équipements des industries agroalimentaires et des réseaux d’eau potable, infections nosocomiales, etc. Un biofilm peut être formé par une seule espèce bactérienne (ex. modèle à
Pseudomonas aeruginosa) ou former un assemblage diversifié de microorganismes
eucaryotes unicellulaires hétérotrophes ou photosynthétiques.
Dans ce travail, l’interface liquide-solide propice à l’installation du biofilm est la pierre (du grec lithos) des galets du fond des cours d’eau (Figure 1.1.b et c). On parle alors de biofilm épilithique ou d’épilithon. Le biofilm épilithique est un assemblage très complexe reposant sur le développement de microorganismes photoautotrophes (EPU, cyanobactéries) qui lui confèrent sa coloration, et qui comprend également divers micro- et méso-organismes procaryotes et eucaryotes, autotrophes et hétérotrophes et, de façon plus ou moins transitoire (selon la période de la journée ou de l’année) des macroorganismes (larves d’Insectes aquatiques, Invertébrés benthiques…).
Figure 1.1. Observations du biofilm épilithique à l’échelle microscopique (a), du galet (b) et
du tronçon de rivière (c).
La terminologie biofilm (associée en anglais aux mots epilithic, river ou lotic) est utilisée plus largement depuis une vingtaine d’années. L’assemblage en question, défini par Wetzel (1983) comme une communauté microbienne complexe (algues, bactéries, champignons, animaux et détritus organiques et inorganiques) attachée à un substrat inorganique ou organique, vivant ou mort, est traditionnellement et majoritairement désigné sous le nom de périphyton. Son utilisation reste néanmoins très souvent associée aux communautés algales benthiques et il est principalement étudié pour son rôle de producteur primaire des milieux lotiques (par ex. Mulholland et al. 1991 ; Stevenson 1996 ; Biggs et al. 1998). D’autres qualificatifs précisent le type d’assemblage selon le type de substrat colonisé : l’épiphyton colonise les plantes aquatiques, l’épisammon colonise les grains de sable, l’épipélon colonise les sédiments fins et la boue, l’épixylon colonise le bois et l’épilithon colonise la pierre. Depuis les travaux de Lock et al. (1984), le périphyton et en particulier l’épilithon sont davantage appréhendés via leur dimension microbiologique (par ex. Romani & Sabater 2000 ; Lawrence et al. 2002 ; Battin et al. 2003a ; Olapade & Leff 2004). Enfin plus généralement, d’autres termes sont ou ont été utilisés : le terme microphytobenthos (plus utilisé en milieu marin) et le terme allemand « aufwuchs » qui signifie « se développer sur » était employé en Europe il y a une dizaine d’années (Stevenson 1996). La complexité structurelle et fonctionnelle de l’agrégat composé de microorganismes et d’EPS est associée à l’idée de biofilm, un modèle particulièrement intéressant pour appréhender les concepts écologiques de diversité, de succession écologique, de relation structure-fonction, d’interactions biologiques, etc.
Photo Samuel Teissier Photo Samuel Teissier
(a) (b) (c)
50 µm
Photo Samuel Teissier Photo Samuel Teissier
(a) (b) (c)
1.2 – Conditions de développement
La présence d’un substrat disponible et suffisamment stable pour résister au courant est un facteur nécessaire mais non suffisant pour le développement de cet agrégat. D’autres facteurs sont désignés pour contrôler le développement de l’épilithon et parmi eux, sont identifiés les ressources, les facteurs abiotiques et biotiques. Contrairement aux facteurs abiotiques et biotiques, les ressources sont directement consommées pour le métabolisme et la reproduction. Les facteurs abiotiques et biotiques influencent directement les capacités de la communauté algale à consommer ces ressources, en modifiant par exemple l’activité enzymatique ou l’intégrité physique des cellules (Stevenson 1997). Plus généralement et à l’échelle de l’hydrosystème, cet ensemble de facteurs est contrôlé par d’autres facteurs indirects tels que le climat, la géologie, l’occupation du sol, l’hydrologie, les apports en nutriments, la sédimentation et les interactions biotiques (Biggs 1995 ; Stevenson 1997).
La lumière est la première ressource indispensable : le biofilm épilithique se développe où et quand l’ombrage (contrôlé par la canopée), la hauteur d’eau et la turbidité rendent possible la pénétration de la lumière jusqu’au biofilm. Plus accessoirement, la présence de substrat vierge est une ressource nécessaire à l’installation de certaines espèces algales qui ont besoin de s’attacher directement au substrat (ex. Cocconeis ou Cladophora). Enfin, la ressource en nutriments est indispensable au métabolisme algal, et constitue d’ailleurs le facteur le plus étudié, souvent en association avec la vitesse du courant. L’assimilation des nutriments dépend de la couche limite (ou diffusive boundary layer DBL) définie comme la zone d’interface entre le biofilm épilithique et l’eau environnante lors d’un mouvement relatif entre les deux (Riber & Wetzel 1987 ; Rasmussen & Lewandowski 1998). Au sein de cette couche limite, la composante verticale du flux de transport des nutriments est fortement contrainte, la diffusion moléculaire devient le mode de transfert prédominant des composés dissous vers le biofilm (Borchardt 1996). L’existence d’un gradient de concentration entre l’eau et le biofilm est alors nécessaire à la diffusion et le taux de diffusion moléculaire est proportionnel à l’intensité de ce gradient (Borchardt 1996).
Le pH, la salinité, la température, la force de frottement liée à la vitesse du courant (shear stress), l’abrasion, ou encore la présence de composés toxiques dans l’eau constituent les facteurs abiotiques les plus importants. Leurs effets dépendent de la tolérance et des conditions optimales de développement de chaque espèce. Parmi les facteurs biotiques, on
peut citer le broutage, l’allélopathie, la lyse virale et la compétition interspécifique (Stevenson 1997).
Si la distinction entre facteurs biotiques et facteurs abiotiques est assez claire lorsque l’on considère les algues benthiques en tant que communauté, elle n’est pas aussi légitime lorsque l’on considère l’assemblage « biofilm épilithique » dans sa globalité en tant qu’unité fonctionnelle. On préférera parler du double contrôle auquel sont soumis la composition, la biomasse et le fonctionnement du biofilm : un contrôle allogène sous l’influence de facteurs qui lui sont extérieurs (vitesse du courant, nutriments, lumière, température, broutage par les poissons, etc.) et un contrôle autogène sous l’influence de facteurs qui lui sont propres (compétition, broutage, allélopathie, gradients de pH, d’oxygène, etc.).
L’ensemble des conditions favorables au biofilm épilithique en rivière co-existe dans les cours d’eau de taille intermédiaire (ordres de Strahler compris entre 3 et 7) d’après le
River Continuum Concept (RCC, Vannote et al. 1980). Les biofilms épilithiques constituent à
ce titre des assemblages largement répandus dans l’environnement aquatique. En revanche les biomasses atteintes sont très variables dans l’espace et le temps. Temporellement, Dodds et al. (1998) montrent que le rapport moyen entre biomasse maximale et biomasse moyenne obtenu dans 176 sites de l’Amérique du Nord et de la Nouvelle Zélande est de 4,52 en rivière contre seulement 1,7 – 2,6 en lacs. Ce rapport de biomasses traduit l’amplitude de la croissance annuelle du biofilm épilithique. Si cette amplitude est plus importante en rivière c’est qu’elle répond à une plus grande amplitude des conditions environnementales alternativement propices ou non au développement du biofilm. Spatialement, une revue bibliographique effectuée par Biggs & Price (1987) répertorie des biomasses maximales atteintes dans différentes rivières de Nouvelle Zélande comprises entre 0,7 et 290 g de matière sèche sans cendre (MSSC) par m2. Cette variabilité importante est largement associée à la richesse nutritive du milieu (par ex. Dodds et al. 1997 ; Biggs 2000). A ce titre, à partir de la biomasse benthique moyenne, Dodds et al. (1998) identifient trois catégories de cours d’eau selon le degré d’enrichissement en nutriments N et P (statut trophique) : le statut oligotrophe (biomasse < 20 mg de chlorophylle a par m2), le statut mésotrophe (20 < biomasse < 70 mg de chlorophylle a par m2) et le statut eutrophe (biomasse > 70 mg de chlorophylle a par m2). D’autres modèles empiriques décrivent cette relation entre la biomasse du biofilm épilithique (surtout exprimée en chlorophylle a) et la disponibilité en nutriments phosphorés principalement mais aussi azotés (par ex. Sabater & Admiraal 2005).
1.3 – Rôle fonctionnel dans les écosystèmes lotiques
Les secteurs de cours d’eau caractérisés par le développement de biofilm épilithique pourraient être qualifiés de rivière « à biomasse fixée » par analogie avec les réacteurs du même type utilisés en épuration. Ces procédés (ex. des biofiltres), caractérisés par une biomasse fixée à un matériau solide sur lequel percole l’eau à traiter, possèdent un potentiel épuratoire bien plus important que ceux à biomasse libre (ex. boues activées), en particulier dans des situations où la capacité du réacteur est limitée par la concentration en biomasse et le temps de résidence hydraulique. La biomasse fixée sur le support bactérien (lit ou disque) permet d’atteindre des temps de rétention élevés au sein du biofilm tout en opérant des temps de rétention hydrauliques relativement faibles. Les cultures fixées sont alors particulièrement utiles lorsque des temps élevés de rétention au sein de la biomasse sont exigés, ce qui est le cas des réactions qui impliquent des organismes à faible taux de croissance (ex. nitrifiants, méthanogènes). Dans les petits cours d’eau (taille intermédiaire), le temps de résidence hydraulique est faible. Les flux biogéochimiques sont alors principalement gouvernés par les processus et échanges aux interfaces hyporhéiques et benthiques (Dahm et al. 1998). En effet, à ces interfaces, les flux hydrauliques sont nuls ou quasi nuls. Les communautés biologiques peuvent donc influencer localement la concentration en nutriments par assimilation et transformation de la matière (et maintien du gradient de concentration). De plus, les biofilms développent des structures filamenteuses, des protubérances de diverses formes et tailles et un réseau complexe de canaux qui augmentent le rapport surface/volume de l’assemblage et favorisent alors le transfert de matière.
Les algues comme les bactéries sont d’importants consommateurs de nutriments inorganiques (Cole et al. 1982) : les autotrophes comme les hétérotrophes du biofilm épilithique utilisent l’azote et le phosphore de l’eau, les premiers pour construire leur propre biomasse (Bothwell 1988), les seconds lorsqu’ils dégradent la matière organique dont les rapports C/N et C/P sont élevés (Mulholland 1992). Le couplage fort entre les deux au sein du biofilm épilithique assure un recyclage plus efficace des nutriments. Leur potentiel d’élimination ou de mobilisation des nutriments a été mis en évidence en particulier concernant l’azote (Duff et al. 1984 ; Sabater et al. 2002 ; Teissier et al. 2007). Récemment les travaux de Teissier et al. (2007) montrent que l’élimination de l’azote s’effectue en deux étapes au cours de la maturation du biofilm : un biofilm jeune et fin assimile l’azote en provenance de la colonne d’eau et le stocke (croissance), les processus de minéralisation,
nitrification et dénitrification sont faibles ; un biofilm épais libère l’azote préalablement stocké sous la forme de N2 grâce au couplage de réactions minéralisation – nitrification –
dénitrification. Soit le biofilm s’autominéralise et donc, sa biomasse diminue, soit l’assimilation d’azote organique en provenance de la colonne d’eau compense la perte de biomasse.
Les biofilms épilithiques sont aussi des sites efficaces pour l’assimilation, l’immobilisation (le stockage) et la transformation des substances organiques particulaires et dissoutes. La croissance du biofilm favorise le dépôt de particules organiques en rivière (Thomas et al. 2001 ; Battin et al. 2003a), grâce à ses propriétés d’adhésion et de filtration. Les cellules attachées tirent profit du piégeage de carbone exogène et d’énergie. La proximité des organismes autotrophes et hétérotrophes au sein de l’agrégat ainsi que la capacité de dégradation extracellulaire au sein de la matrice d’EPS (Sinsabaugh et al. 1991) permettent un recyclage efficace du carbone apporté par la photosynthèse au sein de la communauté (Lock et al. 1984). Les organismes au sein du biofilm sont en interaction constante : l’agrégat est assimilé à une plateforme d’échanges de matériel génétique et d’interactions métaboliques (Stoodley et al. 2002). Ces échanges et interactions sont limités pour les cellules planctoniques puisque ces dernières ne se trouvent pas en contact physique. L’activité métabolique des communautés en biofilm dépasse donc largement l’activité planctonique (Geesey et al. 1978).
Enfin, dans ces cours d’eau, de par sa fonction de producteur primaire, le biofilm épilithique est à la base du réseau trophique. Dodds (2006) démontre effectivement que la biomasse algale benthique est corrélée positivement à la production primaire brute des rivières. Or cette production primaire gouverne le maintien de la structure et du fonctionnement des communautés d’une rivière d’après (Minshall 1978). McIntire (1973) prouve par modélisation que, grâce à un turnover rapide, le périphyton, même en faible quantité, est capable de soutenir d’importantes biomasses de consommateurs.
1.4 – Dynamique temporelle de la biomasse
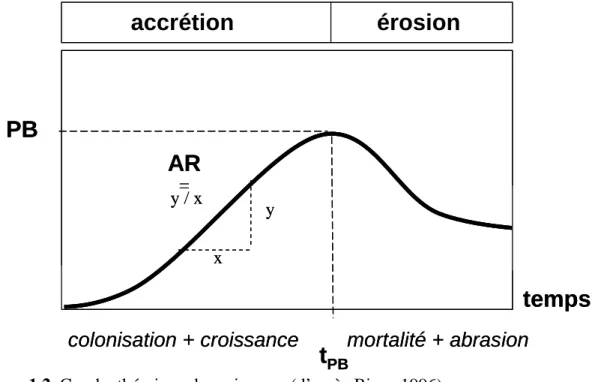
La dynamique temporelle à court terme de la biomasse épilithique résulte de l’équilibre entre des processus d’accrétion (importation et prolifération de cellules) et de perte (mort et/ou émigration de cellules). L’état des connaissances relatives à cette dynamique a été conceptualisé par les travaux de Biggs (1996) qui la décrit par une courbe théorique en deux
phases (Figure 1.2.). La phase de croissance qui présente une évolution exponentielle de la biomasse est successivement dominée par les processus de colonisation et de croissance. Lors de la phase de perte, ces derniers sont largement compensés par des pertes liées à la sénescence, au parasitisme, au broutage ou au détachement autogène (autogenic sloughing). Enfin la biomasse atteint un palier égal à la capacité de charge de l’assemblage (carrying
capacity). La forme de cette courbe universelle est fonction des paramètres tPB, le temps
nécessaire pour atteindre le pic de biomasse, et PB, la biomasse maximale atteinte, paramètres pouvant être synthétisés par la vitesse d’accrétion (AR).
Figure 1.2. Courbe théorique de croissance (d’après Biggs 1996).
En milieu naturel cette évolution théorique est très rarement observée, le processus de croissance étant le plus souvent interrompu par une perturbation hydrodynamique (vitesses du courant élevée et/ou substrat instable et/ou abrasion) à l’origine d’une perte brutale de biomasse. L’évènement ne conduit pas nécessairement à la disparition complète du biofilm. L’amplitude, la durée et/ou la fréquence de cet évènement ainsi que les propriétés morphotypiques, taxonomiques et physiologiques (histoire et âge) conditionnent la réponse structurelle et fonctionnelle (résistance et résilience) du biofilm à la perturbation (Peterson & Stevenson 1992). A long terme aussi le régime de perturbation hydrologique est le principal facteur de structuration temporelle de la biomasse. Biggs (1996) distingue trois patrons de dynamique temporelle de la biomasse à long terme :
PB
t
PBcolonisation + croissance
mortalité + abrasion
y x
AR
= y / xaccrétion
érosion
temps
PB
t
PBcolonisation + croissance
mortalité + abrasion
y x
AR
= y / xaccrétion
érosion
temps
• une dynamique stable et caractérisée par de faibles biomasses est associée le plus souvent à des perturbations hydrologiques fréquentes et/ou une forte instabilité du substrat,
• une dynamique rythmée par des cycles saisonniers associés à une importante saisonnalité du régime hydrologique, de l’activité des brouteurs ou de la lumière,
• une dynamique marquée par la succession de cycles d’accrétion/détachement est souvent caractéristique de rivières où les perturbations hydrologiques sont saisonnières ou peu fréquentes.
L’épaississement du biofilm qui se produit lors des longues phases de stabilité qui caractérisent les deux derniers types d’évolution temporelle s’accompagne d’une modification de la composition (biodiversité), de l’architecture (microgradients et distribution tridimensionnelle des organismes) et du fonctionnement (métabolisme) de l’assemblage. Les formes algales colonisatrices (ou pionnières), généralement de petite taille et au développement rapide, sont progressivement remplacées par des formes algales climaciques, généralement de grande taille et au développement lent (Biggs et al. 1998). Le morphotype (unicellulaire, colonial ou filamenteux) et le mode de fixation (prostrée ou érigée) des algues présentes conditionnent la structure tridimensionnelle de l’assemblage (Stevenson 1996) : l’assemblage rendu très cohésif par la présence de petites diatomées capables d’adhérer très fortement au substrat et de diatomées coloniales fixées à l’apex devient plus aéré et volumineux avec l’installation des algues vertes filamenteuses.
Le concept de succession écologique (Odum 1956) a été généralisé aux communautés bactériennes de biofilms de rivière par Lyautey et al. (2005a). Les OTUs (operational
taxonomic units) identifiées dans cette étude au cours d’une phase d’accrétion indiquent des
profils de présence et des exigences écologiques différents au cours du temps qui pourraient s’apparenter à des stratégies écologiques (Lyautey et al. 2005a). En début de croissance, alors que le biofilm est fin, les populations détectées correspondraient à des taxons aérobies stricts (Spirosoma sp.). En fin de croissance, alors que les couches profondes se désoxygènent, les populations identifiées présenteraient vis-à-vis de l’oxygène une adaptation à de faibles concentrations (aérobie facultative Dechloromonas sp., microaerophile Nitrospira sp.).
Enfin l’épaississement du biofilm modifie physiquement la nature des échanges avec la colonne d’eau. Lorsque le biofilm est fin, le transport actif est prépondérant par rapport à la diffusion parce que la concentration en nutriments au-dessus du biofilm reste importante. Lorsque le biofilm est épais, le flux diffusif au sein de la couche limite devient limitant car il est plus faible que le taux potentiel de consommation des nutriments par les cellules
(Borchardt 1996). Avec la maturation du biofilm, l’activité métabolique devient donc davantage fonction de l’épaisseur du biofilm que des conditions extérieures.
Finalement, en absence de perturbation, si les premières semaines de la vie du biofilm sont largement conditionnées par les ressources nutritives et lumineuses, la température et la vitesse du courant, il semblerait qu’au fur et à mesure de la maturation de l’agrégat, l’augmentation des interactions biotiques entre les différents organismes et l’isolement physique associé à l’épaississement tendent à limiter les interactions avec la colonne d’eau environnante et donc l’importance du contrôle allogène.
1.5 – Intérêt pour la bioindication
Les milieux aquatiques subissent depuis plusieurs décennies une pression croissante des activités humaines qui affectent leur intégrité et leur santé écologique. La prise de conscience de la nécessité de ralentir, stopper voire inverser cette tendance a présidé à la mise en place de la directive cadre européenne sur l’eau (2000/60/CE) qui impose d’atteindre le « bon état écologique » d’ici 2015. La nécessité sous-jacente de caractériser l’état des milieux et d’évaluer les efforts de réhabilitation suppose de disposer de critères de diagnostic fiables et adaptés.
De par ses caractéristiques structurelles et fonctionnelles, le biofilm épilithique possède un fort potentiel de bioindication. En particulier, les potentialités des microalgues comme descripteurs structurels de l’état du milieu ont été déjà largement explorées. Parmi les avantages de ces organismes leur temps de génération assez faible leur permet de répondre à des changements de qualité de l’eau plus rapidement que les macrophytes ou la faune aquatique. La réponse des microalgues est souvent symptomatiquement associée à des changements de nutriment. Par exemple un rapport N/P de l’eau faible et/ou des variations en P ou N total peuvent contribuer au développement de blooms cyanobactériens (Downing et al. 2001). Un faible ajout de P dans des systèmes à fort rapport N/P peut être suffisant pour faire basculer la communauté algale d’une dominance d’espèces limitées par le phosphore (ex. diatomées, algues vertes) à une dominance d’espèces limitées par l’azote, parmi lesquelles les cyanobactéries peuvent jouer le rôle principal. Ces changements affectent la composition des producteurs primaires et par conséquent le fonctionnement de l’écosystème tout entier. Les indices diatomiques ont été mis au point pour synthétiser l’information issue des préférences autoécologiques de la composition taxonomique de la communauté diatomique. Ces indices
renseignent sur la capacité des communautés diatomiques à détecter les variations du pH de l’eau (Van Dam et al. 1994), de la salinité (Veres et al. 1995), du niveau nutritif (Lecointe et al. 1993) et du phosphore total (Poulickova et al. 2004) et sont, pour la plupart, inspirés de l’index de saprobie de Kolkwitz & Marsson (1909).
Les propriétés fonctionnelles des microorganismes photosynthétiques et de la composante hétérotrophe du biofilm épilithique commencent à être étudiées avec l’utilisation du métabolisme comme descripteur fonctionnel. L’équilibre entre la production et la consommation de carbone organique renseigne sur l’importance relative des deux sources d’énergies majeures, les algues (matière autochtone) ou la matière organique terrestre (allochtone). Si la production de carbone organique égale ou excède sa consommation dans l’écosystème la matière organique produite pourra supporter la chaîne trophique. En revanche si la consommation de carbone excède largement sa production la matière organique issue du bassin versant sera nécessaire pour maintenir le système. Le métabolisme permet une mesure directe de la base de la chaîne trophique et une détermination de la capacité de l’écosystème à maintenir la vie. L’utilisation de la production microphytobenthique pour le monitoring est d’autant plus intéressante qu’elle est le résultat d’une conception qui considère le biofilm comme unité fonctionnelle (Lock 1993).
Le biofilm est un agrégat susceptible d’accumuler des toxiques organosolubles et d’en être protégé. Son développement intègre dans la durée de grandes quantités d’eau ainsi que les variations spatio-temporelles de leur qualité. Moins sensible à un pic de contamination qu’à un niveau moyen, le biofilm peut avoir un rôle intégrateur de bioaccumulateur. Les effets à long terme des herbicides (atrazine) et des métaux lourds (cuivre) sur la biomasse chlorophyllienne ont par exemple été observés sur des communautés naturelles (Navarro et al. 2002). C’est aussi un modèle écotoxicologique paradoxal puisque les organismes sont à la fois protégés mais aussi plus intensément exposés à un contaminant qui s’accumulerait dans les EPS.
Le faible temps de génération des organismes qui le composent, ses propriétés bioaccumulatrices, sa complexité structurelle (différents groupes taxonomiques) et fonctionnelle (autotrophie/hétérotrophie) confèrent à cet assemblage des propriétés bioindicatrices indéniables. L’importante gamme de descripteurs structurels (biomasse, composition taxonomique, composition chimique) et fonctionnels (production, respiration, activité enzymatique extracellulaire) qui lui sont associés fait du biofilm un candidat idéal à l’indication de perturbation des écosystèmes lotiques (Burns & Ryder 2001).
2 – Les modèles mathématiques : de la description à la prédiction
2.1 – Définitions
Un modèle est une représentation simplifiée d’un processus ou d’un système en vue de le décrire (de l’expliquer) ou de le prévoir. Deux finalités sont attribués aux modèles : le modèle descriptif et le modèle prédictif. Le modèle descriptif est utilisé pour interpréter une masse d’informations réelles (ex. modèle de croissance exponentielle des microorganismes). Le modèle prédictif est utilisé pour anticiper des événements réels (ex. modèle météorologique). Cette classification reste néanmoins très arbitraire dans la mesure où ces modèles sont très étroitement liés. Une prédiction satisfaisante de la réalité future nécessite préalablement une bonne compréhension et description de la réalité passée ou actuelle. Inversement une bonne prédiction doit servir de diagnostic pour identifier des perspectives futures.
La description d’un système passe généralement par l’association de grandeurs (ou fonction). La plupart des outils de l’analyse mathématique sont particulièrement bien adaptés à l’étude des fonctions réelles d’une ou plusieurs variables. La mise en œuvre d’un modèle suppose une traduction des propriétés physiques, biologiques et/ou chimiques du système en des termes mathématiques. Le langage mathématique représente de manière symbolique et logique des idées et des relations (plus ou moins complexes). Il donne ainsi accès à une description quantitative de l’état du système et des processus qui s’y déroulent.
Trois types de modèles mathématiques sont traditionnellement considérés : le modèle statistique, le modèle mécaniste stochastique (ou probabiliste ou évènementiel) et le modèle mécaniste déterministe. Le modèle statistique permet d’identifier des liens entre les éléments d’un système au moyen d’études de corrélations et d’analyses de variance. Il a avant tout un fort potentiel de description. Il est parfois utilisé comme outil de prédiction mais sa représentation du système est instantanée et représentative du système au seul moment de la prise de mesure. Le modèle (mécaniste) déterministe est basé sur une ou des équation(s) différentielle(s) et ses variables sont des grandeurs précises. L’évolution temporelle de la (ou des) variable(s) est entièrement déterminée par les conditions initiales, les paramètres et les variables externes. La mise en œuvre d’un modèle déterministe est guidée par la théorie et s’appuie sur des équations phénoménologiques ou sur des schémas de fonctionnement (modèle conceptuel). Elle suppose donc de connaître au préalable et au moins en partie les
mécanismes impliqués. Le modèle (mécaniste) stochastique est régi par des lois/distributions de probabilité. Sa mise en œuvre est guidée par les données ce qui suppose de pouvoir observer et analyser les phénomènes. Finalement pour des conditions initiales identiques, le modèle déterministe donnera toujours le même résultat alors que le modèle stochastique produira des résultats différents selon la valeur actuelle des variables prises au hasard. Le développement et l’utilisation d’un modèle mathématique exige, quel que soit le modèle considéré, de disposer en parallèle d’observations adaptées et suffisantes aux échelles spatiales et temporelles requises.
2.2 – Intérêts pour l’écologie
A l’image des expérimentations in vivo, in vitro ou in situ très classiquement utilisées en biologie depuis le XIXième siècle, l’expression in silico désigne une expérimentation entièrement réalisée par ordinateur. Contrairement aux autres qualificatifs, tous issus du latin, cette expression tire son nom de la puce au silicium, composant de base du microprocesseur ayant donné par ailleurs son nom à l’industrie des hautes technologies qu’est la Silicon Valley. L’expérimentation in silico (ou expérimentation numérique) est née grâce à la puissance de calcul des outils informatiques capables de faire tourner des modèles mathématiques en relation avec la biochimie, le comportement et l’environnement animal. Elle permet comme les autres expérimentations de tester une ou plusieurs hypothèses scientifiques mais, dans certains cas, elle est la seule alternative pour représenter et tester des hypothèses complexes. En effet un écosystème est constitué de tant de composantes en interactions qu’il est difficile de les examiner toutes. Lorsque cela est possible, l’interaction examinée isolément en laboratoire est nécessairement différente de celle qui se produit naturellement en présence d’autres interactions. C’est ce qu’illustre la phrase qui résume bien le principe de l’émergence : “tout est plus grand que la somme des parties » (Allen 1988). La complexité d’un système dépend du nombre d’interactions entre ses diverses composantes mais aussi du nombre de feedbacks et de régulations qui permettent au système de se maintenir malgré l’apparition de conditions défavorables. Par exemple, les cellules algales sont capables de réguler leur contenu en chlorophylle a selon l’intensité du rayonnement solaire. Si elles s’avèrent insuffisantes, ces interactions, ces feedbacks et ces régulations évoluent au cours du temps pour une meilleure utilisation des ressources disponibles. Le changement saisonnier par exemple peut conduire à un remplacement d’espèces algales. Cela
implique donc évidemment de prendre en compte l’histoire de l’écosystème et de ses composantes (Patten 1997) et justifie la nécessité de travailler en dynamique. Le modèle dynamique (ou permanent), qui se distingue du modèle statique (ou transitoire), et prédit des variables qui changent au cours du temps est donc préféré en écologie.
2.3 – Démarche de modélisation
La démarche de modélisation est fondée sur le principe selon lequel tout est système ou tout peut être conceptualisé selon une logique de système, principe appelé théorie des systèmes ou théorie systémique, formalisée notamment en 1968 par Ludwig von Bertalanffy. Du grec sustêma signifiant ensemble, le système fait référence à un assemblage d’éléments en interaction permanente et fonctionnant de manière unitaire. Le modèle s’intéresse à un système qu’il considère comme un ensemble fini d’éléments appelés variables d’état, qui comme leur nom l’indique, décrivent son état et, d’interactions (processus) entre ces variables qui permettent de décrire son évolution dans le temps et dans l’espace. Ces processus biologiques, chimiques ou physiques sont représentés par des équations mathématiques. Ces équations sont décrites par un nombre plus ou moins important de coefficients (paramètres ou constantes universelles). Les conditions qui ne sont pas décrites par le modèle lui sont imposées par le biais de fonctions dites de forçage qui peuvent fluctuer dans l’espace et dans le temps et qui sont généralement obtenues à partir de données mesurées.
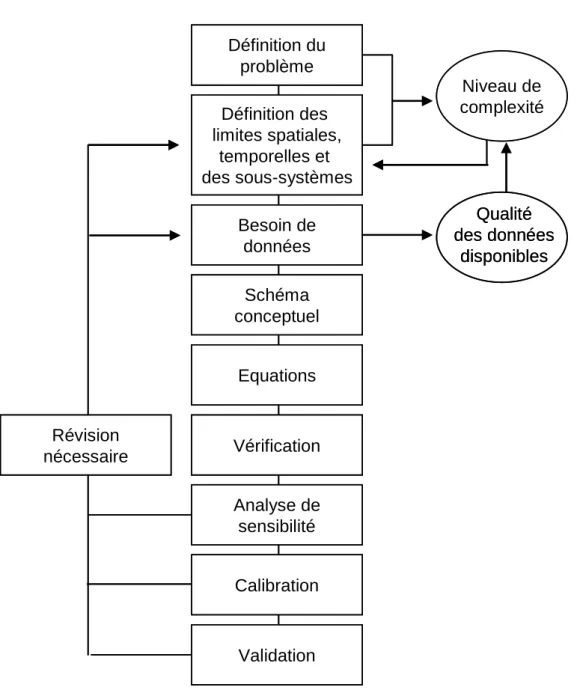
La mise en œuvre d’un modèle nécessite au préalable de définir le problème (Figure
1.3.). Elle exige ensuite de définir une échelle de temps et d’espace adaptée au problème à
résoudre sachant qu’au niveau de perception (spatio-temporel), les éléments du niveau inférieur liés par des interactions intenses et fréquentes ne sont pas décrits mais agrégés et donc mis sous silence. Idéalement l’acquisition de données s’effectue avant même la mise en équation de manière à adapter la qualité des données au niveau de complexité du modèle exigé par l’objectif. L’identification des processus dominants aboutit à l’écriture d’un schéma conceptuel. Elle s’effectue à partir des données du suivi expérimental et dépend donc nécessairement des conditions particulières ayant prévalu pendant la période de ce suivi. A l’issue de cette étape le modèle est formalisé mathématiquement (écriture des équations) et programmé. L’étape de vérification permet ensuite de vérifier si le comportement du modèle est en accord avec ce que l’on connaît du système. L’analyse de sensibilité examine la réponse du modèle à la variation des paramètres et permet ainsi de discriminer les paramètres
du modèle les plus sensibles. La qualité du modèle est ensuite testée lors de l’étape de calibration en comparant le résultat du modèle (la simulation) avec le système réel observé que le modèle est sensé représenter.
Figure 1.3. Représentation schématique de la démarche de modélisation (d’après Jorgensen
1983).
L’étape de calibration consiste à choisir à l’intérieur d’une gamme pré-établie, les valeurs de paramètres qui donnent le meilleur ajustement avec les données observées. Plusieurs cas de figures existent : les paramètres sont mesurés sur le terrain ou en laboratoire ; ils proviennent
Définition du problème Définition des limites spatiales, temporelles et des sous-systèmes Besoin de données Schéma conceptuel Equations Vérification Qualité des données disponibles Révision nécessaire Analyse de sensibilité Calibration Validation Niveau de complexité Définition du problème Définition des limites spatiales, temporelles et des sous-systèmes Besoin de données Schéma conceptuel Equations Vérification Qualité des données disponibles Révision nécessaire Analyse de sensibilité Calibration Validation Niveau de complexité
de la littérature ; ils sont ajustés car non mesurables et/ou n’ayant pas de signification connue. Si la calibration ne permet pas une bonne adéquation entre les observations et simulations, le modèle doit être révisé. Cela peut aussi conduire à la réalisation d’expériences destinées à améliorer la description des processus. Si l’objectif fixé est de générer de nouvelles hypothèses et de synthétiser les connaissances issues d’études expérimentales, ces étapes sont suffisantes. Si l’objectif est prédictif, l’étape de validation est indispensable pour s’assurer que le comportement du modèle est représentatif du système même lors de périodes différentes de la période de calibration. S’en suit alors l’importance de disposer d’un éventail large de situations permettant de s’affranchir de conditions particulières.
Calibration et sélection de modèle
Revenons sur l’étape de calibration. La méthode de confrontation la plus simple entre le modèle et les données est celle qui consiste à minimiser les moindres carrés. Dans certains cas plusieurs modèles de complexité différente peuvent être proposés. La somme des carrés des écarts la plus faible est obtenue avec le modèle comprenant le plus de paramètres mais l’ajout de paramètres augmente les difficultés d’interprétation. Les méthodes de sélection de modèles ont été conçues pour pouvoir comparer un modèle à n paramètres d’avec un modèle à
p paramètres et déterminer lequel d’entre eux est le meilleur vis-à-vis des données. Plus
fondamentalement la sélection de modèles vise à sélectionner l’hypothèse la plus probable (la plus supportée par l’observation) parmi plusieurs hypothèses formulées. Parmi les critères qui permettent de classer les différents modèles et d’évaluer la contribution relative de chacun, deux critères sont utilisés en écologie : le critère d’information d’Akaike (AIC) et le critère de Schwarz (SC), connu aussi sous le nom de critère d’information bayésien (BIC). Ces derniers utilisent le maximum de vraisemblance comme mesure de l’ajustement et pénalisent l’introduction de paramètres supplémentaires. Pour l’instant leur application reste exclusivement réservée aux domaines des analyses de marquage-recapture pour estimer l’abondance de la population et les probabilités de survie (par ex. Lebreton et al. 1992) et de l’évolution pour la reconstruction phylogénétique (par ex. Posada & Crandall 2001).
2.4 – Limites et contraintes d’application
Les premiers modèles mathématiques en écologie sont nés avec les travaux de Lotka-Volterra et de Streeter-Phelps dans les années 1920. L’utilisation des modèles pour la gestion de l’environnement s’est largement répandue depuis 1970. La prise de conscience des problèmes théoriques/fondamentaux associés à la mise en œuvre de la modélisation mathématique en écologie est plus récente. L’estimation des paramètres, l’insuffisance des données et l’incapacité des modèles à représenter le potentiel d’adaptation des organismes et des écosystèmes (contrairement aux systèmes physiques qui sont stables) figurent parmi les points les plus problématiques (Jorgensen 1999). La question majeure éternellement associée à la construction d’un modèle est : quel niveau de complexité le modèle doit-il avoir sachant qu’un modèle trop simple risque d’écarter des processus significatifs du système et qu’un modèle trop complexe risque de rendre impossible la calibration, faute de données suffisantes, et de rendre difficile la compréhension du système?
Contrairement à l’écosystème qui est capable de réguler, modifier et changer ses paramètres en réponse à des changements environnementaux, la structure et le nombre de paramètres du modèle sont fixés. Le modèle se base sur l’analyse du système à l’instant t pour prédire la réponse du système à l’instant t+1 sans tenir compte de l’adaptation des processus au nouveau système. Cela explique pourquoi les paramètres du modèle peuvent représenter correctement la variable d’état telle qu’elle est dans l’écosystème pendant la période étudiée et ne pas être nécessairement valides pour une autre période de temps.
Ce problème constant a conduit certains auteurs à introduire la notion d’incertitude dans la construction des modèles. Hilborn & Mangel (1997) distinguent deux formes d’incertitude : l’incertitude (ou bruit, ou stochasticité ou erreur) de processus i.e. associée à la mise en oeuvre du modèle (process uncertainty) et l’incertitude d’observation (observation
uncertainty). Le fait que les paramètres puissent varier d’une manière imprévue, par exemple
qu’un taux de croissance fluctue d’une année à l’autre représente une erreur de processus. L’erreur associée à l’échantillonnage (par ex. liée à la taille de l’échantillon) est une erreur d’observation. Contrairement à l’incertitude de processus, l’incertitude d’observation ne se propage pas au cours du temps. Cette notion d’incertitude est particulièrement importante dans les modèles écologiques : la forte incertitude d’observation existe chez les communautés épilithiques comme dans la plupart des séries temporelles écologiques : leur dynamique temporelle est fortement contrôlée par des processus stochastiques (potentiel de colonisation
rapide, fort renouvellement d’espèces, etc.) ; leurs biomasse et composition sont fortement conditionnées par la variabilité spatiale des variables environnementales locales comme la vitesse, la granulométrie, la stabilité du substrat, le broutage, etc.
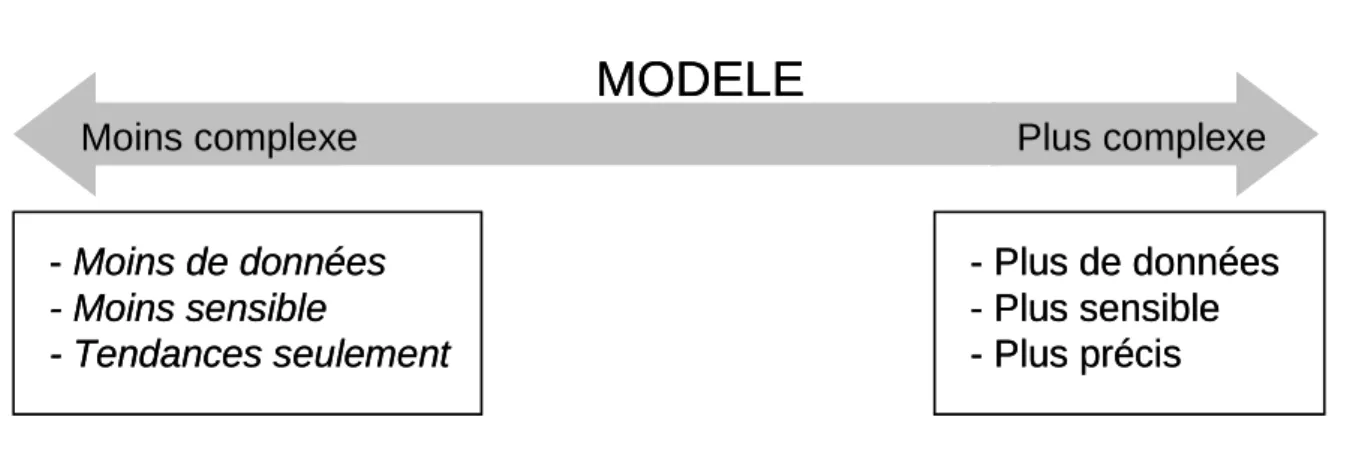
L’incertitude est indissociable des notions de complexité, de flexibilité, de sensibilité d’un modèle (Figure 1.4.).
Figure 1.4. Spectre de la complexité d’un modèle (adapté de Snowling & Kramer 2001). La
flèche représente la gamme de complexité possible pour un modèle, de plus en plus simple (à gauche) et de plus en plus complexe (à droite).
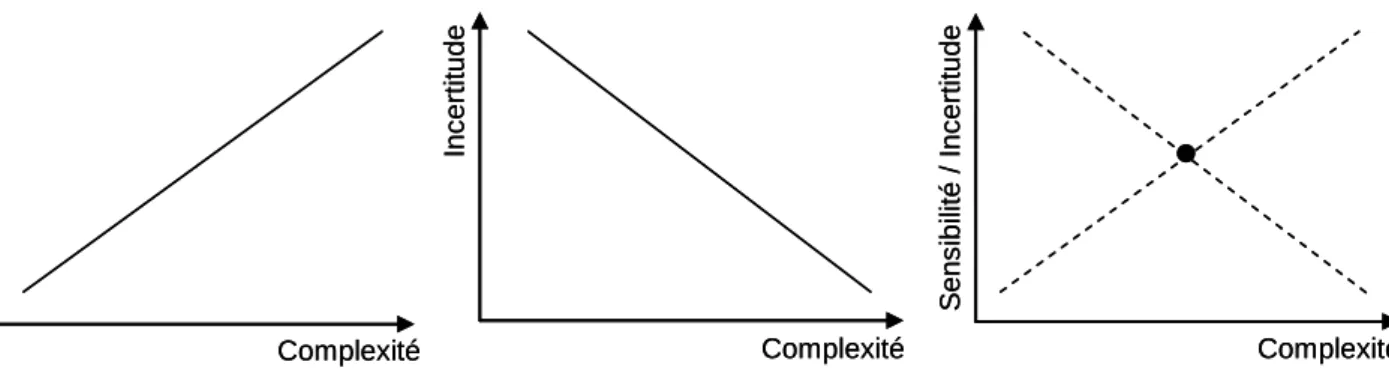
La sensibilité d’un modèle (model sensitivity) ou la performance d’un modèle correspond à sa capacité à décrire l’évolution de sa ou ses variable(s) d’état. Plus un modèle est complexe plus il est sensible, en raison du plus grand degré de liberté et de la structure des interactions entre paramètres et variables d’état. Et plus un modèle est complexe et plus il intègre de processus, moins il assume de simplifications, mieux il simule la réalité et donc moins l’incertitude est élevée. Quant à la flexibilité, elle se réfère au nombre d’hypothèses effectuées lors du développement du modèle. Moins le modèle est complexe, plus il assume d’hypothèses pour réduire le nombre de variables d’état et de paramètres et donc moins il est flexible. Les hypothèses restreignent le modèle aux situations pour lesquelles elles sont valides alors que plus le modèle est complexe et plus il couvre de situations.
Néanmoins la complexification d’un modèle rend l’identification de ses paramètres plus insignifiante, en raison notamment des corrélations croissantes entre paramètres, et requiert davantage de données. Pour Zucchini (2000), il existe un niveau optimal de complexité pour les modèles prédictifs. Il distingue une erreur de prédiction liée à l’approximation, qui diminue lorsque la complexité du modèle augmente, d’une erreur de
Moins complexe Plus complexe
- Moins de données - Moins sensible - Tendances seulement - Plus de données - Plus sensible - Plus précis
MODELE
Moins complexe Plus complexe
- Moins de données - Moins sensible - Tendances seulement - Plus de données - Plus sensible - Plus précis
MODELE
prédiction liée à l’estimation, qui augmente avec la complexité. Quel que soit le modèle et le jeu de données, l’erreur de prédiction diminue puis augmente avec une complexité croissante, justifiant de l’existence d’un niveau optimal intermédiaire de complexité. C’est aussi ce que démontrent Snowling & Kramer (2001) en comparant deux modèles de complexité différente (Figure 1.5.). Plusieurs auteurs s’accordent à dire que le niveau de complexité d’un modèle dépend du modèle mis en œuvre et de l’objectif recherché (par ex. Hilborn & Mangel 1997 ; Snowling & Kramer 2001) et que les modèles de « recherche » peuvent être plus complexes que les modèles de « gestion ».
Figure 1.5. Schéma illustrant la relation entre complexité – incertitude – sensibilité d’un
modèle. Plus la complexité d’un modèle augmente, plus sa sensibilité augmente et plus son incertitude diminue. Il existe un modèle de complexité intermédiaire qui optimise la relation complexité – incertitude – sensibilité (d’après Snowling & Kramer 2001).
3 – Objectifs du travail
3.1 – Problématique
Les communautés microbiennes du biofilm épilithique sont fortement impliquées dans les processus de production, de recyclage, de dégradation ou de stockage de la matière au sein des écosystèmes aquatiques. Leur développement est sous le contrôle d’un ensemble de facteurs allogènes qui coexistent largement dans les écosystèmes lotiques, en particulier dans les cours d’eau d’ordre intermédiaire (entre 3 et 7) tels que définis par le River Continuum Concept. Dans ces milieux le biofilm épilithique constitue la base du réseau trophique et son rôle dans le fonctionnement biogéochimique est largement reconnu (Battin et al. 2003b). Le fonctionnement (voire le dysfonctionnement) biogéochimique des écosystèmes colonisés est directement relié à la biomasse fixée et son évolution au cours du temps. Si les premières étapes de l’accrétion du biofilm (court terme) ont été largement étudiées, peu de travaux
Sen s ib ilit é In c e rti tude Se ns ib ilit é / In c e rt it u d e
Complexité Complexité Complexité
Sen s ib ilit é In c e rti tude Se ns ib ilit é / In c e rt it u d e