HAL Id: hal-02811273
https://hal.inrae.fr/hal-02811273
Submitted on 6 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Comportement et performances du pois protéagineux en
système agroforestier. Approche agronomique et
écophysiologique de l’association pois et arbre en
agroforesterie
Marie Raimbault
To cite this version:
Marie Raimbault. Comportement et performances du pois protéagineux en système agroforestier. Approche agronomique et écophysiologique de l’association pois et arbre en agroforesterie. [Stage] Autres régions du monde. Université de Rennes 1 (UR1), FRA. 2013, pp.36. �hal-02811273�
Master STS 1
èreannée, Mention Biologie, Agronomie, Santé
Sciences & productions végétales (M1 SPV)
Année universitaire 2012-2013
Marie RAIMBAULT
Comportement et performances du pois protéagineux en
système agroforestier
Approche agronomique et écophysiologique de l’association pois et arbre en agroforesterie
Préparé à : UMR System,
SupAgro, 2 Place Viala, Bât. 27 34060 Montpellier Cedex 1 – France Maître de stage : Lydie DUFOUR, Assistante ingénieur, UMR System Membres du jury : Safya MENASSERI, Matthieu CAROF
Sommaire
Introduction
…...p. 1Matériels et Méthodes
1) Site …...…...…...…...…...…...…...…...…...…...p. 4 2) Matériel Biologique Les arbres …...p. 4 La culture …...p. 5 3) Méthodes Dispositf …...p. 6 Variables biologiques étudiées …...p. 6 Variables environnementales …...p. 7 Analyse statistique …...p. 8Résultats
1) Influence de l'arbre sur l'environnement lumineux de la culture
Mesure du rayonnement global le 13/04/13 …...p. 9 Mesure du rayonnement global le 10/06/13 …...p. 9 Capteurs PAR …...p. 9
2) Impact sur la levée …...p. 10 3) Mesure de la biomasse aérienne au stade floraison …...p. 11 4) Les nodules au stade floraison …...p. 11 5) Estimation de l'influence des arbres sur le rendement ...p. 12 6) Mesure de la biomasse aérienne au stade remplissage des gousses...p. 13
Discussion
…...p. 15Conclusion
…...p. 17Bibliographie
…...p. 18Annexes
…...p. 19Introduction
L'agriculture de nos jours doit répondre à des enjeux d'ordre écologique et économique, souvent antagonistes et difficiles à concilier. On assiste en effet à la fois à une augmentation de la demande en denrées alimentaires dans un contexte d'accroissement de la population humaine, et à une pression de plus en plus forte vis à vis de l'impact de la production sur l'environnement. La limitation des ressources naturelles, le changement climatique et l'érosion de la biodiversité sont des éléments qui justifient la nécessité de faire évoluer nos pratiques. L'agroforesterie est un mode de production répondant à ces contraintes, visant à optimiser l'espace en intégrant les arbres et les cultures dans un même système, et à le dynamiser au maximum par le jeu des associations afin de le rendre plus productif. C'est autour de ce concept d' « intensification biologique » que s'articulent les recherches de l'Unité Mixte de Recherche SYSTEM - Fonctionnement et conduite des systèmes de culture tropicaux et méditerranéens, sous la tutelle des organismes Cirad, Inra, et SupAgro.
L'agroforesterie est un système de production ancestral, dont plusieurs exemples de cultures traditionnelles régionales, souvent en association avec du pastoralisme, restent en mémoire : vergers normands ; bocage breton ou encore oulières en zone viticole (Dupraz & Liagre, 2008). Celle-ci a été fortement négligée pendant la Révolution Verte, au profit des monocultures plus adaptées au développement de la mécanisation. Cette tendance s'est encore plus accentuée avec la mise en place de la Politique Agricole Commune en Europe puisque celle-ci rendait plus rentable une gestion par assolement des terres plutôt que par des associations de cultures. Néanmoins, on assiste aujourd'hui à un regain d'intérêt pour cette pratique, qui semble pouvoir répondre aux enjeux économiques, écologiques tout en assurant un niveau de productivité suffisant. De nouvelles caractéristiques distinguent l'agroforesterie moderne de sa forme traditionnelle : on privilégiera plutôt des lignes d'arbres à long tronc, petite couronne pour limiter l'ombrage sur les cultures, et à croissance rapide, visant la production de bois d’œuvre, tandis que le système traditionnel se présentait comme une dispersion plus ou moins aléatoire de petits arbres fruitiers (Nerlich et.al., 2013). Les systèmes sylvo-arables modernes présentent de ce fait une structure appropriée pour le recours à la mécanisation, les rangées d'arbres permettant le passage des engins agricoles, avec pour objectif ultime de rendre la partie culturale exploitable jusqu'à la récolte des arbres.
Un des objectifs visés dans un système pluri-strates est un meilleur partage des ressources, telles que l'eau et les nutriments. Ceci s'expliquerait par une meilleure utilisation de l'espace souterrain, c'est-à-dire par la séparation spatiale des systèmes racinaires des plantes ligneuses et
herbacées. Le recyclage des éléments lessivés, un apport de matière organique par la litière et le turn over racinaire des arbres et une meilleure circulation de l'eau dans le système sont ainsi considérés comme des services possibles issus de cette méthode de production. Cette meilleure capture des ressources au profit de différentes espèces dans un même système, permettrait d'augmenter la biomasse totale produite par unité de surface par rapport à la biomasse cumulée dans des systèmes séparés. C'est la notion de surface équivalente assolée, définie comme « la surface nécessaire pour obtenir avec un assolement agriculture-forêt la même production qu'un hectare de système agroforestier » (Dupraz & Liagre, 2008). De plus, la présence des arbres agirait comme un « effet tampon », stabilisant le sol et permettant ainsi une meilleure résilience des systèmes et donc une stabilité des rendements accrue.
Des exemples de compétition sont cependant cités dans la littérature scientifique, mettant en exergue les points faibles de la pratique : compétition pour l'eau (Jose et.al., 2000), pour la lumière, pour les nutriments du sol. La diminution de lumière peut jouer sur la phénologie de la culture (McGraw at.al.,2008), qui voit sa maturité retardée en système agroforestier. Mais certaines solutions sont envisageables afin de limiter les phénomènes de compétition qui seraient préjudiciables. Un choix judicieux des espèces dans le système et un élagage correct de la ligne d’arbres, sont indispensables, tout en prenant en compte les caractéristiques du sol et du climat de la région (Nerlich et.al., 2013). On peut par exemple atténuer l'effet négatif du manque de lumière en choisissant d'implanter des arbres à débourrement tardif, ou à couronne perméable à la lumière, en privilégiant un axe orienté Nord-Sud, afin de limiter l'hétérogénéité au sein de la parcelle.
La problématique de l'azote s’insère aussi en premier plan dans les services attendus de l'agroforesterie. De nombreuses études portent sur ce domaine, visant à intégrer des légumineuses dans le système pour remplacer les apports d'intrants. Le recours à des arbres fixateurs d'azote, comme les acacias, est assez répandu dans les zones tropicales et subtropicales. Mais ces espèces ligneuses fixatrices sont peu présentes en Europe, l'enjeu ici est donc d'intégrer des légumineuses sous forme de plantes annuelles dans un système agroforestier. L'UMR System s'est déjà penché sur la question lors d'une étude portant sur le pois chiche en agroforesterie en 2009 (Villalba, 2009) puis en 2010 (Gaborit-Varlet, 2010).
L'objet de cette étude est donc de mesurer l'impact des arbres sur la culture d’une espèce fixatrice d'azote. Il s'agit notamment d'étudier l'effet de l'ombre dans le système agroforestier mis en place en 1995, et d'évaluer l'influence des rangées d'arbres sur les performances agronomiques et fixatrices du pois fourrager. Pour cela, nous disposons de deux parcelles agroforestières offrant plusieurs modalités, et l'influence des arbres sera estimée par l'étude de la croissance et des
Matériels et Méthodes
1)
Site
Les parcelles étudiées sont situées sur le domaine de Restinclières, une zone de 250 ha située sur la commune de Prades-le-Lez, à 10 km au nord de Montpellier (Hérault, France). Ce domaine appartient au Conseil Général de l'Hérault, et est utilisé en partie pour les expérimentations de plusieurs équipe de recherche dans le cadre d’un projet intégré (projet PIRAT : http://www.agroof.net/PIRAT/). Des parcelles agroforestières ont été plantées en 1995 sur l'initiative de Christian Dupraz, et l'UMR System y réalise des suivis afin d'évaluer la productivité du système.
Le site d'étude comporte plusieurs parcelles d'expérimentation, comportant différentes modalités. Cependant, l’ensemble des parcelles est géré par un agriculteur qui y applique ses itinéraires techniques classiques. Deux parcelles agroforestières concernent ici la thématique du stage : la parcelle « Agroforesterie 2 » appelée « A2 » est composée d'une dizaine de lignes d'arbres orientées selon l'axe Est-Ouest, tandis que la parcelle « Agroforesterie 3 » dite « A3 » comporte 6 lignes d'arbres orientées selon l'axe Nord-Sud (Figure 1). Chacune des parcelles possède une partie sans arbre constituant le « témoin agricole », le dispositif présente donc quatre sous-systèmes à étudier (Figure 2):
« AF2 » est le système agroforestier de A2 (4,8 ha), les arbres y sont plantés selon l'axe Est-Ouest « AF3 » est le système agroforestier de A3 (1,8 ha), les arbres y sont plantés selon l'axe Nord-Sud « TA2 » est le témoin agricole de A2 (1,4 ha), et correspond à la partie Nord de la parcelle A2 « TA3 » est le témoin agricole de A3 (0,5 ha), et correspond à une zone déboisée de la parcelle A3
2)
Matériel Biologique
Les arbres
Les deux parcelles étudiées respectent les critères suivants : à la plantation, les arbres était disposés tous les 4m en lignes, séparées de 13m les unes des autres. Une éclaircie a été faite en 2004, pour supprimer la moitié des arbres et arriver à une densité d’environ 100 tiges/ha. Cette éclaircie n’a pas été faite de façon régulière : sur 4 arbres consécutifs, on a gardé les 2 plus beaux L'essence choisie est un noyer hybride Juglans regia x nigra. Cet arbre est largement utilisé
en agroforesterie puisqu'il présente nombre de caractéristiques qui semblent bien appropriées à ce système. En effet, son système racinaire plutôt en profondeur semble gêner dans une moindre mesure la culture adjacente. Son débourrement tardif retarde la compétition pour la lumière à des stades phénologiques avancés de la culture, et atténue ainsi son impact. Sa croissance rapide, son architecture verticale (Fady et al., 2003) et un entretien régulier par de l'élagage latéral permet la production de bois de bonne qualité et de valeur, destiné à la construction.
Une des particularités de cet arbre, est sa grande hétérogénéité due à son statut hybride. Celle-ci est notamment visible sur le plan phénologique, présentant une grande variabilité sur la date de débourrement. On peut supposer que la culture associée est donc différemment affectée selon la précocité de l'arbre à proximité.
La culture
Les deux parcelles A2 et A3 sont soumises à la même rotation de culture depuis 1995, c'est-à-dire une alternance de deux à trois années de blé dur d'hiver avec une année de colza. En 2009 (uniquement sur de petites placettes) et en 2010 (sur parcelle entière) est expérimenté l'introduction d'une légumineuse dans le système : le pois chiche de printemps (type Kabuli) et le pois protéagineux sur le reste des parcelles en 2010. Depuis, le pois protéagineux a remplacé le colza dans les rotations.
Cette année, la culture est du pois protéagineux d'hiver, de variété Blizzard. Le semis, précédé d'un labour et suivi d'un désherbage, a eu lieu le 5 Décembre 2012, les conditions météorologiques ne l'ayant pas permis plus tôt. La densité de semis était de 80 grains/m² (soit 250kg/ha), visant une levée de 60 plantes/m².
Le pois Pisum sativum L. est une plante annuelle à croissance indéterminée, de ce fait une même plante porte des organes de différents stades de maturité, arrivés successivement au cours du développement caulinaire.
Comme toutes les légumineuses, le pois est capable de réaliser une fixation symbiotique de l'azote atmosphérique. Le symbiote Rhizobium leguminosarum est naturellement présent dans le sol, et induit la formation de nodules sur la racine primaire et les racines latérales de premier ordre. Son taux de fixation est estimé à 40mg N/m²/degré-jour, soit environ 7kg N/ha (Voisin et.al., 2002). On estime qu'une culture de pois laisse un reliquat d'azote de 20 à 50 kg/ha sur la parcelle cultivée, constituant ainsi une fertilisation naturelle potentiellement utilisable par la culture suivante.
Dispositif
L'étude vise à mettre en évidence l'influence, s'il y en a, du noyer sur le pois. Trois paramètres sont ici analysés :
– la présence de l'arbre, par comparaison des parcelles agroforestières avec les témoins agricoles
– l'orientation de la rangée d'arbres, par comparaison entre AF2 et AF3
– la distance de l'arbre, en comparant les mesures effectuées en « bordure » et en « milieu de l'allée »
En effet, trois modalités sont étudiées en parcelles agroforestières afin de prendre en considération l'effet de la distance de la ligne d'arbres sur la culture. Ces trois modalités situées dans les inter-rangs de AF2 et AF3 sont placées sur des transects perpendiculaires à la ligne d'arbres, entre deux noyers bien développés, et respectant les distances suivantes : à 2m, à 6,5m et à 11m d'un arbre. Ces modalités permettent d'étudier deux facteurs liés à la présence des arbres : tout d'abord l'effet de la distance de la ligne d'arbre sur la culture selon sa position dans l'inter-rang, et ensuite l'exposition dont elle bénéficie.
Dans le premier cas, les modalités « 2m » et « 11m » sont regroupées en une seule : l'effet « bordure » (celles-ci étant en fait à égale distance de l'arbre le plus proche), et sert de comparaison à la modalité « milieu de l'allée » correspondant aux mesures à « 6,5m ». Dans le second cas, on tient compte de l'orientation de la ligne d'arbre qui influence le micro-climat directement sous la canopée. Ainsi, le pois cultivé au nord de l'inter-rang sur la parcelle AF2, est exposé « Sud », la ligne de noyers le protégeant du Nord, et ainsi de suite pour les autres cas (Figure 3).
Sur chaque parcelles A2 et A3, sont disposées 20 placettes de 1m², chacune sont repétées: 5 répétitions en témoin agricole et 5 transects sur l'inter-rang tels que décrits ci-dessus (Figure 3, Tableau I). Ces placettes permettent d'effectuer un suivi de la croissance des pois selon les différentes modalités. Les pois de ces placettes seront récoltés avant la moisson des parcelles par l’agriculteur, et permettront de faire une estimation du rendement et de ses composantes.
Lorsque des prélèvements de plantes sont effectués, ils respectent aussi ces modalités mais sont réalisés en dehors des transects.
Variables biologiques étudiées
La première mesure réalisée a été le comptage du nombre de plantes dans les placettes d’1 m² (le 19/04/13), afin d'estimer le pourcentage de levée du pois, par rapport à un semis initial de 80
grains/m².
Sur ces mêmes échantillons, sont aussi mesurés : la surface foliaire à l'aide d'un planimètre, la biomasse aérienne, après séchage pendant 48 h à l’étuve à 60 °C, en séparant la biomasse des tiges et des feuilles, ce qui permet de calculer la masse surfacique foliaire. Ces mesures correspondent à une estimation de la croissance des plantes à un stade végétatif précoce.
L’un des services attendus par la culture d’une légumineuse est l’apport d’azote au système. C'est donc une estimation de l'activité fixatrice qui est visée, que l'on quantifie par la mesure du nombre et du poids des nodules présents sur les plantes. La fixation de l'azote atmosphérique est la plus importante en période de floraison, les prélèvements ont donc été effectués au début de celle-ci, le 26 Avril 2013. A cette date, on a prélevé les parties aériennes et souterraines de 3 plantes consécutives sur un rang selon les différentes modalités : AF2 et AF 3, 3 distances aux arbres et TA 2 et TA3, avec 5 répétitions. Les racines ont été triées pour en prélever les nodules qui ont été comptés, puis séchés à l’étuve à 60 °C pendant 48 h et pesés.
Sur ces mêmes échantillons, sont aussi mesurés : la surface foliaire à l'aide d'un planimètre, la biomasse aérienne, après séchage pendant 48 h à l’étuve à 60 °C, en séparant la biomasse des tiges et des feuilles, ce qui permet de calculer la masse surfacique foliaire. Ces mesures correspondent à une première estimation du rendement à un stade végétatif précoce.
La récolte du pois se déroulant en général fin-Juin début-Juillet, les résultats de l'expérience ne sont pas disponibles à ce jour. Cependant, une autre mesure plus tardive que la précédente, permet d'effectuer une estimation du rendement final. Le 3 Juin 2013, de nouveaux prélèvements de 3 plantes consécutives sont effectués, permettant la mesure de la biomasse aérienne après passage à l'étuve à 60°C pendant 48h, et l'estimation du nombre de grains et du nombre de gousses par plante jeunes et matures, du nombre de grains et du poids de grains dans les gousses matures par plante. Nous avons fait la distinction entre les gousses matures et jeunes étant donné le stade phénologique de la plante. Le pois a une croissance indéterminée, même à cette période les plus jeunes phytomères possèdent encore des fleurs. Il est donc possible que le nombre de gousses par plante évolue encore. Cependant la mesure des composantes du rendement à ce stade est intéressante parce que le pois a bénéficié d'une saison assez fraîche et pluvieuse jusque-là, alors que le mois de juin s'est montré bien plus chaud et sec par la suite. Or, de la précédente étude, on retient que le pois chiche avait souffert de la sécheresse de la région.
La présence des arbres affecte sur différents paramètres l'environnement de la culture. L'objet de l'étude se focalise ici sur l'impact de l'ombrage sur la croissance des pois. En effet, dans ce système, on peut supposer que l'interférence racinaire des arbres sur la culture est minime puisque chaque année un labour profond a lieu dans l'inter-rang, permettant en théorie d'éviter le développement des racines de noyers dans la zone cultivée. De ce fait on peut estimer que la compétition pour les nutriments et pour l'eau entre la strate arborée et la culture est moindre.
C'est donc la mesure du rayonnement atteignant la culture qui nous intéresse ici. Deux types de mesures sont ainsi effectués.
Tout d'abord, des photos hémisphériques sont réalisées à l'aide d'un objectif fisheye, à deux dates (le 13/04/13 et le 10/06/13) correspondant respectivement au début et à la fin du débourrement des noyers, (Figure 4). Ces images sont par la suite traitées grâce au logiciel GLA (Gap Light Analyser) qui transforme ces informations en rayonnement global (c'est-à-dire la somme du rayonnement direct et du rayonnement diffus reçu par jour) incident à l’emplacement de la photo. Ce rayonnement peut être exprimé en Watt/m²/jour, ou bien rapporté au rayonnement global en dehors de tout couvert végétal, propre aux données spatiales de la station, en %.
Le rayonnement photosynthétiquement actif (PAR Photosynthetic Active Radiation) a aussi été suivi. Des capteurs PAR ont été installés le 17/04/13 (Figure 5), au début du débourrement des arbres. Ils étaient reliés à une centrale d’acquisition de données (CR1000 – Campbell Scientific) qui enregistrait une moyenne de µmol/m²/s, toutes les heures, permettent par la suite de faire des moyennes journalières. Ces capteurs étaient disposés sur un transect dans chaque parcelle AF2 et AF3 selon les modalités précitées. Les données de ces capteurs ont été comparées à celle d’une station météo Campbell installée dans le témoin agricole de la parcelle A2, qui représente l’exposition plein soleil.
Analyse statistique
Les données mesurées lors de cette expérimentation sont quantitatives, et font l'objet d'une analyse par tests statistiques. Les données biologiques récoltées sont étudiées par analyse de variance à un facteur pour estimer l'influence de l’emplacement par rapport aux arbres sur le comportement du pois. On analyse également les données quantitatives environnementales de cette manière, afin de mettre en avant les caractéristiques lumineuses de chaque emplacement.
Les variables quantitatives n'étant pas toutes issus des mêmes individus statistiques, on effectue une régression linéaire à partir des moyennes des différentes variables.
Toutes ces analyses sont effectuées sur le logiciel R. Enfin, une analyse graphique des tendances est réalisée.
Résultats
Influence de l'arbre sur l'environnement lumineux de la culture
Mesure du rayonnement global le 13/04/13
Aux premiers stades du cycle végétatif des arbres (début du débourrement pour les arbres les plus précoces), on mesure une influence de l'arbre sur le rayonnement global qui atteint le sol de l’inter-rang (Figure 6). En effet, la moyenne du rayonnement global en milieu d'allée est significativement plus élevée qu'en bordure (p=0,01 au seuil de 5%). Cependant, à ce stade phénologique de l'arbre, la différence n'est pas très grande (seulement de l'ordre de 5% du rayonnement incident), et les arbres n'affectent donc la culture que dans une moindre mesure. Mesure du rayonnement global le 10/06/13
Deux mois plus tard, alors le feuillage des arbres s'était bien développé et que tous les arbres avaient débourré, e rayonnement global arrivant sur la culture avait diminué en moyenne de 34% sur la AF2 et de 39% la AF3par rapport aux mesures du mois d'Avril (Figure 6). On peut remarquer que l'effet bordure est devenu négligeable, et que c'est désormais l'exposition qui influence le plus l'environnement lumineux de la culture. En effet, le test d'analyse de variance est significatif au seuil de 5% (p=0,002), et montre que l'exposition Sud et le milieu de l'allée de AF2 reçoivent en moyenne plus de lumière que les autres emplacements (de l'ordre de 60% du rayonnement incident), tandis que les placettes orientées Ouest en reçoivent moitié moins (33%).
La parcelle AF2 semble montrer plus d'homogénéité, avec un maximum de différence de 10% entre les expositions Nord et Sud, tandis que les intervalles se creusent à 20% entre l'exposition Ouest et le milieu de l'allée dans AF3, ce qui traduit une plus grande hétérogénéité. Ces résultats vont contre les conclusions d'une précédente étude réalisée sur le même site (Gaborit-Varlet, 2010), certainement à cause du vieillissement du système.
Capteurs PAR
Les capteurs positionnés sur deux transects perpendiculaires aux lignes d'arbres dans AF2 et AF3 devaient suivre en continu les variations de radiations. Cependant des soucis de calibration rendent l'utilisation de ces données difficile. Il n'y a pas de corrélation positive entre les mesures de rayonnement global des photos hémisphériques et les mesures de PAR. En effet, il est possible de convertir des W/s de rayonnement global (Rg) en µmol/m²/s de PAR selon la relation : 1W/s de Rg = 2,02µmol/m²/s PAR.
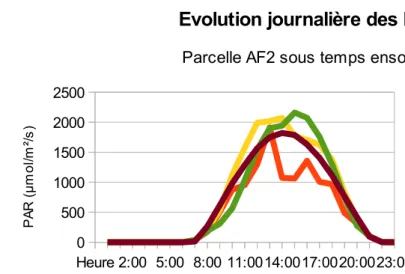
intéressante à considérer. En effet, l'allure des courbes représentant les PAR au cours de la journée par temps ensoleillé confirme la disparité Nord-Sud dans la parcelle AF2 (Figure 7a). Les pics observés, particulièrement visibles sur la courbe représentant la lumière reçue sur la placette exposée « Nord », correspondent certainement au passage des rayons du soleil entre le feuillage de deux arbres. Le côté exposé « Nord » est clairement désavantagé, car le cumul des PAR qu'il reçoit en une journée est très inférieur à celui du milieu de l'allée ou de l'exposition « Sud » (en l’occurrence pour le 11 Mai 2013 : avec un déficit de 5240 et de 3824µmol/m²/s par rapport au milieu de l'allée et à l'exposition « Sud » respectivement). On remarquera que le témoin agricole donne des valeurs anormalement inférieures au système agroforestier, ce qui peut s'expliquer par un défaut de calibration.
Dans la parcelle AF3, on s'attendait à observer une permutation entre des valeurs fortes le matin aux placettes exposées à l'Est et faibles à l'Ouest, et le contraire l'après-midi. C'est ce qui était notamment visible dans les résultats de la précédente étude sur cette parcelle, en 2010 (Annexe I). Ce « switch lumineux » n'est pas observable à partir de nos données (Figure 7b), et on pose l'hypothèse que c'est la croissance des arbres qui en est responsable. En effet, il semble que la hauteur qu'ils atteignent aujourd'hui ne permette plus le passage de la lumière dans certains endroits. Le capteur du milieu de l'allée de l’AF3 est éclairé tard dans la journée, son éclairement maximal arrive environ deux heures après celui du milieu de l'allée dans l’AF2. De ce fait, c'est la bordure exposée à l'Est qui cumule le plus d'énergie lumineuse en une journée : par exemple pour le 6 Mai 2013, la somme des PAR de l'exposition « Est » dépasse respectivement de 3377 et de 2584 µmol/m²/s celle de l'exposition « Ouest » et du milieu de l'allée AF3. L'orientation des rangées d'arbres de la parcelle explique ce phénomène. En effet, les noyers ne sont pas parfaitement alignés sur l'axe Nord-Sud (mais plutôt NNE-SSO), et offrent donc un angle entre les rayons incidents et la rangée d'arbres plus important le matin que l'après-midi.
Les différences de rayonnement détaillées ci-dessus ne sont visibles que par temps ensoleillé. En annexe II est présenté un exemple d'évolution des PAR journaliers sur les deux parcelles par temps couvert, relevé le 20 Avril 2013, où les courbes se superposent.
Impact sur la levée
C'est essentiellement la présence de l'arbre qui influe sur la levée de la graine. La moyenne de levée des deux parcelles est bien plus importante au milieu de l'allée (85,5%) par rapport au témoin agricole (71,6%) et au pois situé en bordure des arbres (67,9%), avec une différence significative au seuil de 5% (p=0,006) (Figure 8a). Le test de comparaison de moyenne entre le témoin agricole et le milieu de l'allée est aussi significatif.
Cependant, il n'est pas exclu que l'exposition influence aussi cette variable (Figure 8b). Les résultats suggèrent que les expositions Sud et Ouest favoriserait la germination des graines (de respectivement 71,7% et 72,7%) par rapport aux expositions Nord et Est (66,7% et 60,5% respectivement).
Mesure de la biomasse aérienne au stade de floraison
On observe que la biomasse aérienne moyenne est plus importante sur les plantes du milieu de l'allée que sur les bordures ou les témoins agricoles, dans les parcelles A2 et A3 (Figure 9). La masse surfacique des feuilles ne présente pas de différence significative entre les différents échantil-lons, néanmoins il semblerait que les feuilles soient un peu plus charnues dans les témoins agricoles (Annexe III) ce qui est conforme aux résultats de Evans & Poorter, 2001.
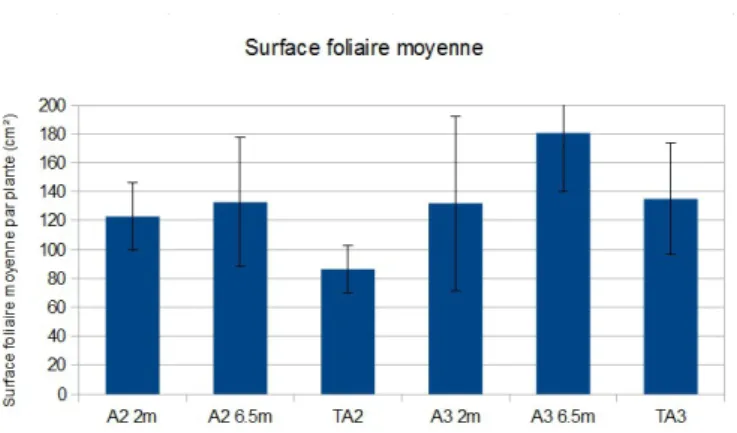
La surface foliaire est plus variable : elle est plus élevée sur les échantillons issus des milieux des allées, et est globalement supérieure dans la parcelle A3 que dans A2 (Figure 10).
Il est donc probable que les allées d'arbres protègent le pois à ses premiers stades de développement, influençant positivement sa croissance, et que même les plantes des bordures produisent plus de biomasse aérienne que les témoins agricoles (ceci est vrai pour les deux parcelles). Il semble que les plantes profitant d'une zone un peu plus humide et ombragée en système agroforestier produisent de plus grandes feuilles, surtout en milieu d'allée. Ceci semble cependant ne pas pouvoir s'appliquer aux plantes exposées à l'Ouest, celles-ci montrant au contraire de plus petites feuilles que les autres sous-systèmes. Les plantes des témoins agricoles ont quant à elles tendance à faire des feuilles plus petites et plus épaisses.
Les nodules au stade floraison
L'analyse des données de biomasse et de comptage des nodules est difficile, car les écart-types très élevés. Le nombre de nodules par plante varie cependant de manière significative en fonction de la distance à l’arbre (Figure 11) et de l'exposition, avec des probabilités critiques respectives de 0,04 et de 0,01 au seuil de 5%. Le nombre de nodules par plante est en effet plus important en milieu d'allée qu'en témoin agricole, mais ceci est surtout vrai pour la parcelle A2 ; et l'exposition Ouest semble la plus défavorable à l'élaboration des nodosités.
Le poids des nodules est très variable, les nodosités semblent plus grosses en parcelle agroforestière AF2 qu'en témoin agricole TA2, tandis que c'est la tendance inverse qui est observée dans la parcelle A3, même si la variabilité est moins visible (Annexe VI).
parcelles avait mis en évidence une légère tendance de la parcelle A2 à avoir des nodules plus petits et plus nombreux qu'en A3. Cette tendance est vérifiée par nos données.
Mais il est notable que le témoin agricole TA2 sort du lot : il possède en effet les plus petits nodules et les moins nombreux de toutes les modalités. Or, on sait que le témoin agricole TA3 est bien plus soumis à une ambiance « agroforestière » que le TA2. En effet, ce dernier est issu de l'abattage récent d'arbres devenus menaçants pour la ligne électrique, il reste entouré d'arbres et de haies à proximité, et constitue donc un milieu bien moins ouvert que le TA2 (Figure 1). On peut ainsi émettre l'hypothèse, sur la base de nos données, que l'ambiance agroforestière accroît la capacité des pois à établir une fixation symbiotique, puisqu'ils développent de plus gros nodules et en plus grand nombre.
Estimation de l’influence des arbres sur le rendement
Les mesures de gousses faites en juin mettent en exergue le retard phénologique des plantes en systèmes agroforestiers par rapport au témoin agricole (Figure 12a). En effet, les témoins agricoles possèdent significativement plus de gousses matures que la moyenne des emplacements en AF2 et AF3 (p=0.009 au seuil de 5%). La réduction de lumière est donc certainement un facteur retardant la croissance du pois. On remarque que le pois du milieu d'allée en AF2 ne présente pas de retard phénologique, et a même tendance à se montrer plus mature que son témoin agricole TA2, avec un ratio gousses matures/gousses jeunes plus élevé. La parcelle A3 présente par contre des ratios bien plus faibles et donc un retard de croissance notable (ratio moyen de 0,56 en AF3 et de 3,72 en TA3, contre 5,05 en AF2 et 6,00 en TA2).
L'effet de la bordure sur la phénologie du pois n'est pas flagrante dans AF3 ; tandis qu'elle est incontestable en AF2 : le ratio du milieu d'allée est deux fois plus important que celui de la bordure. Comme on pouvait s'y attendre, c'est le côté de l'allée exposé Nord qui a le plus de retard, tandis que le côté exposé Sud est presque équivalent au témoin agricole (ratio de 0,94 et 5,8 respectivement pour les expositions Nord et Sud) (Figure 12b).
Le poids moyen d'une gousse jeune est sensiblement le même d'un système à l'autre.
Par contre, le poids moyen d'une gousse mature varie significativement au seuil de 5%, selon la parcelle (p=0,0004). Les gousses matures sont légèrement plus lourdes dans la parcelle A2 que dans la A3. Il n'y a pas de différence visible à ce stade phénologique sur le nombre moyen de grains par gousse mature entre les modalités, la variation du poids des gousses résulte d'une différence de poids moyen d'un grain.
montre en effet que les pois exposés au Nord produisent significativement les plus petites graines et ceux exposés au Sud les plus grosses (respectivement : 62,8mg et 94,3mg de poids sec en moyenne). La différence est plus légère dans la parcelle AF3, mais tout de même notable : l'exposition Est produisant de plus gros pois que l'exposition Ouest (84,1 et 68,5mg respectivement). Les témoins agricoles TA2 et TA3, donnent en moyenne de plus grosses graines (93,5 et 88,8 mg respectivement) que la moyenne des emplacements agroforestiers. Il y a cependant une divergence de résultats concernant les relevés de milieu d'allée, les placettes de AF2 montrent en effet de belles performances avec un poids moyen d'un grain de 90,2mg, alors que les plantes de AF3 produisent les plus petites graines, d'environ 62,2mg. Ceci peut être expliqué par la différence de rayonnement atteignant la culture : les capteurs montrent en effet que AF3 reçoit moins de PAR que AF2 en milieu d'allée.
Mesure de la biomasse aérienne au stade remplissage des gousses
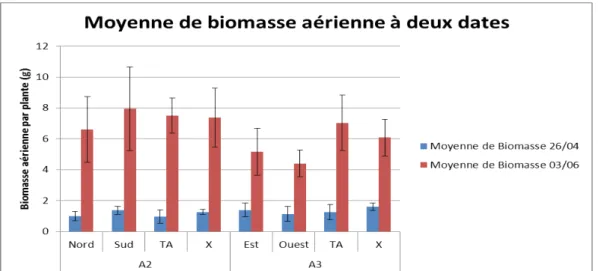
La biomasse aérienne totale par plante est significativement plus grande en A2 qu'en A3 (Figure 14a), avec une probabilité critique de 0,006 au seuil de 5%. La biomasse aérienne par plante est la plus élevée chez les plantes récoltées exposition « Sud » (7,94 g de matière sèche), elle est du même ordre en TA2 et en milieu d'allée de AF2 (7,513 et 7,367g respectivement) (Figure 14b). La parcelle A3 présente des valeurs, plus faibles, ne dépassant pas 6 g de MS/plante en agroforesterie, et pas plus de 7 g en témoin agricole. Une corrélation existe entre les composantes du rendement et la biomasse aérienne estimée par plante le 03/06. En effet, la biomasse aérienne dépend essentiellement du poids des feuilles et du poids des gousses par plante (matures et jeunes).
Une régression linéaire entre les moyennes de biomasses estimées le 26/04 et le 03/06 et les moyennes de Rayonnement global, montre que la biomasse du pois au mois de juin est positivement corrélée avec le Rayonnement global intercepté sous la canopée à cette période, et négativement avec la biomasse du mois d'Avril. Ceci confirme que l'impact de l'ombre arrive aux derniers stades de développement du pois. L'orientation des arbres selon l'axe Est-Ouest semble avantageuse au début de la croissance du pois, la biomasse aérienne en AF3 le 26/04 étant plus grande que celle de AF2 (Annexe V). Mais, les tendances s'inversent ensuite au fur et à mesure que les arbres débourrent. En juin, l'estimation du rendement donne de meilleurs résultats pour la parcelle AF2 que AF3, et ceci est directement lié aux conditions lumineuses mesurées le 10/06, la parcelle AF2 étant plus éclairée.
En prenant en considération la biomasse estimée par plante le 03/06, et la densité de levée au m², on peut estimer la biomasse finale des différents systèmes. Ces données sont à prendre avec
précautions, puisque n'y sont pas inclus la perte de biomasse par résidu agricole, et les pertes associées à la récolte (manœuvre, verse...).
On trouve donc les rendements suivant : – 44.2 q/ha en AF2
– 44.9 q/ha en TA2 – 29.9 q/ha en AF3 – 38.5 q/ha en TA3
En tenant maintenant compte des lignes d'arbres non cultivées, on obtient en moyenne : – 39.0 q/ha en AF2
– 26.5 q/ha en AF3 – 41.6 q/ha en TA
On observe donc à ce stade de la culture, une perte potentielle de 6.5% pour la parcelle AF2 et de 36% pour la parcelle AF3 par rapport à la moyenne des témoins agricoles. En comparant ces données avec les valeurs estimées pour le pois chiche en 2010, on remarquera que la parcelle AF3 est bien moins productive que précédemment, la perte de rendement par rapport au témoin agricole n'étant alors que de 25%.
Discussion
La variable environnementale suivie dans cette étude , à savoir la lumière, semble être un facteur crucial influençant le développement de la culture en agroforesterie. En effet, nos mesures montrent une diminution de plus de 30% du rayonnement global atteignant le sol après le débourrement des arbres. De plus, la présence des arbres accroît l'hétérogénéité au sein des parcelles, le rayonnement incident sur les différents emplacements des parcelles étant clairement dépendant de l'exposition associée. Les différentes placettes montrent des patterns d'évolution de PAR distinctes au cours de la journée, et le cumul des PAR sur une journée montre aussi une disparité, celle-ci étant certainement déterminante pour la réponse des pois.
On peut supposer que l'ambiance agroforestière aux premiers stades de développement de la culture est plutôt bénéfique pour le pois. On observe en effet un taux moyen de levée de la graine meilleur en agroforesterie qu'en parcelle agricole, mais ceci est surtout dû à une germination largement supérieure en milieu d'allée, peut-être liée à un effet brise-vent. De ce fait, la densité semble être de manière générale plus importante en système agroforestier qu'en témoin agricole.
Les mesures de biomasse au stade floraison du pois, correspondant au début du débourrement des noyers les plus précoces, montrent encore une influence positive des arbres sur la croissance des pois. La biomasse aérienne est plus élevée en parcelle agroforestière qu'en témoin agricole à cette période. Cependant cette tendance s'inverse lorsque le pois approche de la maturité, en effet la biomasse aérienne est supérieure dans les témoins agricoles en Juin. Ceci est directement corrélé avec la chute de rayonnement global atteignant la parcelle due au débourrement des noyers. L'impact de l'ombre des arbres se fait donc sentir à la fin du cycle du pois. Néanmoins, on peut supposer que cette incidence aurait été d'autant plus importante pour un pois de printemps, son cycle végétatif commençant plus tard dans l'année.
En plus d'une influence négative des arbres sur la biomasse produite, la baisse de rayonnement global en agroforesterie retarde la phénologie du pois. Mais ceci est encore une fois très variable selon l'exposition (l'exposition « Sud » est bien en avance par rapport aux autres sous-systèmes), ce qui accentue encore l'hétérogénéité au sein des parcelles agroforestières. Cette différence de maturité du pois est incontestablement pénalisante pour l'agriculteur, puisque la récolte ne peut se faire en plusieurs fois, pour des questions de logistique notamment.
agroforestier. Cette observation est valable si on écarte les données du TA3, qui paraît subir une influence des arbres du fait de sa petite surface, et sa proximité avec le AF3.
L'étude montre une différence notable de la réponse du pois dans chacune des parcelles. Il est généralement conseillé de planter ses arbres en agroforesterie selon un axe Nord-Sud (comme dans la AF3), afin de faire bénéficier du soleil direct au zénith, à une portion maximale de la culture, et de réduire ainsi les écarts entre les bordures (Dupraz & Liagre, 2008). Cependant, c'est totalement l'inverse qui est observé cette année à Restinclières. L’hétérogénéité et la perte de rendement sont plus importantes dans la parcelle AF3 que dans la AF2. Il faut noter que la croissance des arbres n'est pas homogène sur l'ensemble du site, notamment à cause de l'inégalité de profondeur des sols alluviaux. Il semblerait que la croissance des arbres de la parcelle AF3 a été plus rapide que ceux de la AF2, et que de ce fait, la compétition avec la culture adjacente soit plus importante. Dans la parcelle A3, les arbres faisait en moyenne 10.4 m de haut en mars 2010, 12.2 m en mars 2013, ce qui veut dire que leur hauteur est aujourd’hui, en moyenne pratiquement égale à la largeur de l’inter-rang
Malheureusement, le système n'est âgé que de 18 ans, et la récolte des noyers, n'est prévue qu'au bout de quarante ans, ce qui laisse à supposer une diminution forte de productivité pour les cultures à venir. La question est donc de mesurer la rentabilité du système, en terme de performances agronomiques des cultures années après années, accompagnée du gain rapporté par la production de bois à maturité du système. Les conseils pour la plantation de parcelles agroforestières prévoient aujourd’hui des allées plus larges, au moins égales à la hauteur finale estimée des arbres pour pouvoir cultiver jusqu’à la fin.
Conclusion
L'étude met ici en avant les difficultés de produire en système agroforestier. La baisse de luminosité sous des arbres adultes joue incontestablement sur la diminution de rendement et sur le retard phénologique de la culture. De plus, la présence des arbres influe de manière différente selon l'exposition, et augmente l'hétérogénéité au sein de la parcelle, ce qui accentue les difficultés rencontrées pour la récolte. En effet, cette différence de luminosité prodiguée par les arbres constitue un patchwork de micro-climats, bien distincts du témoin agricole classique, s'accompagnant certainement d'une modification de la température et de l'humidité du sol (Gaborit, 2010), et du vent. On distingue bien évidemment un environnement plutôt favorable à la culture en milieu d'allée, bénéficiant d'un micro-climat « protégé » ou « tamponné », et plutôt défavorable en bordure des arbres par le manque de lumière et aussi potentiellement par une augmentation de la compétition racinaire (avec les arbres ou les adventices qui se développent en bordure). La variation d’éclairement est cependant fortement dépendante de l'exposition dont bénéficie la culture, qui sera incontestablement favorisée par une exposition « Sud ».
Le site de Restinclières offre la possibilité d'étudier l'évolution du système agroforestier sous climat méditerranéen. La reproduction des expérimentations années après années, permet d’affiner le système, et de corriger au mieux les interférences des arbres afin de le rendre le plus productif possible. Cependant, le schéma de plantation qui a été prévu en 1995 montre également ses limites. Il serait éventuellement possible de relancer la productivité des cultures en supprimant une ligne d’arbres sur deux.
La problématique de l'azote s’insère directement dans l'intégration des légumineuses dans les rotations culturales. Il serait de ce fait pertinent d'étudier de manière plus approfondie l'impact du système agroforestier sur la fixation symbiotique des légumineuses, notamment par analyse de l'azote de la culture suivant la légumineuse par des méthodes isotopiques (abondance naturelle en
Bibliographie
P. Balandier, C. Dupraz (1999) Growth of widely spaced trees. A case study from young agroforestry plantations in France. Agroforestry Systems 43 : 151-167
J.R. Bohanek, J.W. Groninger (2005) productivity of european black alder (Alnus glutinosa) interplanted with black walnut (Juglans nigra) in illinois, U.S.A.. Agroforestry Systems 64: 99-106
V. Chifflot, G. Bertoni, A. Cabanettes, A. Gavaland (2006) Beneficial effects of intercropping on the growth and nitrogen status of young wild cherry and hybrid walnut trees. Agroforestry Systems 66 : 13-21
G. Corre-Hellou, J. Fustec, Y. Crozat (2006) Interspecific competition for soil N and its interaction with N2 fixation, leaf expansion, and crop growth in pea-barley intercrops. Plant and Soil 282 : 195-208
R. Dixon, D. Kahn (2004) Genetic regulation of biological nitrogen fixation, Nature Reviews 2 : 621-631
C. Dupraz, F. Liagre (2008) Agroforesterie- Des arbres et des cultures. Ed. France Agricole
Evans J.R., Poorter H. (2001). Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant, Cell and Environment, 24, pp.755-767
B. Fady, F. Ducci, N. Aleta, J. Becquey, R. Diaz Vazquez, F. Fernandez Lopez, C. Jay-Allemand, F. Lefèvre, A. Ninot, K. Panetsos, P. Paris, A. Pisanelli, H. Rumpf (2003) Walnut demonstrates strong genetic variability for adaptative and wood quality traits in a network of juvenile field tests across Europe, New Forests 25 : 211-225
A.C. Franke, G. Laberge, B.D. Oyewole, S. Schulz (2008) A comparison between legume technologies and fallow, and their effects on maize and soil traits, in two distinct environments of the West African savannah, Nutr Cycl Agroecosyst 82 : 117-135
P. Gaborit-Varlet (2010), Mémoire de fin d'études : Comment l'agroforesterie modifie-t-elle les facteurs qui conditionnent les performances agronomiques du pois chiche ?, Agrocampus Ouest, Rennes
K.M. Goh, I. Mansur, D.J Mead, G.B. Sweet (1996) Biological nitrogen fixing capacity and biomass production of different understorey pastures in a Pinus radiata-pasture agroforestry system in New Zealand, Agroforestry Systems 34 : 33-49
A.M. Hejl, K.L. Koster (2004) Juglone disrupts root plasma membrane H⁺-ATPase activity and impairs water uptake, root respiration, and growth in soybean (Glycine max) and corn (zea mays). Journal of Chemical Ecology, Vol. 30, No. 2
S. Jose, A.R. Gillespie, S.G. Pallard (2004) Interspecific interactions in temperate agroforestry. Agroforestry Systems 61 : 237-255
S. Jose, A.R. Gillespie, J.R. Seiffert, D. J. Biehle (2000) Defining competition vectors in a temperate alley cropping system in the midwestern USA -2. Competition for water. Agroforestry Systems 48 : 41-59
C.H. Lin, R.L. McGraw, M.F. George, H.E. Garrett (2001) Nutritive quality and morphological development under partial shade of some forage species with agroforestry potential. Agroforestry Systems 53 : 269-281
R. L. McGraw, W. T. Stamps, J. H. Houx, M. J. Linit (2008) Yield, maturation, and forage quality of alfalfa in a black walnut alley-cropping practice. Agroforestry Systems 74 : 155-161
M.R. Mosquera-Losada, J.H. McAdam, R. Romero-Franco, J.J. Santiago-Freijanes, A. Rigueiro-Rodríguez (2009) Definitions and components of agroforestry pratices in Europe. In A. Rigueiro-Rodríguez et al., eds, Agroforestry in Europe : Current status and future prospects, Springer Science, Business Media B. V., pp 3-19
E. Munyanziza, H.K. Kehri, D.J. Bagyaraj (1997) Agricultural intensification, soil biodiversity and agro-ecosystem function in the tropics : the role of mycorrhiza in crops and trees. Applied Soil Ecology 6 : 77-85
K. Nerlich, S. Graeff-Hönninger, W. Claupein (2013) Agroforestry in Europe : a review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agroforestry Systems 87 : 475-492
M. Oelbermann, R.P. Voroney (2007) Carbon and nitrogen in a temperate agroforestry system : using stable isotopes as a tool to understand soil dynamics. Ecological Engineering 29 : 342-349
C.A. Palm (1995) Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agroforestry Systems 30 : 105-124
A. Villalba, Gonzales (2009) Etude de l’effet de l’ombre sur la culture du pois chiche. Rapport de fin d’études. Montpellier, UMR System, 2009
A. Vocanson (2006) Thèse : Evaluation ex ante d'innovations variétales en pois d'hiver (Pisum sativum L.) : approche par modélisation au niveau de la parcelle et de l'exploitation agricole, Ecole Doctorale ABIES ; Institut National Agronomique Paris-Grignon
A.S. Voisin, C. Salon, N. G. Munier-Jolain, B. Ney (2002) Quantitative effects of soil nitrate, growth potential and phenology on symbiotic nitrogen fixation of pea (Pisum sativum L.) Plant and Soil 243 : 31-42
G.R. Von Kiparski, A.R. Gillespie (2008) Agroforestry management Effets on plant productivity vectors within a humid-temperate hardwood alley-cropping system. In S. Jose, A.M. Gordon, eds, Toward Agroforestry Design : an ecological approach. Springer, pp 149-173
Annexes
Annexe I: Évolution journalière du rayonnement global dans les parcelles AF2 et AF3, mesures effectuées en 2010 ; tirées du mémoire de stage de Pierre Gaborit-Varlet
-2,00 0,00 2,00 4,00 6,00 8,00 10,00 12,00 R ay onne m en t gl oba l ( W /m ²)
Evolution journalière du Rayonnement global
Exposition Nord Milieu de l'allée Exposition Sud 0,00 1,00 2,00 3,00 4,00 5,00 6,00 7,00 8,00 9,00 10,00 R ay onne m en t gl oba l ( W /m ²)
Evolution journalière du Rayonnement global
Exposition Ouest Milieu de l'allée Exposition Est
Annexe II : Évolution journalière des PAR par temps couvert sur les parcelles AF2 et AF3. Mesures effectuées le 20/04/13. 0 100 200 300 400 500 600 700 P AR (µm ol /m ²/ s)
Evolution journalière des PAR
Exposition Nord Milieu allée Exposition Sud PAR calculé station
0 100 200 300 400 500 600 700 P AR (µm ol /m ²/ s)
Evolution journalière des PAR
Exposition Ouest Milieu allée Exposition Est PAR calculé station
Annexe III : Masse surfacique moyenne des feuilles de poids au stade floraison, récoltés le 26/04/13
Annexe IV : Poids moyen d'un nodule de pois récolté le 26/04/13, en fonction de la distance à l'arbre dans les parcelles A2 et A3.
Annexe VII : Évolution des biomasses aériennes de pois en fonction de la distance à l'arbre : « 2m » pour la bordure ; « 6,5m » pour le milieu de l'allée ; et « TA » comme témoin agricole.
Figure 1 : Vue aérienne du site d'expérimentation, au domaine de Restinclières, à Prades-Le-Lez (Hérault, France), avec les
deux parcelles agroforestières étudiées : « A2 » plantée d'arbres selon l'axe Est-Ouest, et « A3 » selon l'axe Nord-Sud.
Figure 2 : Plan du dispositif avec ses quatre sous-systèmes : « AF2 » et « AF3 », les parties agroforestières des parcelles A2 et
A3, et « TA2 » et « TA3 » leur témoin agricole respectif.
A3
Figure 3a : Disposition des placettes selon un axe Nord-Sud dans la parcelle AF2.
Figure 3b : Disposition des placettes selon un axe Est-Ouest dans la parcelle AF3.
Figure 3 : Schéma de l'emplacement des placettes dans les inter-rangs des parcelles AF2 (figure 3a) et AF3 (figure 3b), selon
les trois modalités « 2m » ; « 6,5m » ; et « 11m » de l'arbre, avec indication de leur exposition.
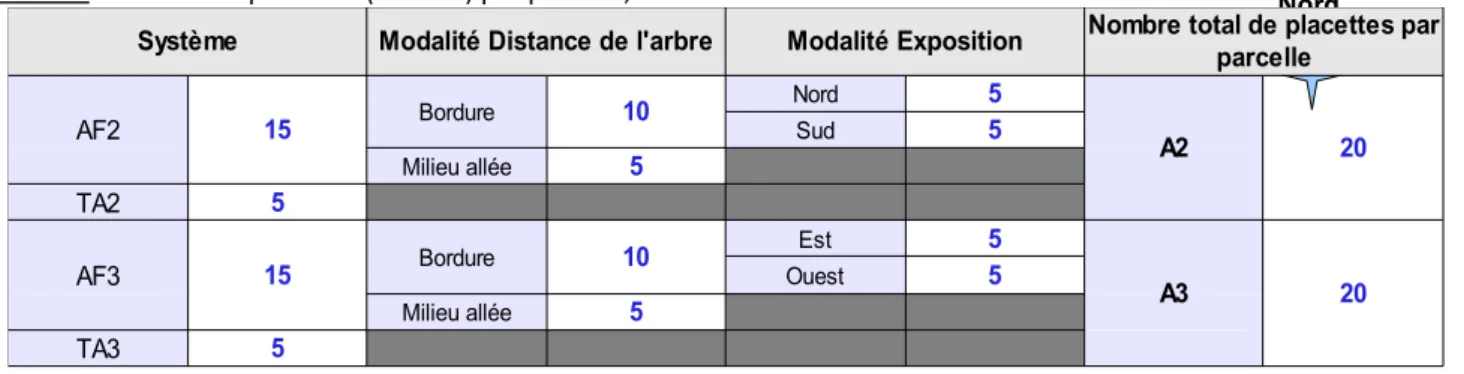
Tableau I : Nombre de placettes (en bleu) par parcelle, selon les modalités étudiées.
Parcelle A3
Distance de l'arbre 2m 6,5m 11m
Exposition Est Ouest
Ligne de
noyers Ligne de noyers
Parcelle A2 Exposition Ligne de noyers 2m Sud 6,5m 11m Nord Ligne de noyers Distance de l'arbre Nord
Système Modalité Distance de l'arbre Modalité Exposition
AF2 15 Bordure 10 Nord 5 A2 20 Sud 5 Milieu allée 5 TA2 5 AF3 15 Bordure 10 Est 5 A3 20 Ouest 5 Milieu allée 5 TA3 5
Nombre total de placettes par parcelle
Légende
Figure 4 : Exemple de photos hémisphériques effectuées au même emplacement le 13/04/13 (à gauche) et le 10/06/13 (à
droite).
Figure 5 : Installation des capteurs PAR sur un transect perpendiculaire à deux lignes d'arbres, deux capteurs à 2m des lignes
Figure 6 : Représentation du Rayonnement global (en % du rayonnement incident) atteignant le sol en parcelle
agroforestière avant et après le débourrage des arbres, le 13/04/13 (en bleu) et le 10/06/13 (en rouge). Plusieurs modalités sont ainsi évaluées : en parcelle « A2 », les expositions « Nord » et « Sud », en parcelle « A3 » les expositions « Est » et « Ouest ». La modalité notée « X » correspond aux mesures du milieu des allées dans chacune des parcelles.
Figure 7a : Évolution journalière des PAR sous temps ensoleillé sur AF2
Figure 7b : Évolution journalière des PAR sous temps ensoleillé sur AF3
Figure 7 : Évolution journalière des PAR sous temps ensoleillé sur les parcelles AF2 (Figure 7a) et AF3 (Figure 7b) selon les
différentes expositions « Nord » ; « Sud » ; « Est » ; « Ouest » et au milieu des inter-rangs ; avec le témoin agricole « PAR calculé station ». Mesures effectuées le 11/05/13 pour AF2, et le 06/05/13 pour AF3.
Heure 2:00 5:00 8:00 11:0014:0017:0020:0023:00 0 500 1000 1500 2000 2500
Evolution journalière des PAR
Parcelle AF2 sous temps ensoleillé
Exposition Nord Milieu allée Exposition Sud PAR calculé station
P A R ( µ m o l/m ²/ s ) 0:00 3:00 6:00 9:00 12:00 15:00 18:00 21:00 0 500 1000 1500 2000 2500
Parcelle AF3 sous temps ensoleillé
Exposition Ouest Milieu allée Exposition Est PAR calculé station
P A R ( µ m o l/m ²/ s )
Figure 8a : Représentation du pourcentage de graines levées en conditions agroforestières sur des placettes de 1m² situées
en bordure (« 2m ») de la rangée d'arbre ou au milieu de l'inter-rang (« 6,5m »); en comparaison avec les témoins agricoles « TA » sur les parcelles A2 et A3.
Figure 8b : Représentation du pourcentage de graines levées en conditions agroforestières sur des placettes de 1m² selon
leur exposition : les bordures exposées « Nord », « Sud », « Ouest » et « Est » sont à comparer avec les placettes des milieux de l'inter-rang « Milieu » et aux témoins agricoles « TA » des parcelles A2 et A3.
Figure 8 : Représentation de l'influence des conditions agroforestières sur la germination de la graine de pois, selon la
Figure 9 : Représentation de la biomasse aérienne par plante de pois prélevée pendant la floraison, en condition
agroforestière en « Bordure » à 2m de la rangée d'arbres, et au milieu de l'inter-rang (« Milieu allée »), à comparer avec les témoins agricoles « TA » des parcelles A2 et A3.
Figure 10 : Représentation de l'indice foliaire moyen du pois cultivé en agroforesterie et prélevé au stade floraison en
« Bordure » à 2m de la rangée d'arbres, et au milieu de l'inter-rang (« Milieu allée »), à comparer avec les témoins agricoles « TA » des parcelles A2 et A3.
Figure 11 : Représentation du nombre moyen de nodules par plante en agroforesterie et prélevée au stade floraison en
« Bordure » à 2m de la rangée d'arbres, et au milieu de l'inter-rang (« Milieu allée »), à comparer avec les témoins agricoles « TA » des parcelles A2 et A3.
Figure 12a : Représentation du nombre de gousses matures et jeunes chez des pois cultivé en agroforesterie en stade fin de
floraison, en « Bordure » à 2m de la rangée d'arbres, et au milieu de l'inter-rang (« Milieu allée »), à comparer avec les témoins agricoles « TA » des parcelles A2 et A3.
Figure 12b : Représentation du nombre de gousses matures et jeunes chez des pois cultivé en agroforesterie en stade fin de
floraison, selon leur exposition : les mesures effectuées en bordures exposées « Nord », « Sud », « Ouest » et « Est » sont à comparer avec celles des milieux de l'inter-rang « Milieu » et aux témoins agricoles « TA » des parcelles A2 et A3.
Figure 12 : Représentation du nombre de gousses matures et jeunes chez des pois cultivé en agroforesterie en stade fin de
Figure 13 : Représentation du poids sec moyen de graines de pois, prélevées sur les gousses matures de plantes cultivées en
agroforesterie, au stade fin de floraison, selon différentes expositions : les bordures exposées « Nord », « Sud », « Ouest » et « Est » sont à comparer avec le milieu des l'inter-rangs « X » et aux témoins agricoles « TA » des parcelles A2 et A3.
Figure 14a : Évolution des biomasses aériennes de pois dans les deux parcelles A2 et A3.
Figure 14b : Évolution des biomasses aériennes de pois en fonction de l'exposition reçue par la culture : « Nord » ; « Sud » ;
« Est » ; « Ouest » ; avec « X » pour le milieu d 'allée et « TA » comme témoin agricole.
Figure 14 : Évolution des biomasses aériennes de pois récoltés le 26/04/13 (en bleu) et le 03/06/13 (en rouge), en fonction
de la parcelle (en haut) et en fonction de l'exposition (en bas) dans les parcelles A2 et A3.
A2 A3 0 1 2 3 4 5 6 7 8
Biomasse aérienne totale
le 26/04 et le 03/06 sur A2 et A3 Biomasse 26/04 Biomasse 03/06 B io m a s s e a é ri e n n e p a r p la n te ( g )
Marie RAIMBAULT, 2013.- Comportement et performances du pois en système agroforestier.
Master SPV 1ère année, mention BAS, Sciences et Productions Végétales, Agrocampus Ouest - Université de Rennes 1
UMR System – INRA, 2 place Viala, Bât 27, 34060 Montpellier Cedex 1 – France
RESUME
Cette expérimentation porte sur l'étude de la réponse du pois protéagineux (Pisum sativum L.) dans un système agroforestier sous climat méditerranéen. La station expérimentale date de 1995, et présente deux parcelles agroforestières de noyers hybrides, qui sont dans l'une alignés selon l'axe Nord-Sud et dans l'autre selon l'axe Est-Ouest. L'impact de la diminution de la lumière dans le système est évalué à l'aide de mesures de rayonnement global et de Rayonnement photosynthétiquement actif, en fonction de la distance à l'arbre (bordure ou milieu de l'allée) et de l'exposition. Le comportement du pois en agroforestrie est ensuite comparé à un témoin agricole. Le système, avant le débourrement des arbres, semble plutôt favorable au développement du pois, mais cette tendance s'inverse par la suite. Nous estimons la perte de rayonnement global à plus de 30% en système sylvo-arable, qui s'accompagne d'une diminution de rendement. L'hétérogénéité est renforcée au sein de la parcelle, l'exposition jouant sur la phénologie et la biomasse aérienne produite. C'est la parcelle orientée Nord-Sud qui est la plus affectée, montrant une plus grande hétérogénéité au niveau maturité et biomasse, et une chute de rendement de 36% par rapport au témoin.
Mots clés : Agroforesterie, Légumineuses, Rendement, Rayonnement
Marie RAIMBAULT, 2013.- Pea behaviour and performances in an agroforest system.
Master SPV 1ère année, mention BAS, Sciences et Productions Végétales, Agrocampus Ouest - Université de Rennes 1
UMR System – INRA, 2 place Viala, Bât 27, 34060 Montpellier Cedex 1 – France
ABSTRACT
This experiment aims to study the response of the proteaginous pea (Pisum sativum L.) in a agroforest system under mediterranean climate. The experimental station, created in 1995, displays two agroforest plots of hybrid walnut trees, with North-South and East-West oriented tree rows each. The impact of the light reduction is evaluated thanks to global radiation and Photosynthetic Active Radiations measurments, according to the tree distance (border or middle of the row) and the exposure. Pea performances in agroforestry are compared to a sole crop control. Before the trees budburst, the system seems to be favourable to pea development, but it trends to be the opposite after. We estimate that the global radiation decreases of about 30% in the agroforestry system, which comes with a yield decrease. The heterogeneity is strong in the field, and the exposure influences on the phenology and the biomass. The North-South orientated field is the most affected, showing a bigger heterogeneity than the other one, concerning the maturity and the biomass, and a yield loss of about 36% as compared to the control