f
Université de Montréal
Structure génétique et sérotinisme de populations
de pin gris (Pinus banksiana Lamb.)
soumises à deux régimes des feux distincts
par
Sylvie Gauthier
Département de sciences biologiques Faculté des arts et des sciences
Thèse présentée à la Faculté des études supérieures en vue de l'obtention du grade de
Philosophiae Doctor (Ph. D.) en sciences biologiques
Décembre 1991
Mise en garde
La bibliothèque du Cégep de l’Témiscamingue et de l’Université du Québec en Abitibi-Témiscamingue a obtenu l’autorisation de l’auteur de ce document afin de diffuser, dans un but non lucratif, une copie de son œuvre dans Depositum, site d’archives numériques, gratuit et accessible à tous.
L’auteur conserve néanmoins ses droits de propriété intellectuelle, dont son droit d’auteur, sur cette œuvre. Il est donc interdit de reproduire ou de publier en totalité ou en partie ce document sans l’autorisation de l’auteur.
Université de Montréal Faculté des études supérieures
Cette thèse intitulée:
"Structure génétique et sérotinisme de populations de pin gris
(Pinus banksiana Lamb.) soumises à deux régimes des feux
distincts" présentée par Sylvie Gauthier
a été évaluée par un jury composé des personnes suivantes:
André Bouchard (président-rapporteur) Jean-Pierre Simon (Directeur de recherche) Yves Bergeron (Co-directeur de recherche) Jean Bousquet (Examinateur externe) Pierre Legendre (Membre du jury)
i i i
SOMMAIRE
L'importance des feux de forêt, tant sur la dynamique des communautés végétales et des populations d'espèces que sur les adaptations des organismes à ces perturbations, est de plus en plus
reconnue. Toutefois, peu d'études ont tenté de démontrer l'effet
de différents régimes des feux, à 1' échelle régionale, sur des
caractères génétiques ou adaptatifs, au sein des populations d'une même espèce. Dans le cadre de cette étude, deux unités de paysage adjacentes ont été sélectionnées dans le sud de la forêt boréale du Québec, afin de déterminer l'effet de l'insularité et celui de deux régimes des feux distincts sur la structure génétique et sur le sérotinisme de populations de pin gris (Pinus banksiana Lamb.), à
une échelle régionale. L'unité de paysage insulaire, le lac
Duparquet (Abitibi), constituée de nombreuses îles, possède un régime de perturbations complexe comportant des feux d'intensité variable, qui affectent de faibles surfaces. L'unité terrestre, ad-jacente au lac, est affectée par un régime de feux destructeurs de grandes superficies, caractéristique de la forêt boréale.
Pour définir l'effet de l'insularité et des régimes des feux sur la structure et la variabilité génétiques, deux populations insulaires et deux populations terrestres ont été comparées au moyen de marqueurs isoenzymatiques. En moyenne, pour les 22 loci analysés, des taux de polymorphisme et d'hétérozygotie, respectivement de 60,0% et de 17,1%, ont été observés. La différenciation entre les
populations est faible (Fst
=
0,018) et aucune différencesignificative entre les populations pour les cinq paramètres de
variabilité génétique estimés n'est décelée. Le flux génique
important semble être responsable de la faible différenciation observée entre les populations des deux unités de paysage.
L'effet sélectif des régimes des feux différentiels a été évalué sur le sérotinisme des individus puisque la capacité des pins
gris à produire des cônes sérotineux est considérée comme une
adaptation majeure aux feux de forêts. Le sérotinisme de plusieurs centaines d'individus de pin gris a été déterminé dans vingt-cinq
iv populations localisées dans les deux unités de paysage. Dans un
premier temps, la taille et l'âge requis pour développer le
caractère sérotineux ont été déterminés. La majorité des individus ne possèdent pas de cônes sérotineux avant d'avoir atteint 7 cm de diamètre à hauteur de poitrine (DHP) et la proportion de cônes séro-tineux a tendance à augmenter annuellement jusqu'à ce que les arbres aient atteint une taille de 10 cm de DHP. La proportion de cônes sérotineux chez les cônes âgés de 4 années ou plus est inférieure d'environ 10% à celle des cônes plus jeunes. Cette proportion est également plus faible pour les cônes situés sur les branches inférieures à une hauteur de 2, 6 rn comparativement à celle des cônes sur les branches plus élevées. Ce même résultat est observé pour des cônes soumis à une température contrôlée de 41 °C , indiquant que l'environnement thermique naturel auquel les cônes sont soumis n'est pas le seul facteur impliqué dans leur ouverture sur les branches les plus basses. Les résultats permettent de suggérer que l'absence de cône sérotineux chez les pins gris de faible taille est liée à la juvénilité des individus.
La majorité des pins gris échantillonnés sont fortement séro-tineux. Cependant, environ 30% des arbres sont non sérotineux ou mixtes. Les individus non sérotineux et mixtes sont plus abondants lors d'établissement en absence de feu, au détriment des arbres sérotineux qui sont favorisés, quant à eux, lors d'établissement après un feu létal. L'âge de la forêt agit également sur la
répartition des individus dans les classes de sérotinisme à
l'intérieur des populations. Les individus non sérotineux sont
avantagés, d'une part, dans les forêts dont l'initiation est
relativement récente et, d'autre part, dans les forêts âgées de plus
de 200 ans. La répartition des arbres dans les classes de
sérotinisme diffère significativement entre les deux unités de paysage. L'intensité de la perturbation apparait être un facteur important, favorisant les individus sérotineux lorsque les feux sont létaux et avantageant les individus non sérotineux, lorsqu'ils sont non létaux. Les résultats suggèrent que les régimes des feux
v distincts ont exercé des effets sélectifs à long terme suffisants pour différencier les populations sur le caractère adaptatif qu'est le sérotinisme et ce, en dépit du flux génique important entre les populations.
TABLE DES MATIERES
SOMMAIRE
TABLE DES MATIERES
LISTE DES TABLEAUX
LISTE DES FIGURES
REMERCIEMENTS . . . . INTRODUCTION GENERALE . . . . . 1. LE PROCESSUS EVOLUTIF 1.1. Mutation . . . . 1.2. Dérive génétique 1. 3. Migration 1.4. Sélection naturelle
2. LES PERTURBATIONS RECURRENTES
SELECTIVE . COMME PRESSION vi i i i vi xii xvi xx 1 2 3 4 5 6 9 3. CONTEXTE DE L 1 ETUDE ET OBJECTIFS GENERAUX . . . . . 11
3 .1. Effets de l 1
insularité et des régimes
3.2 .
des feux sur la structure génétique Effets sélectifs des deux régimes des
feux .
4. BIOLOGIE DE Pinus banksiana Lamb.
5. DESCRIPTION DU TERRITOIRE . . . . .
5.1. Physiographie et végétation
5.2. Régimes des feux . . .
13 1 7 21 24 24 2 7
TABLE DES MATIERES (suite).
CHAPITRE I. Effets de l'insularité et des régimes des feux sur la variabilité et la structure génétiques de populations
vii
de pin gris (Pinus banksiana Lamb. . . . 31
1. INTRODUCTION 2. MATERIEL ET METHODES 2 .1. 2.2. 2. 3. Echantillonnage . . . . Analyses électrophorétiques Analyses statistiques 2. 3 .1. 2. 3.2. Ségrégation Variabilité génétiques et structure 3. RESULTATS 3 .1. 3. 2. 3. 3. 3. 4. 3. 5. Ségrégation . . Structure d'âge
Fréquences alléliques et variabilité des populations . .
Structure génétique des populations Différenciation . . . .
3. 5 .1. 3. 5. 2.
Toutes les populations
A 1' intérieur des unités de
paysage
3. 5. 3. Entre les unités de paysage
4. DISCUSSION
. . .
. . . . .
.
.
.
. . . . .
4 .1. Ségrégation.
. .
.
.
.
. . . .
4.2. Caractéristiques génétiques générales
4. 3. Structure génétique et différenciation
des populations
.
.
. . . .
5. CONCLUSION. .
. . .
. . .
.
. . . .
6. BIBLIOGRAPHIE.
.
.
32 36 36 36 40 40 41 44 44 49 49 58 60 60 68 68 70 70 70 73 76 77TABLE DES MATIERES (suite).
CHAPITRE II. Effets de la position des cônes et de l'âge des
individus sur le sérotinisme du pin gris (Pinus
viii
banksiana Lamb.) . . . 86
1. INTRODUCTION 87
2. MATERIEL ET METHODE . . . 89
2 .1. Echantillonnage . . . 89
2.1.1. Origine des individus et
évalua-tion du degré de sérotinisme des individus sur le terrain
2 .1.2. Evaluation du degré de sérotinisme
89
des individus en laboratoire 90
2.2. Analyse des données . . . 90
2.2.1. Taille ou âge requis pour
2.2.2.
2. 2. 3.
développer le sérotinisme
Taille ou âge requis pour le
dévelbppement complet du sérotinisme Effets et/ou de de l'année la de maturité hauteur sur
l'ouverture des cônes 3. RESULTATS
3 .1. Taille ou âge requis pour développer le
3.2.
3. 3.
sérotinisme
Taille ou âge requis pour le
développement complet du sérotinisme Effets de l'année de maturité et/ou de
90 92 93 95 95 98
la hauteur sur l'ouverture des cônes 98
4. DISCUSSION 106
5. CONCLUSION 110
TABLE DES MATIERES (sui te) .
CHAPITRE
:r:r:r.
Effets de deux régimes des feux distincts sur le sérotinisme du pin gris (Pinus banksiana Lamb.)1. INTRODUCTION
2. REGIME DES FEUX DES DEUX UNITES DE PAYSAGE 3. MATERIEL ET METHODES . . . .
3.1. Echantillonnage
3.2. Analyses statistiques
3.2.1. Analyses préliminaires
3 . 2 . 2. Comparaison de la répartition des
individus dans les classes de
sérotinisme selon le type
d'établissement
3 .2 .3 . Hétérogénéité entre les stations
3.2.4. Modèles log- linéaires . . . .
4. RESULTATS 4 .1.
3. 2. 4 .1. Rappel théorique . . . .
3.2.4.2. Utilisation des modèles
Analyses préliminaires . . . .
4 .1.1. Comparaison de la distribution des
DHP des individus selon la classe
de sérotinisme et l'unité de
paysage
4 .1. 2. Comparaison des deux ensembles de
ix 115 116 120 12 6 126 129 12 9 130 131 132 133 1 34 1 3 6 136 1 36 données . . . 13 8
4. 2. Effet du type d'établissement sur la
répartition des individus dans les
TABLE DES MATIERES (suite).
4. 3. Variabilité dans le sérotinisme entre
les stations
4.3.1. Globalement
x
143 143
4.3.2. Unité de paysage terrestre 150
4.3.3. Unité de paysage insulaire 150
4.4. Effet de la période d'initiation sur le degré de sérotinisme des individus selon leur type
d'établissement par unité de paysage 162
4.4.1. Unité de paysage terrestre 163
4.4.2. Unité de paysage insulaire 167
4. 5. 5. DISCUSSION 5 .1. 5.2. 5. 3. 5. 4. 5. 5. 6. CONCLUSION 4.4.2.1. Individus provenant de
stations sans feu non létal après l'initiation . . .
4. 4.2 .2. Individus provenant de
stations avec feux non
létaux après l'initiation
Comparaisons de la répartition des
individus dans les classes de
sérotinisme entre les unités de
paysage
Effet du type d'établissement sur le sérotinisme des individus . .
Effet de la période d'initiation sur le sérotinisme des individus .
Effet de l'occurrence de feux non
létaux sur le sérotinisme des individus des populations des deux unités de paysage . . . .
Effet des régimes des feux
Conséquences des différences de
sérotinisme entre les unités de paysage sur la dynamique des populations
167 167 170 180 180 183 184 186 187 189
TABLE DES MATIERES (suite). 7. BIBLIOGRAPHIE
CONCLUSION GENERALE . .
BIBLIOGRAPHIE (INTRODUCTION ET CONCLUSION GENERALES)
xi
191
197
xii
LISTE DES TABLEAUX
CHAPITRE I
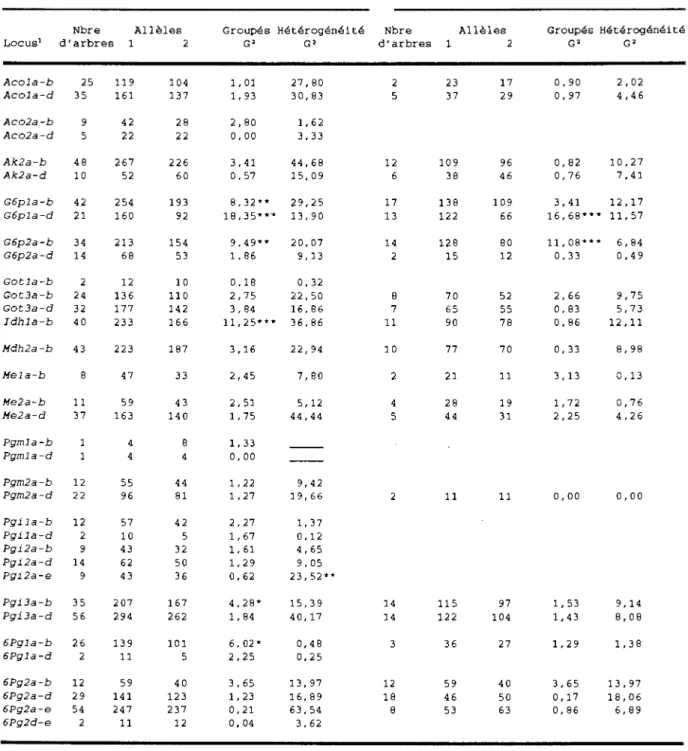
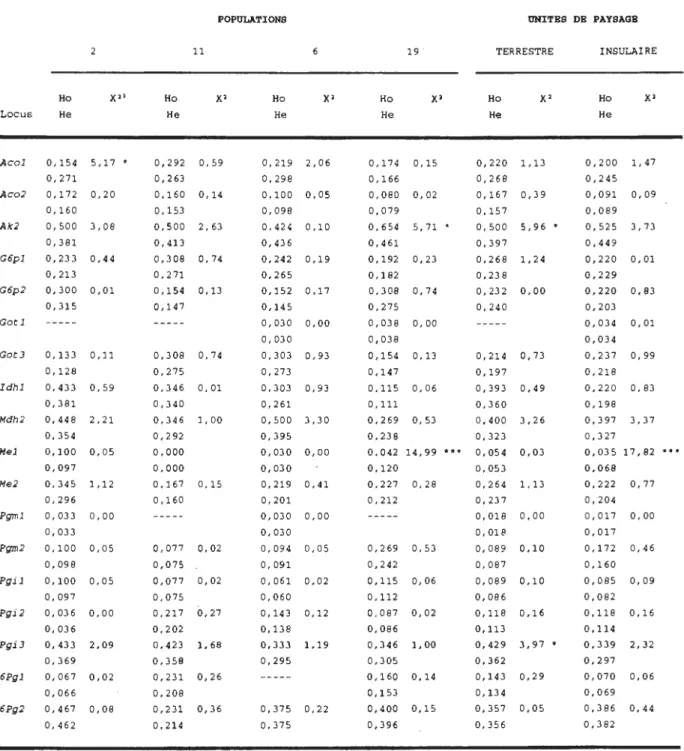
Tableau I. Caractéristiques des quatre populations de
pin gris étudiées 39
Tableau II. Rapports de ségrégation des alloenzymes
observés pour tous les pins gris
hétérozygotes groupés, et pour ceux analysés
avec 10 mégagamétophytes et plus . . . 45
Tableau III. Tableau IV. Tableau V. Tableau VI. Tableau VII . Tableau VIII. Tableau IX.
Fréquences alléliques et nombre effectif d'allèles par population et par unité de paysage Paramètres génétique gris estimés des quatre de la variabilité populations de pin
Khi-carré testant 1 'égalité entre les taux d'hétérozygotie observée (Ho) et espérée (He)
selon l'équilibre de Hardy-Weinberg par
locus, par population et par unité de
paysage
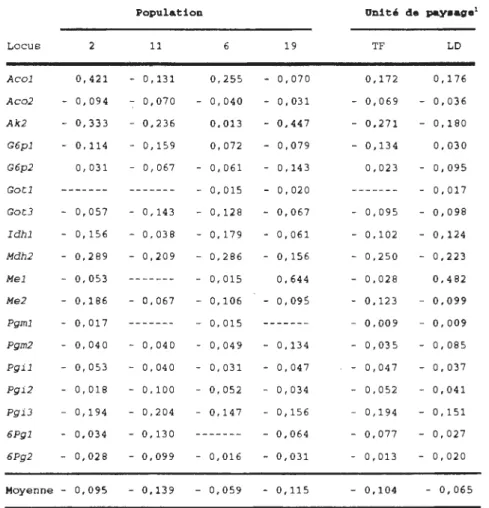
Valeurs estimées de Fis par population et par unité de paysage pour les 22 loci analysés
Valeurs estimées des statistiques de F (F1s ,
Fit et F st) selon les trois modèles de
répartition des populations
Estimation probabilité
par test de khi-carré de la
d'homogénéité des fréquences
allélique s par locus
Distances géographiques en kilomètres et
distances génétiques non-biaisées de Nei (1978) entre les quatre populations
52 55 57 59 61 64 67
xiii
LISTE DES TABLEAUX (suite)
Tableau I. Tableau II. Tableau III. Tableau IV. Tableau V. Tableau VI. Tableau VII. Tableau I. Tableau II. CHAPITRE II Tests de khi-carré
d'incertitude pour les classes des variables selon la présence ou fermés et coefficients partitions en deux DHP, âge et hauteur l'absence de cônes
Tableau de contingence de la présence-absence de cônes fermés selon 6 classes de DHP
Relation entre l'âge, le DHP ou la hauteur et les indices de croissance de la proportion de cônes fermés dans le temps . . . . . Test de Friedman apparié par individu et comparaisons multiples des proportions de cônes fermés par année de maturité
Comparaison appariée par individu de Wilcoxon des proportions de cônes fermés entre les cônes de 1 an et les cônes plus vieux
Comparaison appariée par individu de Wilcoxon des proportions de cônes fermés par classes de hauteur de branche pour les cônes témoins
96
97
99
100
102
et pour les cônes de 2 ans et plus 103
Analyses de rang multidimensionnelles de
l'effet de la hauteur des branches et de
l'année de maturité des cônes sur la
proportion de cônes fermés 105
CHAPITRE III
Synthèse des différences entre les régimes
des feux des deux unités de paysage . . . . 125 Description des populations échantillonnées
xiv
LISTE DES TABLEAUX (suite)
Tableau III. Tableau IV. Tableau V. Tableau VI. Tableau VII. Tableau VIII. Tableau IX. Tableau X.
Analyse de variance à 2 critères sur le DHP des individus
Comparaison des fréquences d'individus dans les classes de sérotinisme des deux ensembles
de données globalement et par unité de
paysage
Effectifs, diamètre à hauteur de poitrine moyen et pourcentage de cônes fermés moyen
dans les 4 types d'établissement, par
station, par période d'initiation et par
137 141 uni té de paysage . . . . 142 Hétérogénéité entre d'établissement dans les la différents types répartition des
individus dans les classes de sérotinisme: 144
Hétérogénéité entre les stations dans la répartition des individus dans les classes de sérotinisme, par unité de paysage et entre les unités de paysage . . . .
Hétérogénéité entre les stations dans la
répartition des individus dans les classes de
sérotinisme, par période d'initiation et
entre les périodes d'initiation pour l'unité de paysage terrestre
Hétérogénéité entre les stations dans la répartition des individus dans les classes de
sérotinisme, pour l'unité de paysage
insulaire
Modèles log-linéaires pour l'unité insulaire
entre les classes de sérotinisrne, les
périodes d'initiation et l'occurrence ou non d'un feu non létal depuis l'initiation
147
153
154
LISTE DES TABLEAUX (suite) Tableau XI. Tableau XII. Tableau XIII. Tableau XIV. Tableau XV.
Modèles log-linéaires entre la classe de sérotinisme, la période d'initiation et le
type d'établissement des individus pour
l'unité terrestre
Modèles log-linéaires entre la classe de sérotinisme, la période d'initiation et le type d'établissement des individus pour le
groupe d' individus
stations sans feu
l'initiation
insulaires issus de
non létal depuis
Modèles log-linéaires entre la classe de sérotinisme, la période d'initiation et le type d'établissement des individus pour le
groupe d'individus insulaires issus de
stations avec feux non létaux depuis
l'initiation
Hiérarchie de modèles log-linéaires à 4
dimensions entre la classe de sérotinisme, la période d'initiation, le type d'établissement et le groupe d'individus
Paramètres calculés et écart-types sous le modèle 4 pour les différents effets simples et les différentes interactions
xv 166 168 169 172 174
xvi LISTE DES FIGURES
Figure 1. Figure 1. Figure 2. Figure 1. Figure 2. Figure 3. Figure 4. Figure 5. Figure 6. INTRODUCTION GENERALE
Carte de localisation du territoire et des 25 populations de pins gris étudiées
CHAPITRE I
Carte de localisation du territoire et des quatre populations de pin gris étudiées Structures terrestres insulaires d'âge et des des CHAPITRE III deux deux populations populations
Carte de localisation du territoire et des vingt-cinq populations de pin gris étudiées Comparaison des fréquences · relatives d'individus dans les classes de sérotinisme entre les deux ensembles de données,
25
37
50
121
globalement et par unité de paysage 139 Comparaison des fréquences relatives
d'individus par classe de sérotinisme entre
les quatre types d'établissement . . . 145 Comparaison des fréquences relatives
d'individus par classe de sérotinisme entre
les deux unités de paysages . . . 148 Comparaison des fréquences relatives
d'individus dans les classes de sérotinisme entre les trois périodes d'initiation pour
l'unité de paysage terrestre 151 Comparaison des fréquences relatives
d'individus dans les classes de sérotinisme
LISTE DES FIGURES (suite)
Figure 7. Comparaison des fréquences relatives
d'individus dans les classes de sérotinisme pour l'unité de paysage insulaire par période d'initiation selon l'occurrence de feux non
xvii
létaux 160
Figure 8.
Figure 9.
Comparaison des fréquences
d'individus dans les classes de par type d'établissement et
relatives sérotinisme par période d'initiation selon l'unité de paysage
Paramètres standardisés estimés par modèle
log-linéaire montrant les excès ou les
déficiences d'individus dans les cinq classes de sérotinisme entre les deux unités de paysage
164
"Nothing in biology makes sense except in the light of evolution."
Dobzhans ky
" ce qui justifie l'existence du Chercheur inconnu, ce n'est pas la découverte, qui est l'éclair inattendu d'un millième de seconde, mais le cheminement patient et passionné, qui trouve en lui-même sa récompense."
xx REMERCIEMENTS
Ma reconnaissance et mes remerciements vont, en tout premier lieu, au Dr Jean-Pierre Simon, mon directeur, pour son réalisme et son honnêteté, ainsi qu'au Dr Yves Bergeron, mon co-directeur, pour son optimisme, son intuition et, particulièrement, pour rn' avoir fait découvrir le lac Duparquet. La confiance, les encouragements, le support financier, technique et scientifique, les critiques et les conseils judicieux qu'ils m'ont apportés tout au long de ce projet, m'ont été des plus précieux. Qu'ils sachent l'amitié et le respect que je leur porte.
Mille mercis à mes amis et amies, qui, comme moi, ont été Abitibiens d'adoption, dont le sens du partage et de la concession ont rendu l'été 1987 très agréable. Maryse Rochefort, ma p'tite ta ... , pour le répertoire de chansons des années '60 ... ; Sylvain Archambault, pour ses yeux qui brillaient le soir en comptant ses galet tes de vieux cèdres et son inépuisable énergie ... ; Pierre Dansereau, le faux!!!, pour sa naïveté et les belles ballades en canot sur la Magusi ... , Yvan Belouin, pour sa gaieté et son entrain ... ; Danielle Lalonde, pour son énergie et Richard, pour son calme imperturbable. Nos nombreux échanges ont été une source de motivation.
A tous les membres du laboratoire du Dr Jean-Pierre Simon, des remerciements sincères: à Lise Gobeille, pour notre amitié, nos réf lex ions et ses nombreux encouragements; à Jean Beaulieu, pour nos discussions et ses commentaires sur différents manuscrits; à Benoit Gagnon, Sieu Ngo, Yves Fayette, Odette Préfontaine, pour leur efficacité sur le terrain comme en laboratoire; à Josée Prud'homme, Nadia Hakam et Christiane Charest, pour leur présence chaleureuse.
A tous les membres du Groupe de recherche en écologie forestière, un grand merci, pour les idées et questions surgies lors des réunions: à Julie Gagnon, pour m'avoir initié aux mouches noires et aux maringouins de l'Abitibi et pour la permission d'utiliser certains de ses quadrats; à Danielle Charron, François Tétrault et
xxi
Jacques Brisson, pour le sablage et la lecture des galettes; à
Jacques Tardif, Ting Xian Li, Pierre Cartier, Brian Harvey, Marie-Claude Rousseau, Marie-Marie-Claude Brisson et Janet Wyman pour leur énergie et leur amitié.
Il me fait plaisir de remercier tous ceux et celles qui ont
participé à la réalisation de ce projet. Pour nos nombreuses
discussions devant un tableau noir, cigarette, café et craie en mains, pour ses encouragements et sa présence dans les moments les
plus difficiles, un merci particulier à Alain Leduc, mon ami et
conseiller; un merci bien spécial à Andrée Nault, pour ses
encouragements, sa confiance et son aide; des mercis sincères vont à Valérie Bolliet, Pierre Drapeau, Alain Gauthier, Henri-Paul Gauthier, Mario Gauthier, Réjeanne Périard-Gauthier, Vincent Jarry et Chantal Ménard, sans la présence, le réconfort et le support de qui, je n'aurais pu mener ce travail à terme.
Les suggestions, corrections et commentaires apportés par Messieurs André Bouchard, Pierre Legendre et Jean Bousquet ont grandement contribué à améliorer la qualité de ce travail. Je les en remercie profondément.
La courtoisie et l'efficacité du personnel de la bibliothèque de Biologie de l'Université de Montréal, ainsi que du personnel des départements de sciences biologiques de l'Université de Montréal et
de l'Université du Québec à Montréal, méritent d'être soulignées.
Des remerciements particuliers sont adressés à Henriette Gagnon qui a réalisé les cartes.
Je tiens finalement à remercier le CRSNG et l'Université de Montréal pour les bourses qu'ils m'ont accordées. Les subventions de recherche, attribuées à Jean-Pierre Simon (CRSNG et FCAR-équipe) et à Yves Bergeron (FCAR-équipe), ont également grandement contribué à la réalisation de ce projet.
1. LE PROCESSUS EVOLUTIF
Les mécanismes en jeu dans l'hérédité des caractères, de même que ceux impliqués dans le processus de l'évolution, demeuraient imprécis jusqu'à tout récemment. L'élaboration de la théorie de la sélection naturelle par Darwin en 1859 et la redécouverte, par de Vries en 1900, des lois de Mendel définies en 1866, constituent deux éléments clés ayant mené la communauté scientifique à une meilleure compréhension du processus évolutif. De fait, une synthèse de la théorie évolutive émergeait vers 1950 grâce aux travaux de Huxley (1942), Dobzhansky (1937), Stebbins (1950), Mayr (1942) et Simpson (1944), pour n'en citer que quelques-uns, créant un consensus entre les écoles de pensée qui s'opposaient jusqu'alors, principalement celle de génétique et celle de sciences naturelles et de systéma-tique (Mayr et Provine 1980).
L'existence de variations entre individus d'une même espèce (polymorphisme) au sein de caractères, qui apparaît d'ailleurs être généralisée à la grande majorité des espèces (Selander 1976; Ayala 1982; Lewontin 1985), de même que la possibilité que ces caractères soient transmis à la progéniture (hérédité), constituent le point de départ des changements évolutifs. Ainsi, chaque individu est muni d'un bagage génétique, son génotype, qui, en interaction avec l'environnement dans lequel l'organisme évolue, résulte en un phéno-type particulier. Les gènes d'un individu, lors de la méiose, sont assemblés en nombreux gamètes mâles et /ou femelles, tous rendus différents par la recombinaison génétique. Chez des organismes sexués tels les conifères, les croisements entre individus engen-drent un nouvel ensemble de génotypes. Les individus qui vivent dans un espace donné, lorsqu'ils ont la possibilité de se croiser aléatoirement, forment une population, unité d'intérêt en biologie évolutive. Cette population a un pool génique particulier. Pour une population théorique idéale (i.e. une population d'organismes diploïdes à reproduction sexuée, de taille infinie, où les générations sont discrètes et où les individus se croisent de façon aléatoire), les fréquences alléliques et génotypiques demeurent stables d'une génération à l'autre. Dans cette situation de stabilité, connue sous le nom d'équilibre de Hardy-Weinberg, il n'y
3
a pas de changements évolutifs. De fait, l'évolution peut être définie comme un changement cumulatif dans la constitution génétique d'une population, de génération en génération (Spurr et Barnes 1980). En d'autres termes, l'évolution est un changement dans les
fréquences alléliques des populations, cumulé d'une génération à
l'autre (Spiess 1977). Quatre forces évolutives majeures sont reconnues pour leurs effets sur les fréquences alléliques des populations naturelles: les mutations, la dérive génétique, la migration et la sélection naturelle. Chacune de ces forces sera décrite succinctement dans les prochaines sections.
1.1. Mutation
Chez les conifères, comme chez les autres organismes vivants, les variations entre les phénotypes des individus sont dues tant à leurs constitutions génétiques différentes qu'à l'effet de l'envi-ronnement dans lequel ils évoluent. Toutes modifications dans la séquence de nucléotides ou toutes altérations du génome sont appelées mutations. Elles constituent la source ultime de variation génétique. Ces changements dans la structure moléculaire à un locus particulier ont pour effet d'introduire de nouveaux allèles, dispo-nibles pour la recombinaison dans les populations. Le résultat des mutations, lorsque celles-ci sont viables, est donc d'introduire de
la variabilité dans le pool génique d'une population. Pour des
organismes tels les plantes, qui ont en grande majorité une
reproduction sexuée, les croisements donnent lieu à une quasi
infinie possibilité de nouveaux génotypes, constituant par la suite, le matériel brut sur lequel les autres forces évolutives vont agir.
4
1.2. Dérive génétique
Le terme dérive génétique réfère aux fluctuations aléatoires dans les fréquences alléliques. Peu de populations naturelles cor-respondent parfaitement à la population théorique idéale, définie précédemment. Ainsi, peu de populations se comportent comme si elles avaient une taille infinie, puisque les individus ont en général plus de chance de se croiser avec leurs proches voisins (Levin 1988). Une population où la répartition des individus est apparem-ment continue, est souvent subdivisée en plus petites unités où les individus se croisent effectivement de façon aléatoire (unité
panmictique, breeding units), les sous-populations (Wright 1978b) .
Le nombre d'individus qui forme une sous-population insulaire, par exemple, peut être limité. Lorsque la taille d'une population est réduite, le pool génique est également réduit, constituant alors un sous-échantillon aléatoire plus ou moins représentatif du pool génique de la population globale, disons par rapport à celui de l'espèce. De plus, ce processus aléatoire d'échantillonnage des gamètes se répète dans le temps, de sorte qu'à chaque génération, on peut obtenir simplement par chance, un sous-échantillon génétique
plus ou moins différent du pool génique global. Le processus de
dérive, de génération en génération, peut être vu comme une
fluctuation aléatoire dans les fréquences alléliques conduisant, à la longue, à la fixation complète ou à l'élimination complète d'un allèle. Ce processus se produit dans toute population ayant une taille finie et sera d'autant plus rapide que 1 'effectif de la population est faible (Futuyma 1986). La taille des populations peut également varier dans le temps et dans l'espace, sous l'effet d'une catastrophe, par exemple, qui réduira considérablement l'effectif de la population. L'établissement d'une nouvelle population à partir de peu d'individus affecte également le pool génique. Tous ces
facteurs font en sorte que les fréquences alléliques d'une
population peuvent fluctuer dans le temps et/ou dans l'espace, de
façon purement aléatoire. C'est le pool particulier d'une
population qui sera soumis, par la suite, aux pressions sélectives. La dérive génétique a deux conséquences évolutives principales.
5
variabilité génétique intra-population (Futuyma 1986). D'autre part, ces fluctuations du pool génique des populations dans le temps ou dans l'espace favorisent la différenciation des populations, particulièrement lorsque les effectifs sont réduits (Dobzhansky et Pavlovsky 1957).
1.3. Migration
La migration, en d'autres termes le flux génique, a un effet homogénéisateur entre les pools géniques observés dans différentes populations, en permettant le maintien dans chacune d'elles des différents allèles de la population globale. De plus, le flux génique constitue une source de variation importante dans le pool génique d'une population particulière, en introduisant de nouveaux allèles, auparavant absents de cette population. Un flux génique efficace entre les populations aura donc un effet homogénéisateur sur les fréquences alléliques. Ainsi, en absence de sélection, une migration aussi faible que 1 migrant par 2 générations sera suffi-sante pour empêcher la fixation d'un allèle due simplement à la dérive génétique (Slatkin 1985; Wright 1978a). Par contre, si le flux génique est restreint, (i.e. moins de 1 migrant par généra-tion) , plusieurs populations verront certains allèles devenir fixés sous l'effet de la dérive. Par ailleurs, même quand le flux génique est supérieur à 1 migrant par génération, les effets de la dérive peuvent se faire sentir, bien qu'ils ne conduisent pas à la fixation ou à l'élimination de certains allèles (Wright 1978a). De façon générale, on considère que le flux génique chez les conifères, tant par le pollen que par les graines, est relativement élevé,
particu-lièrement parce que la pollinisation est assurée par le vent et que le pollen peut voyager sur de grandes distances (Lanner 1966; Koski 1970; Loveless et Hamrick 1984; Hamrick et Godt 1989). Toutefois, il peut exister des barrières physiques, génétiques ou biologiques
à la migration. Par exemple, la phénologie des individus d'une population peut être différente de celle d'une autre, rendant difficile la pollinisation à partir de pollen migrant. De même, la distance géographique entre des populations peut être une barrière
6 reproductive varie selon l'individu, certains produisant davantage de gamètes femelles, d'autres davantage de pollen, tandis que d'autres sont momentanément stériles. Cette situation réduit le nombre d'arbres qui peuvent se croiser dans une population et par conséquent, le flux génique.
1.4. Sélection naturelle
Pour que le processus de sélection naturelle puisse opérer au sein d'une population, trois conditions sont requises: 1) des varia-tions entre les individus au niveau de certaines caractéristiques
ou attributs doivent exister; 2) ces variations doivent être
transmises à la descendance de façon héréditaire; et 3) la variation au sein de ces caractères doit affecter les capacités de survie et
de reproduction (fitness differences) des individus qui en sont
munis (Endler 1986) . Parmi les milliers de graines produites (donc des milliers de génotypes différents) par les individus d'une population à un endroit donné, très peu survivront jusqu'à maturité pour arriver à contribuer à la prochaine génération. La compétition entre individus, la disponibilité de la lumière, le milieu où la graine a germé sont quelques-uns des facteurs de l'habitat qui peuvent causer la mortalité de certains individus tout en permettant la survie et la reproduction de certains autres, mieux adaptés à ces conditions. On peut définir la sélection naturelle comme la survie et la reproduction différentielle et non aléatoire des individus, sous l'effet de leur niveau d'adaptation aux conditions environ-nementales (tant biotiques qu'abiotiques), permettant à certains génotypes de gagner ou de perdre en représentativité d'une génération à l'autre. Puisqu'une partie des caractères individuels sont transmis à la prochaine génération grâce à l'hérédité, la descendance laissée par les individus les plus aptes face aux
conditions environnementales rencontrées, ressemble, jusqu'à un
certain point, à la génération parentale. Il est opportun de
rappeler que la sélection naturelle agit sur le phénotype global des individus et non pas sur leur génotype. L'effet de la sélection naturelle se situe, au sein de la population, par un changement dans
7
le pool génique, puisque les individus les plus aptes de la génération précédente auront laissé plus de descendants que ceux qui 1 'étaient moins. Les individus d'une génération particulière peuvent donc être considérés comme adaptés aux conditions qui prévalaient pour les générations précédentes. Begon, Harper et Townsend (1990)
l'ont énoncé clairement:
Past environments act as a filter through which
cornbinations of characters have passed on their way to the present. But organisms appear to be adapted (fitted)
to their present environment only because present
environrnents tend to be similar to past environrnents. The word 'adaptation' gives an erroneous impression of prediction, forethought or, at the very least, design. Organisms are not designated for, or adapted to, the present or the future -- they are consequences of, and therefore adapted by, their past."
Le terme adaptation, utilisé abondamment dans la littérature
écologique, se définit donc comme la capacité des individus à
exploiter un environnement particulier avec succès, en laissant des
descendants. Cette adaptation des individus à un environnement se
répercute sur la composition génétique des populations d'une espèce. Des populations qui évoluent dans des conditions environnementales
différentes auront tendance à diverger en termes génétiques, sous
l'effet différentiel de la sélection naturelle, qui augmente
l'adaptation des individus à leur environnement. Toutefois, des
populations soumises à des conditions environnementales similaires,
auront tendance à se ressembler.
En résumé, ce sont les interactions entre les quatre forces évolutives, ayant parfois des effets contrastants, qui affectent le
cours de l'évolution. Ainsi, le processus de dérive génétique
tendra à augmenter la différenciation entre les populations et ce, d'autant plus que les populations ont de faibles effectifs , tandis qu'une migration efficace entre les populations aura pour effet de les homogénéiser. En conditions environnementales similaires, la sélection naturelle ralentira la différenciation entre les popula-tions alors qu'elle la favorisera lorsque les condipopula-tions sont distinctes. La variation génétique d'une espèce, résultant des mutations, sera donc maintenue, éliminée, organisée et/ou distribuée
8
à l'intérieur et entre les populations selon l ' équi l ibre et les interactions complexes entre la sélection naturelle, la migration et la dérive génétique. Comme le dit si clairement Speiss (1 977):
"Evolution results when one force or another gains an edge over others, when the balance is perturbated and
when directional change in gene frequencies at a
2. LES PERTURBATIONS RECURRENTES COMME PRESSION SELECTIVE
Harper ( 1977) a introduit la distinction entre ce qu '.i l appelle un désastre et une catastrophe. Ainsi, les désastres sont des perturbations naturelles récurrentes et suffisamment fréquentes, qu'elles sont susceptibles de se produire pendant le cycle vital des
individus de générations successives d'une espèce, constituant alors
une pression sélective. Les catastrophes, toutefois, ont une
occurrence tellement faible et imprévisible qu'il est peu probable qu'elles aient exercé des pressions sélectives sur les individus des populations d'une espèce. Par contre, elles peuvent avoir entraîné
une réduction considérable de l'effectif de la population
(population bottleneck), phénomène qui peut également avoir un effet
évolutif. Les perturbations naturelles et récurrentes1
, telles que
les feux de forêts peuvent, pour des espèces comme que le pin gris, être considérées comme des désastres. La variabilité rencontrée au sein de certaines des caractéristiques biologiques ("life-history
characteristics") tant au niveau intra- qu'inter spécifique,
apparaît être le résultat, du moins en partie, des pressions sélectives exercées par la récurrence de perturbations (Hendrickson 1972; Harper 1977; White 1979; Gill 1981; sousa 1984). De plus, une partie importante de l'hétérogénéité spatiale et temporelle observée dans la structure et la dynamique des communautés est la résultante des perturbations naturelles (West et al. 1981; Pickett et White
1985; Poster 1988). Ces deux rôles des perturbations récurrentes
1 Pickett et White (1985) font la distinction entre les termes
anglais perturbation et disturbance. Ils considèrent que le terme
perturbation devrait être utilisé dans un sens relativement
restreint. Il s'agit d'un écart, défini explicitement, à un état,
un comportement et/ou une trajectoire préalablement établis comme
étant normaux. Cet écart peut être dû
à
l'introduction d'un nouveautype de perturbation dans le système (perturbation anthropique) ou
encore
à
l'effet des manipulations d'un expérimentateur. Ladéfinition plus large de disturbance inclut
à
la fois lesfluctuations environnementales et les événements destructeurs. Il s'agit d'un événement relativement discret dans le temps qui affecte la structure d'un écosystème, d'une communauté ou d'une
population en changeant la disponibilité des ressources, des
substrats et/ou de l'environnement physique. La langue française ne permet pas cette distinction. Ainsi dans le présent texte, le mot
10 sont importants et interdépendants, en termes évolutifs. Des régimes de perturbations distincts ont exercé et exercent des pressions différentielles sur les caractéristiques des espèces, tout en résultant en des conditions environnementales et des assemblages différents d'espèces. Cette hétérogénéité, tant dans les conditions biotiques qu'abiotiques, constitue également un élément sélectif de l'habitat sur les caractéristiques biologiques des espèces (Sousa 1984).
Les feux, par exemple, ont été des perturbations naturelles suffisamment importantes et ' répandues, pour que les espèces végétales aient développé des mécanismes et adaptations qui leur ont permis, et leur permettent de faire face à des régimes des feux
par-ticuliers (Hendrickson 1972; Gill 1981; Pickett et White 1985). A l'échelle des populations, on peut également s'attendre à ce que des régimes des feux distincts aient contribué à différencier les populations d'une même espèce, au niveau de caractéristiques morphologiques, génotypiques ou biologiques (Keeley 1981; Jelinski
3. CONTEXTE DE L'ETUDE ET OBJECTIFS GENERAUX
En forêt boréale, les feux sont des perturbations naturelles et récurrentes, responsables, en grande partie, du dynamisme des communautés végétales (Rowe 1961). Les diverses composantes du régime des feux (l'occurrence, l'intensité, la sévérité et le cycle des feux) sont des éléments moteurs des différences perceptibles dans la mosaïque de ces communautés (Heinselman 1973, 1981a, 1981b; Wein et McClean 1983). Bien que le régime des feux soit essentielle-ment contrôlé par le climat régional, la configuration du paysage par l'intermédiaire du relief et de la présence de plans d'eau, constitue un élément de contrôle sur sa dynamique (Frissell 1973; Heinselman 1981a; Foster 1983; Grimm 1984; Payette et al. 1989). Les travaux récents de Bergeron et collaborateurs dans le sud de la forêt boréale québécoise ont mis en évidence 1 'existence d, un régime . des feux particulier, relativement complexe, dans une unité de paysage morcelée, les îles et les péninsules · du lac Duparquet (Bergeron et Gagnon 1987; Bergeron et Brisson 1990; Bergeron 1991). En effet, les îles et les péninsules de ce lac sont affectées d'un régime des feux caractérisé par une occurrence élevée de feux d'intensité variable, et qui incendient des surfaces relativement faibles (Bergeron 1991). Par contre, la forêt terrestre adjacente est affectée par le régime des feux typique de la forêt boréale: les feux y sont intenses, leur occurrence est relativement faible mais ils incendient de grandes surfaces (Heinselman 1981a; Van Wagner 1983; Dansereau 1991; Bergeron 1991).
Le pin gris, espèce sérotineuse, est en étroite relation avec le régime des feux, se régénérant presqu'exclusivement après feu (Eyre et LeBarron 1944; Cayford et McRae 1983). Des études ont montré que le feu est le principal facteur contrôlant sa distri-bution dans le secteur à l'étude (Gagnon 1990; Gauthier, Gagnon et Bergeron, données non publiées). Sa capacité à produire des cônes sérotineux est considérée comme son adaptation majeure à la récur-rence des feux (Cayford et McRae 1983) . Chez cette espèce, on retrouve des individus sérotineux, non sérotineux et mixtes (Rudolph et al. 1959; Schoenike 1976) . Ce polymorphisme, au sein d'un caractère adaptatif, pourrait permettre à un effet sélectif de
12
l'environnement de se manifester (Rudolph et al. 1959; McMaster et Zedler 1981; Muir et Lotan 1985). Ainsi, des populations de pins gris
(Pinus banksiana
Lamb.) sont susceptibles d'être sensibles àdes régimes des feux différentiels et de démontrer des différences dans un caractère adaptatif tel que le sérotinisme.
La région du lac Duparquet nous offre donc la possibilité de tester l'effet sélectif différentiel de deux régimes des feux, à une échelle régionale, sur certaines caractéristiques des populations d'une espèce arborescente forestière. Puisque la configuration du paysage du lac est morcelée comparativement à celle de 1 'unité terrestre adjacente, il est également possible d'étudier l'effet de l'insularité, à une échelle régionale, sur la structure génétique de populations d'arbres. Les populations insulaires sont constituées de plus petits nombres d'individus que les populations adjacentes terrestres, augmentant les possibilités de croisements apparentés. La migration, en milieu insulaire, pourrait également être plus faible que sur l'unité terrestre. La structure d'âge plus complexe des populations insulaires (Gagnon 1990; Gauthier, Gagnon et Bergeron données non publiées) , sous 1 'effet du régime des perturba-tions, où des générations successives se chevauchent, a pour effet d'augmenter encore les chances de croisements apparentés et ainsi, réduire la taille effective des populations (Futuyma 1986). Tous ces facteurs pourraient contribuer à complexifier la structure génétique des populations de cette unité de paysage comparativement à celle d'une unité de paysage moins morcelé , telle que le territoire terrestre adjacent.
C'est dans ce contexte général que se situe la présente étude. Elle a deux objectifs principaux visant à déterminer si, à une échelle régionale:
et,
1) l'insularité et le régime des feux affectent la structure génétique des populations de pins gris, en augmentant la dif-férenciation entre les populations insulaires comparativement
13 2) des régimes des feux distincts exercent des pressions sélectives qui engendrent des différences dans certaines
caractéristiques de populations de pins gris qui y sont
soumises.
La thèse est divisée en deux grandes parties. La première, constituée du chapitre I, vise à répondre au premier objectif tandis que les chap i tres II et III concernent le deuxième objectif. La problématique et les objectifs spécifiques de chacun y sont
claire-ment établis, puisqu'ils sont rédigés sous forme d'article2
• Le
texte qui suit vise toutefois à replacer chacun des chapitres dans un c ontexte plus général.
3.1. Effets de l'insularité et des régimes des feux sur la
structure génétique
Afin de déterminer si l'insularité et le régime des feux de populations de pin gris affectent leur structure génétique, nous avons utilisé des marqueurs génétiques, les isoenzymes. Le dévelop-pement et l'utilisation grandissante des techniques d'électrophorè-ses ont permis, depuis une vingtaine d'années, d'augmenter nos connaissances de la génétique des populations de plusieurs groupes d'espèces animales et végétales et ce, particulièrement chez les conifères (Lewontin 1985; Hamrick et Godt 1989). Auparavant, notam -ment pour les arbres, la plupart des études portaient sur des caractères morphologiques polygéniques, tels que la taille ou la hauteur, caractères qui sont grandement influencés par
l'environne-ment. Ces études, bien qu'elles permettent de quantifier les
variances génétique, environnementale et phénotypique de caractères polygéniques, nécessitaient de faire croître les arbres dans des conditions similaires en plantation, une méthode aussi longue que coûteuse à réaliser. Par contre, les isoenzymes ont l'avantage de
2Bien que les manuscrits anglais se rapportant aux deux
premiers chapitres soient soumis au Canadian Journal of Forest
Research, ce travail est présenté en français, sous forme
d'article. L'auteure, inscrite dans une université francophone, a jugé opportun de rédiger cette thèse en français.
14 permettre de déterminer directement le génotype de l'individu analysé pour un locus donné, puisque ce sont des marqueurs géniques (Gottlieb 1971, 1981; Lewontin 1974). Cette méthode est rapidement devenue un outil important pour les études de diversité génétique puisqu'elle permet la collecte rapide et efficace de données généti-ques. En plus d'avoir permis l'acquisition des connaissances sur la di ver si té génétique de plusieurs espèces, cette technique a été utilisée pour quantifier le degré de variation géographique de caractères génétiques, ce qui était difficile, voire même impossible avec des caractères morphologiques polygéniques (Mitton 1983). Depuis les premiers travaux portant sur les isoenzymes par Lewontin et Hubby (1966), Hubby et Lewontin (1966) ainsi que Harris (1966), plusieurs études ont démontré que le degré d' hétérozygotie, le poly-morphisme et la variabilité génétique des espèces sont relativement élevés chez la majorité des organismes vivants (Lewontin 1974; Ayala 1982; Nevo et al. 1984; Hamrick et Godt 1989). Plus spécifiquement chez les conifères, les études ont révélé que plusieurs espèces ont des degrés de variabilité génétique élevés (Hamrick et al. 1979, 1981; Loveless et Hamrick 1984; Hamrick et Godt 1989). En général, les conifères forment un groupe d'espèces très variables et ce,
vraisemblablement à cause de leurs grandes aires de répartition, leur pollinisation par le vent, leur allogamie, leur grande fécondi-té, leur longévité et la continuité relaiive de leur répartition
(Hamrick 1979; Hamrick et al. 1979, 1981; Hamrick et Godt 1989). Par ailleurs, ils montrent généralement un faible degré de différencia-tion alloenzymatique entre les populadifférencia-tions, sur leur aire de répartition. Cette faible différenciation serait due au manque de barrières efficaces pour contrer la migration, principalement parce que la pollinisation par le vent permet une homogénéisation des populations, par un flux génique important (Boyle et Yeh 1987; Hamrick 1987; Govindaraju l988a, l988b; Hamrick et Godt 1989).
Une des caractéristiques intéressantes des gymnospermes est que les graines contiennent un endosperme haploïde, appelé mégagamé-tophyte. Ce tissu possède une grande diversité d'enzymes qui ont des activités relativement élevées (Mitton 1983). Ce tissu permet de vérifier de manière probabiliste si la ségrégation allélique
15 s'effectue de façon mendélienne et de déterminer les relations de
1 inkage entre les loci, sans avoir besoin de recourir à des
croisements contrôlés. Puisque le mégaspore haploïde se divise de façon mitotique pour engendrer les autres cellules de l'ovule, l'embryon possédera la contribution gamétique maternelle identique au génotype haploïde du mégagamétophyte. Ceci permettra, si l'on fait migrer l'embryon et le mégagamétophyte sur le même gel, de dé-partager l'apport maternel de l'apport paternel.
Il apparaît difficile de déterminer la contribution du poly-morphisme isoenzymatique à la valeur adaptative d'une espèce. Les tenants de la théorie neutraliste estiment que les alloenzymes sont relativement neutres, conférant pas ou peu d'avantages directs aux individus qui en sont munis et qu'ils sont principalement maintenus dans les populations sous l'effet de la dérive et de la migration (Kimura 1968). Par contre, les tenants de la théorie sélective suggèrent que le polymorphisme est maintenu sous l'effet de la sélection naturelle. Il semble aujourd'hui que la majorité des variations alloenzymatiques ont de faibles valeurs adaptatives et que les forces évolutives telles la dérive et le flux génique, joueraient un rôle important dans le maintien du polymorphisme à ce niveau. Toutefois, certaines études révèlent que les hétérozygotes sont avantagés en termes de croissance ou de capacité reproductive (Mitton et Grant 1984; Bush et al. 1987). De plus, le polymorphisme enzymatique n'est pas réparti équitablement selon la fonction remplie par l'enzyme (Prakash 1973; Johnson 1974). Quoiqu'il en soit, ce type de marqueur a été choisi car il permet d'étudier si-multanément plusieurs loci et d'en déterminer les fréquences alléliques, de sorte qu'il est possible d'analyser la structure et la variabilité génétique des populations. Les isoenzymes sont donc des marqueurs génétiques qui sont affectés par les différentes forces évolutives qui interagissent sur les populations étudiées.
16
Les deux populations insulaires de pin gris sélectionnées sont composées d'un faible nombre d'individus comparativement aux populations de l'unité terrestre, augmentant ainsi les probabilités de croisements apparentés. De plus, la migration entre les îles pourrait être plus faible que celle observée entre les populations terrestres où, suite à un feu, de grandes populations de pin gris s'établissent. On s'attend donc à ce qu'elles soient plus sensibles
à la dérive génétique que les populations terrestres adjacentes, même si le flux génique, à cette échelle, devrait être suffisamment élevé pour empêcher la perte ou la fixation d'allèles, strictement sous l'effet de la dérive. Finalement, il existe des différences dans le régime des feux des deux unités de paysage, celui de l'unité insulaire étant plus complexe que celui de l'unité terrestre. Par conséquent, la dynamique des populations est également différente entre les deux unités de paysage (Gagnon 1990; Gauthier, Gagnon et Bergeron, données non publiées). Les structures d'âge plus complexes, engendrées par le régime des feux du lac, où des généra-tions successives se chevauchent, pourraient également favoriser les croisements apparentés, réduisant alors la taille effective des populations. De plus, les régimes différents peuvent exercer des pressions sélectives différentielles qui contribueront à distinguer les populations insulaires entre elles de même que des populations terrestres. Les marqueurs isoenzymatiques pour cette portion d'étude permettent de quantifier la structure et la variabilité génétique de populations de pins gris qui se retrouvent dans des unités de paysage qui diffèrent en terme de configuration. Ces marqueurs seront affectés par les quatre grandes forces évolutives, de telle sorte qu'ils nous donnent une image de la structuration et de la différenciation entre les populations selon l'ensemble des forces en jeu.
17
3.2. Effets sélectifs des deux régimes des feux
Il est peu probable que les marqueurs isoenzymatiques répon-dent fortement aux pressions sélectives différentielles des régimes des feux. Par contre, des caractères adaptatifs tels que 1 'épaisseur de l'écorce ou le degré de sérotinisme peuvent conférer des avantages directs aux individus qui en sont munis, selon le régime des perturbations qui les affecte. Dans cette optique, le caractère choisi pour vérifier l'effet des régimes des feux différents est le sérotinisme3
• Ce caractère est considéré comme étant une des
adaptations majeures de plusieurs espèces à la récurrence des feux de forêt (Vogl 1973; McMaster et Zedler 1981; Cayford et McRae 1983; Cowling et Lamont 1985; zammit et Westoby 1988; Lamont et al. 1991 ) .
Chez le pin gris, on sérotineux tout comme
latifolia Dougl.) et
observe des individus sérotineux et non
chez le pin tordu ( Pinus contorta var.
le pin polymorphisme est essentiel pour l'environnement de se manifester
rigide (P. rigida Mill.). Ce
permettre à un effet sélectif de (Rudolph et al . 195 9 ; McMaster et
Zedler 1981; Muir et Lotan 1985). De plus, le contrôle génétique assez élevé de ce caractère a été démontré (Rudolph et al. 1959; Teich 1970; Sittman et Tyson 1971). Ces travaux suggèrent un
contrôle génétique assez simple, vraisemblablement monogénique à
deu x allèles codominants (Teich 1970; Sittman et Tyson 1971).
Afin de pouvoir répondre au deuxième objectif, un certain nombre de vérifications concernant le sérotinisrne s'imposaient. De fait, des études rapportent que les jeunes individus ne portent pas de cônes sérotineux (Cayford 1957; Schoenike 1976) et les facteurs qui affectent l'apparition de ce caractère sont inconnus. Le second chapitre vise à déterminer à partir de quelle taille ou de quel âge
3 Le mot sérotinisme vient du latin serotinus qu i signifie
venir tardivement. Selon le Larousse du XXe siècle, i l s'agi t du rapport existant entre l'époque de la floraison d'une plante et la quantité de chaleur qu'elle a reçue. Dans la littérature botanique
le mot anglais serotiny réfère à la dispersion tardive des graines.
18 le sérotinisme apparaît chez le pin gris et à définir, de façon empirique, si l'absence de sérotinisme chez les jeunes individus est liée à leur juvénilité.
Le troisième chapitre vise à démontrer que des régimes des feux distincts constituent une pression sélective sur le sérotinisme des arbres des populations qui y sont soumises et ce, à un niveau régional. La récurrence des feux est généralement considérée comme le principal facteur ayant exercé une pression sélective sur le sérotinisme, tant chez des espèces boréales, comme le pin gris ou l'épinette noire (Picea mariana (Mill.) BSP) que chez le pin rigide
(Pinus rigida), chez des espèces de climats méditerranéens, telles
les Banksia australiens ou encore des espèces du genre Pinus de la côte ouest américaine (Hendrickson 1972; Vogl 1973; Schoenike 1976; McMaster et Zedler 1981; Cowling et Lament 1985; Muir et Lotan 1985; Zammit et Westoby 1987a et b). Le sérotinisme dans le genre Pinus apparaît s'être développé à partir du caractère ouvert des cônes et serait apparu indépendamment dans différentes lignées du genre (Duffield 1952; McMaster et Zedler 1981). McMaster et zedler (1981 ) suggèrent d'ailleurs que se sont les pressions sélectives des régimes des feux différents qui expliquent la variabilité observée dans le degré de sérotinisme des pins de la côte ouest. Pour des espèces du genre Banksia, la variabilité observée dans le sérotinis-me des espèces correspond à la variation dans les comportements géographiques et historiques des feux (Cowling et Lament 1985). Au niveau intra-spécifique, des études ont également montré des variations dans le degré de sérotinisme qui seraient liées aux variations dans le régime des perturbations par le feu (Givnish 1981; Borchert 1985; Cowling et Lament 1985; Muir et Lotan 1985).
Les graines dans les cônes sérotineux sont viables pendant plusieurs années, constituant une banque de graines sur l'arbre. Après un feu, les cônes sérotineux s'ouvriront, sous l'effet de la température élevée, pour permettre la dispersion des graines, tandis que celles déjà dispersées pourront avoir subi les dommages du feu (McMaster et Zedler 1981; Muir et Lotan 1985). Cette situation confère un avantage aux individus sérotineux pour la régénération
19 après un feu (Vog1 1973; Zedler 1977; Perry et Lotan 1979; Zamrnit et Westoby 1988). Toutefois, ces individus sont moins susceptibles de profiter d'autres types de perturbation que le feu, telles les trouées laissées lors d'un chablis, comparativement à des individus porteurs de cônes non sérotineux (Muir et Lotan 1985; Zarnmit et Westoby 1988). Pour plusieurs espèces ayant des fruits ou des cônes sérotineux, on estime donc que les individus sérotineux sont favorisés lorsque les feux sont suffisamment fréquents pour se
produire dans 1 ' intervalle qui correspond à la longévité des
individus de l'espèce (Vogl 1973; Zedler 1977; Perry et Lotan 1979;
Givnish 1981; Muir et Lotan 1985; Zamrnit et Westoby 1988). Le
patron spatial et l'intensité des perturbations sont des paramètres
importants du régime des feux, qui risquent d'influencer les
stratégies reproductives d'une espèce (Keeley 1981). Ainsi, des feux de grandes surfaces et d'intensités élevées devraient favoriser les individus sérotineux, chez des espèces où la régénération végétative n'est pas possible, puisque dans de telles circonstances la régéné-ration de tout le territoire incendié à partir des progéniteurs dans des sites protégés, serait pratiquement impossible (McMaster et Zedler 1981). Les feux de faible intensité sont, pour leur part, susceptibles de favoriser les individus mixtes et non sérotineux
(Muir et Lotan 1985). L'histoire locale et récente des feux est
également un paramètre qui peut contribuer à différencier les
populations en ce qui concerne leur degré de sérotinisme (Perry et Lotan 1979; Givnish 1981; Muir et Lotan 1985).
Dans cette optique, nous supposons que, même à un niveau
régional, les pressions engendrées par des régimes distincts de perturbations par le feu sont suffisantes pour affecter le degré de sérotinisme des populations de pin gris et ce, même si le flux génique est susceptible d'être important (Givnish 1981 ). Perry et
Lotan (1979), dans un modèle mathématique, proposent que le
maintien, au niveau régional, du polymorphisme du caractère
sérotineux chez P. contorta , est le résultat de régimes des feux variables dans l'espace ou dans le temps.
20
A notre connaissance, il n'existe aucune étude visant à
démontrer que, à un niveau régional, des régimes des feux distincts peuvent constituer une pression sélective différentielle suffisante pour engendrer des différences importantes dans le degré de sérotinisme des populations. Le régime particulier des feux du lac Duparquet nous offre la possibilité de tester l'effet de régime des perturbations à l'échelle du paysage, sur un caractère adaptatif tel que le sérotinisme. De fait, ce lac situé en forêt boréale possède un régime des feux dont plusieurs paramètres diffèrent
substantiel-lement de ceux de la forêt boréale (Bergeron 1991) . Le trois i ème chapitre vise donc à démontrer que des régimes des feux différents dans l'espace mais relativement constants dans le temps constituent une pression sélective importante sur le polymorphisme du caractère sérotineux des cônes chez le pin gris et ce, à un niveau régional.