___----,~---'---.,- n . _ _ u <
-
... ~---"'----t
"-ETUDE. DE L' lMMUNOSU?PRESSION CONSEQUENTE A LA REACTION DU GREFFON CONTRE L'HOTE INDUITE PAR UNE DIFFERENCE AU NIVEAU D'UN SEUL Op DE QUELQUES
ANTIGENES MINEURS D 'HISTOCO)œATIBlLITE·
PAR"
PAULINE BROUS SEAU
DEPARTEMENT DE MEDECINE EXPERIMEN'ULE UNIVERSITE HeGILL
MONTREAL
o.
'.
,
~
Thèse présetttée à la faculté d~s études graduées
en vue de l' obtentolon du grade de
PHILOSOPHAE DOCTOR (Ph.D.) 'b
©
Févr'1er 1985..
)<-
'"' , ... "'-~f"""'''''''~ ~-" , ...... ~-~-... " ...
Î~~
.
r
-~"
)\1'\-;'
~
:
.
..
\\1\"
1 \,
\
la
.
" 1 '),
1 \1 ,1 1 .,,>,
~ j,
\
J . \ ' " / !:> / ,"
"
\ ~Tous
droits de reproductiob, d'adaptati~n ou de traduction réservés.(
_ 1
.
,
{" , ".
... ... ~." .... ""'--r"""-; ... ;"O ... - : ... ,... ~ ~-.~ .. "" v 1 • ~ "~""- --~-_ .... ~"' .. ~' ... 0#1 .....
.
, .......
".•
littars de transplantation de oelle osseuse, même quand oies ~onneurs et les receveurs sont appariés u niveau du lU..A, i l en résulte. fréquem-ment une réact:fJln du greffon c 'nire l'hôte (GVH).
Dans cette recherche no s avons utili~é u~ système expérimental murin pour verifier si nous ouvions induhe une GVH èntre animaux H .. 2 histocompatibles, et qui se ,manifesterait par une immunosuppressioh.
L • injec tion de'
Colt.
e. 1 ymphoId.s
de"d~n/~s
his
tOinc~mp.
tib
1 es 'des receveurs par un seu ou plusieurs système~ mineurs a, induit une ,GVH' caractérisée par une fo te diminut~on de la réponse humor~le des ~èCè
veurs, et par une bais~e maTquée de la ~t1mu1at1on mitogénique de leur!!
1ymphocy tes. ,1 • , "
.
,
Cette immunosu6pression était attribuable
à
la présence de cellules s\lppressives dans 1~ rate des animaux en GVH' •• De plus, uné étude histo:' pathologique a'r~élé que plus tardivement, le thymus des'sntm4ux en réactionGVH,
induite contre plusieurs antigènes mlneürs, montraitles
signes caractéristiques d'une dy'splasie. . ' . .
,
'1
'.
..
J.
"'j
l !.'
, ,)1
1
1
,
,1, ' " ,
..
v ' ~ _ _ _ l:! ~._ ...... "" _ _ _ _ .... "' ... _ _ " k . . _ ~r-_ .... _ - - \ . ... .-" If(
.
.
•
•
..
c·
1 . 1 ABSTBACT "-"""l ivBoné mar~ow transplantations frequthtly result in a
graft-versus-host reaction
(GVHRj
e~n uhèn donors and recipiepts a~eHLA matched.
'- In the present work wi.th mice, an experiment.1 protoeal .:
~a8 designed to verify whe~er or not a ~R.ensuin8 i~unosup
pression can occuy even in 1\e absence of a dUference at the level of the major histocompatibUity ànUgeus.,.
Injection
of
lymplloid cells' from donors his,to,incompatible, vith recipients at the level of one or multiple minor antigen. ,,-nduced a GVHR resultih& in a marked decTeue of humoral respohse
,
~t· theo redpientst and aIso 8. lack of ability cif thely lymphoc#es
d ~
.' ,
ta reapon to mitogens. \
.
\
of\ This 1mmunosuppress1on was aSsociated with the presence B.uppreE!Bor cells ln the spleen of these animiils. Fur thermore
histopathological stuqies demonstrat~d that ,the thymus es from'
animaIs expèr1encing a lat~ GVHR induced accroa~multjple minor
histocompatibil~ty'anti&ensJ .howe~ signs of dysplasia.'
.
'. " '1 " '.' .'..
{
J •• ,\
\
, \ ,\!' '
i
.
. '
----~.~.~--I ... ~" '~~""""""'~~ ... "'II"i> ... ..., ... - ... --~~-.. .. _ ... -."A--~---...---~
... ---.. -... ---..:.'·----...
î..-. _ _ _ _ _ _ _ _ _ '-!<p _ _ _ _ _ iii.'
\..
!
l ' .\
"..
, A Michel,-
.
'f,.
..
\ '", ~,'f
---'- "J(
.--
; 1> ...,
~ , ,\,
..
II " IJ .....
-'
" ";l, " '-, ,,' "-, -.r-,:<-"-) .; ~ ~I.(:
\','\
.-~.
'1 ~' • j \ : } ~ . , -~ '~.:\,1
1 ,;1 j ',~.~!\ d ,;,. " ... , " ~ 1 ~ ' ..."
.
,....
" " Jo If",
".
.
<'
,
...
f ... l" • " , \. -, 1 .<.~
*
,
TABLE DES MATIERES,
~O~tjtE •••••••••••••• l ' r I ' • • • • • p • • ' • • • • • • • Q • • , • • • • • • • • • • " • •
ABS TRA.Cf . . . " , , .... .
DEDICACE •• 'so' • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • •
T.ABLE DES MATIERES ••••••••••.••••..••••••.•••• ,. •..••••• LISTE DES TABLEAUX •.. ~ •• " •....•••...••.... -•••....•••
LISTE DES FIGURES ••••.••••••..•••..•..• •••.. -. ... .
LISTE DES ~REVIATIONS •••••••••••••••••••••• , •••••••••••
INTRODUCTION .••••••• , •.•••••••••••••••••••••••••.•••••• CHAPItRE l
..
~ Revue ,bip~iogrâphique ••••.••.•••. " , , •••• 1.~1.2
1.31.4
1.5 His torique ... ' ... ~:.,Les types expérimen taux de
1 réac: t ion GVII •••••••.• •••••••••••••••
Popu~at1on(s) cellulaire(s-)
tesponsù1e(s)' de "1' indu~tion
de ~a réact ion GVH •••••••••••••••••
Le co~plexe majeur ,~
d' his tocolllpa tiblli té, ••••••. , •••••• 1.4.1
1'.4.2
1.4 .. 3 Introduction •••••••••••• ' •••• Le c01llP1exe U-2"" •••••••••••"
.
La région T~a ..• , •••••• , ••••...
L'imarumogénétique' de laréaction du greffon. contre
1
l' hô te . . . , . . . .
1.5. l L'es aIl tigènes maj eurs
d'histocompatibilité., .•••
1. 5·~ 2 Les antigènes mineurs
.' d'histocompatibilit:é., .•••
....
1.6 Les conséquences sur la
réponse immun1t~ire de la
~"téaction du greffon contre
i
'hôte :1ndu:tte contre desan tigènes Ùlaj eurs
d 'his tocompatibili té ... . 1. 6 .1 Observations pr~liminair.es •••••••.•.••• ~ • 1.6.2 La' réaction GVH et . l'influence ;riralé ••••.••• ~. , J
.,
.
--«'"
vi '\ '('?..
11i iv v .vi /1" lÇiii xiv xvi 1 3 3 5v
10 1"J';
10 • l > 11 14 15 20 26 26 27).
fL,
"\.
~
q ""'.
1.6.2 .'1, La résativation i.
d'ün virus..
" , '" ".
, 0 endogène . . . 28 1.6.2.2 La réaction)t~VH.,
e't l'indue tlon
"
t
de tumeurs ... " ... 29 1 1.6.2.3 La -réactidn GVH"
et'l'auto- " f~ ll1lmuni té ... , . . . 31 C'\
1. 6'.3.
L~ réaction GVH et'immuni té non spécifique: ... 33 '-.. .. ,
.
,1'.6.3.1 Phagoèytose .et' résistance ••••••••• 33 , « 1 1.6.3.2 Cellules
m<..
Pros tàglap.dines et Interféron ..•••• ;34..
.; 1.6.4 La réact'ion GVll et la ,réponse à médiation humoral*e ...•...•.. ... ~ •. ,. -37 ~~ j o 'C " 1 réaction, 'liVH , 1.6.4.1 La , " ,\
et effEl't 37 ' 0 ~ al1ogénéiqüe ••.•.••,
, 1 1.6.4.2 'La r-éaction GVH. .
et 1 'augment~-tian de la réponse pumo.ra~e ••• ) 3-9 ,1.6.4.3 La réacti~n G~i
0 et 1~, suppre~sion'.
'\'dè
la, réponse (',Î
\,
humorale ••••••.•••• ' 4:\, j -~, 1.'6.5 La réactipn Gv,H et l'a 1,.
réponse à tnédJation - " )•
cellulaire ... ~ ... / ... 45 " i'1
1.6.6- La réaction GVH et les -(cellules suppressives •••.• ',' 47 f"'On
r
1.6,.7 L'M.stopatheilogie de
~V
,contre la réaction' du gref!on. 1 'hôt e ... 51, 1) '. ..,,,\,1' '-2 :0 --8 1.6.r.l L 'histolosie f ~I, " 1-".
...
générale ... 51....
-0,.
,
1.6.7.2 L '~tude' histo-.
0 " J logique et.
0•
..
fonctionnelle <>...
l.
<::1 du thymus durant , une réaction du..
1 grdfon, tontre /.
,
~.
1 1 1 f,hôte" •••••••••••• 54,
J \ -g '110 \-~1..
,,' ---~- ~---..; --- ------(
/ 1.6.7.3 Discussion sur l~importance des antigènes la au cours de la réaction du greffon contre l'hôte . . . • . . . . 1.6,.8 La réaction GVH et la moelle osseuse •.•...••.... 1.7 Description et conséquences de la réaction du greffon contre l'hôte induite contre plusieurs antigènes mineurs57 58 d' histocompatibilité. ~ . . . '. . .. 60 1.7.1 1. 7.2 Introduction ..••... ~ ... Antigènes mineurs et induction de réaction GVH léthale . . . • . . . 1.7.3 La réaction GVH et la réponse à médiation 60 60 humorale. • •.•.•. . . ...•. • •.•• 64 1. 7.4 1. 7.3.1 La réponse humorale dirigée contre le soi. ••••. 1. 7.3.2 La réponse humoralë dirigée contre le non-soi . . . , •.... La r&action GVH et la 64 66 réponse à médiat~n ~ cellulaire. .•.... •...•. ... 68 1. 7.5 L'histopathologie de la réaction GVH .••••.••••• "..., • 69 1.8 Description et conséquences de la réal;tion du greffon contre l 'hôte induite contre un seul antigène mineur
d'histocompa-tibilité. soit le H-Y . . . .. 72
viii 1.8.1 Introduction ....•.•...•.•. 1.6.2 L'antigène H-Y et la 72 f~ réac t ion GVH •....••••...•••.
7~ ~'
1 ... 9 ,Çc.nclusion . . . f . ' • • • • · • • • • • • • 78l '
- - - .
ix
-,
CHAPITRE
2 Matériel et Mé thodes .•••..•...• , ... 812.1 Les animaux .. , ... , ... 81 2.2 La suspension cellulaire •••.•.•••.. 82 2.3 L' aIltigène ... , ... " ... 82 2.4 La greffe peau bilatérale
~ 2.5 L'induction de la réaction dorsale ... , ... GVH ••••• 83 82
2.6 La technique d'hémolyse en
milieu liquide .•••..•..•••....••.. ~ 84 2.7 Histologie ••....•••.•..••.•.•.••••. 86 2.8 La transformation blastique ...••... 86 2.9 Production in-vitro de
cellules formatrices d' antic0!'fs •.• 88
)
2.10 Détection des cellules Thy-1.2 .•.. 91 (
2.11 Détection des cel'lules NJ<. ... 92 2.12 Préparation des so ut'-popula t ions
de cellules adhérentes et
non-adhérentes ...••...••.•..•••.• 94 2.13 Statistiques ....••...•...•. " .•.. 95
CHAPITRE 3 Travail expérimen tal ... , ... 96
,
1
~
3.1 Introduction •....•...••••..•.•.• 96 3.2 Le système eJllpérimenta1 1:
plusieurs antigènes mineurs
d 'histocompa tibili té .••.••••••••••• 97 'i , 3.2.1 Cinétique de la réponse humorale in-vivo ... 98 3.2.1.1 La réponse humorale directe ..•••..•••.• 98 3.2.1.2 La réponse humorale indirecte ••••••.••• 101 3.2.2 Cinétique de
la
transfor-mation blas tique .••••••..••• 101 3.2.2.1 Cinétiques des
réponses .... a la
"
PRA
et à la Con# .... ,104•
3.2.2.2 Cinétique dela réponse au, LPS ••
.
i073~2. 3 Mécanismes impliq~s dans
~ la suppressionDde a
réponse immunitaire ••.•••••• 113
~---=---(
..
(
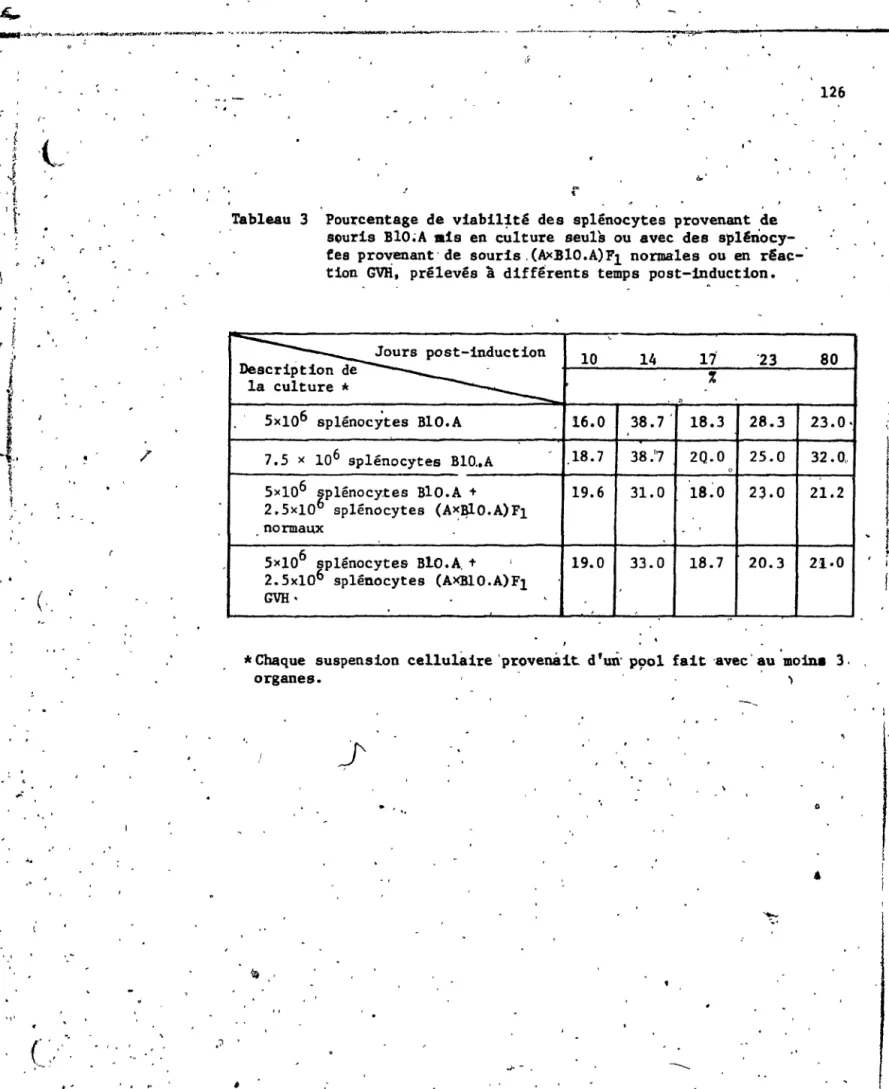
3.2.3.1 Cinétique de 1'activité NK •...•...• 3.2.3.2 Cellules 113 suppressives. . .•. .•• . ••• 115 3.2.3.2.1 Eta- blisse-ment de la Con-centration de cellules parentales ••• 3.2.3.2.2 Co-cult~ semi-all,?-généique . . . 3.2.3.2.3 Co-culture syngénéique •• 3.2..3.3 Etude de l' antigène Thy-l et histologie 11& 119 du thymus... 128 3.2.3.3.1 Acquisition de 1 '<an tigène Thy-l ... . 3.2.3.3.2 Histopatho-logie du 131 thymus. . • • • •• 1343.2.4 Résumé des résultats ••••••••••• ~. 139
3.3 Système expérimental 2: un seul
système mineur d'histocomp~tibilité... 139
3.3.1 H-"Y... ••• .•• •••.•••• •...•••.••• 140 3.3.1.1 Cinétique de la réponse humorale in-vivo. • • . • • • • . . . . . • 140
.
-~,
3.3.1.1.1 Donneurs non pré- sensibili-ses ... . 3.3.1.1.2 Donneurs pré-sensi-6 140 bilisés. ~ . • • • 144 3.3.1.'2 Cinétique de latrans-fonna·tion b1as tique ••••.
,
•
xi ~ .... 3.3.1.2.1 Cinét,!que des répon-ses" à la PHA et à la Con A .••••• 147 3.3.1.2.2 Cinétique des répon-ses au LPS .••• , 152 3.3.1.3 Cellules suppressives ..•..••.••••• 156 3.3.1.3.1 Etab1iss~-ment de la " concentra-tion opt!'!" oylle de, cellules' de souris CIl.C57BL/6
f ~melles ••• ; .• 156 ) 3.3.1.3.2 Co-culture ~ de sp1éno- l , , cytes prove- ~ ( nant de } souris mâles et femelles ••••.. 159 343.1. 3.3 Co-culture de apléno-. cytes prove-nant de souris mâles ... 162 3.3.1.4 Etude de l'acquisition. de l~antigène Thy-l et histologie du thymus \il ••••••••••••••••• 164 3.3.1:4~1 Acq\lisition \ ~ de l'anti-\ gène Thy-l ... 164" ---3.3.1.4.2 Histop'atho-10gie du thrmus ..•.••••• 1673.3.1.5 Résumé des ré sul tata ••••• 167
(
3.3.2 H-l •••••••••••••• ~ •••• _ .... , ••••••• 170\,~
p-
,CHAPITRE 4 3.3.2.1 Cinétique de la ~ )
,
reponse humo~ale in-vivo . . . lit_ • • • 3.3.2.1.1 Donneurs non présen-~ sibilisés •.••• 3.3.2.1.2 Donneurs pré-sensi-bilisés ••••••• 3.3.3.2 Résumé des résultats •••••Discussion et coaclu.ioR •••••••.••••.••••.••• 4.1 4.2 4.3 4.4
4.5
4.6 In troduction ... _ . . . . Conséquences de la réaction GVH induite par une différence au niveau de plusieurs antigènes , mineurs d'histoco~atibilité •••••••••••• -Conséquences de la réaction GVHinduite par une différence au ' , niveau du complexe H-Y ou H-1 •••••••.••• Comparaison entre les conséquences ' causées par aes réactions GVH de
diverses intensités •••••••••• : •••••••••• Situation chez l'humain ..•••••••••••..•• Conclusion générale ." •••••• ' ••. _ .••••••••• , RE}ŒRCI El-ŒNTS •••••••••••••••••••••••.•••••••••••••• Il • .. ~ • • • • • • COMMUNICATIONS ET PUBLICATIONS •••••••••••••••••••••••••••••• BIBLIOGRA.PHIE •..••.•. ; •.••••••.•.••••••••••..•••••••••.••••• AN'NEXE 1 ••••••..•••••••••••••••••••••• ~ ••••••••••.•••••••••• ANNEXE 2 .•..••.. ., ...•. !Î . . . • 1 • • • • • • • • • • • • • • • • • • • Î xii
-170 170 173 177 " 178 178 180 192 c . 199 204 206 209 211 212 246 247..
\., , ~ ,
.
" '..
-~-.
(
-
"'.
LISTE DES TABLEAUX
...
Tablaau 1 Cinétlque(qe l'activité NK (système
expérimen tal 1)"."."""".""""".:"""
.~.
'" • " " " " " " " • " •114
Tableau 2
~.r. } ... .f.
Tau'X de PFC obtenus dans les' co-cultures,
~.;~~t""
syngénéiques réalis~# s pour déceler la ( , / . ---. ~
prése~cè de ~ellule ~upprrsslves "',/
,(Systeme ·experIment 1) • ••• • • • • •• •••• • •••• • • • • • 124
Tableau 3 Etude de la cytoto'xicité -(Système,
... im 1 1 ) " . •
exper enta " " , " " " " " " " " " " " " " " " " " " " " " . " " " " " " . "
o "
Tableau 4 Co-cultures seml-a1l~généiques'réall~ées
avec dèS splénocytes fractionnés o~ non,
prOve1)Bn t d'an imaux témo ihs •• ' ... .
. ~
Tab1~u' 5 ·Co-cù1~ures. s,eJ!li":'dlogénéiques réalisées .;
avec des splénpcytes 'fractionnées- ou non,
.'provènant,d'animaux 'en réactio~ GVH ... ~~ ••
..
Tableau 6 Etude
~·partir'd~.thYmus ~u'nombre
normal', de ceHu1es et du pourc.enfàge du nombre
: de çellul,ës Thy":~ + i 'diffétents ~emps :
post-~nduction. de même qu'un 'résumé, des
-T.abieau 7
'\
.
"
Tableau 8
ob'~eryat1ons hbto1og1ques (Système ~
expérimental 1)""""."" _(le" " " " " " .~. " • " " " " .. " • " " " " " ....
,
.
Taux tle PFC
~obten'us
"dans les co-culturès,'"~éailsées pou~ dic~ler l~ fr~sence des .
cellules suppres~iv~s dan~ la'rate des
~ntmaux ~ya~t développé unè réaction GVH
... " ' . ..
contre le lt--Y ••• i " ••• \ • • • • • " • • • • • ... " ....
... .. .. 'l .. •
. Etudè' à 'pa:r..tlr 6u thymus' du ·nombre normal
de cellules et du pouréentage du nombre
de, ceÜules Th.y:-l~. à. différe,n'ts t~ps
p0E!-t ... lnd\lctlon de même qu
'un
résumé. des "lt1 observations' histologiques (Système "
expér:~ental "2) •• :-, .~ •.•••• •• ,,: ... , ~.' •••• :. ... ~
() ..
.
.
" , '..-.
Tabl~au 9 Classlficst!on proposée ~our_~a rêactio~
, GVH tnauite ~ontre dlff'tentes d~spa~lt's
ant~8~n1ques ... ft. . . . ~ • • • • • • • • • • • • • • • • • • .' 126 -129 13Q 13.j " ,163
16-5
20S \ , xiii.-Figure 1 Figure 2' Figure 3 Figure
4
Figure5
(
Figure 6 Figure 7 Figure 8 Figure 9 Figure 10 Figure 11 ,. Figure 12~
Figure 13 Figure 14 , )),
.
/
l:ISTE DES FIGURES
Schéma du sys tème H- 2 ••••.••••••••••••••••••••. , ~ •
Représentation d'une'paTtie du gên6me .
\
'.
,
mur1n •••.•••.•....••..•••..••. Il . . . .
Représentation schématique d'une chambre de
1
13
,25
&rbroQk. . . . . . . . . . .. . . . . . . . . . . . . . . 90
.0
Ciné tique de la réponse humora1e'd irec te
(Sys tèll)e expérimental 1h .••.••.••.••...••••••..• 100 Cinétique de la réponse humorale indirecte
(Sys tème expérimen ta 1. 1) •..•...•.•.••••••••••.• 103 Cinétique de la transfbrmation blastique
avec la
PHA
(Système,expérimental1) •••••••••••••
'lO~
~inétique ~e la transformation b1astique
àvec la Con A (Système expérimental 1). '... •.•. 109
, Cinétique de la transformation blastique 1
avec le LPS (Système expérimental 1) ••••••••••••• Inf1uence'de la concentration de splénocytes de souris
BIO.A
dans:la détection de PFC112
in-vi tro. .. . . . . . ,; ., . . ... . . . . . . . . . ~ . . . 118
Etude des cel1ul~s suppressives (Système "
expérimen ta! 1) ... 1.' • • • • • • • , • • • • • • • • • • • • • • • • • 121
D-=-=-~\
Sec tion d" un thymus provenan t d'une sout;is "
(A )( B10.A)F
1 normale ... :,\~... 136
1.
Sec tion dl un thymus provenan t" dl une souris (A )( BIO.A)F
1 prélevé au jour 34 post-GVH •••••• '.' '138
\
Section d'un thymus' p~ovenant'~'une souris
(A x BIO.A)F
1 prélevé au jour 80 post-GVH... 138
Cinétique de la réponse humorale directe en , utilisant des donnéur$ pOn pré-sensibiliSés
.
(H-Y) ••••.•••••.•..• " •.• 1 • • • • • • • • • • • • • • • • • • • _ • • • • •
.
.
.
142 xiv
{
"
~._---..,
, ~ -.,-Figure 15 Figure 16 Piaure 17'-
•
•
",•
Cinétique de la réponse humorale directe
~n ùti1isartt des donneurs pré-sensibilisês
[j ~,"
(H-Y) • " ... '1/' •••••••••••
Cinétique de
la
transformation blastiqueave_c la PHA (H-y) -.! !-!...-. . . .
Cinétique de la
transfo~ation
blastique.avec la Con A (ij-Y) •••• ., ... ' •••• ., •
,
.
~~ ••••••••.Figure 18
Figure 19'
Figure 21
l" ,
Cinétique de
fa
transformation blastique . '1~ avec le LPS
r-Y) ...
~..
Inf1yence .de la co'ncent-ratipn de splénocytes de souris C57BL/6 fétnel1es dans la
détection de PFC in7vitro ••.•.••••••• : •••••••.••
Etude des cellules suppressives (H-Y) •••••.• ' •.••
,Sect,ion d'un thymus pr~venant "d 'une 8oûri~'
C57BL/6 'prélevé au jour 75 post-GVH ... .
Figure 22. Section d'un thymus provenant d'une souris '.
C57BL/6 ,prélevé au jour.95 post-CVK ••••• ~ •••••••
·''104
Figure 23 Cinétique de '~a ré.ponse humorale direc;te
en. utilisant des donneur~ non
pré-s'ensl-Figure 2~
b11isés (H-l), ••••••••••• " •• '.' ... ',"
"
.
, "
Q ,
Cinét~qu'e de la' réponse humorale directe
en ,utilisa /aes do~neurs pré'-sensibilisés
~
' ' 0 '-
-
. "(li .. \ ••••••• , •••••••••••••••• , ••• ;. . . . .
Figure
25
Représentation schématique d~s co~séquences'de la réactiQn GVH induite contre plu$ieurs systèmes miheurs d'histocomp.atibilité •••••••••••
Figure 26 Rèltrêsentation schél!latique dés conséquenc'es
.
.
146
149
151,.
155158
161169
169 1,72 ,176
190de, la réaction' GVH induitE contre 'un seul
"systèu1'e mineur d '~i~tocompat1bi1ité (H-Yl •••••• ~ " • 198
' A
'.
., "",
--'.
• 1 ~ " 1xv
• 1 ,t" ' \ ,.
." , , fJ
1i
,
.
l' l' 1,
l
, 1 11·,;
.
l
E d14
•
.., ~CHC
·
·
Con A:
~-~CPH
Q GRM·
GVH :HEPES:
\"
LPS·
·
,.
(
r'.PFC
PRA
RML·
·
"' ---Ihy~l: " el) , ' " '" , ~.
, " : L b..
.
-(
, 1 ~-~----~~~~~~~~--~----~-q---Q l)
, ,LISTE DES ABBREVIATlONS
,
,~ytotoxicité • m6dlation cellulaire
éoneanavaline A Coup par minute
"
.
, Globùle x:ouge "de mouton
Réa~t~on du greffon contre l'hôte (Orai t-versus-hos't)
" l
N-2-aydroxy~thylpipérazine-N -2-éthanesulfoniaue acide
, tlpopolysaccharide
Cell.ules formatrices de plagès de lyse
(Plaque.formi~8 cells~ ;
Phyto~émaggiu~~~~.
, ~ , ,
.
R'actlon mixte lymphocytaire
l '
Ani::Lg~ne.
tHêta,all~antig~ne
des lYmphocytes"T de:a.~
souTis..
.
o "b f ' ,,
,,
..
" ,-,,
,
" , t(~' v"~·.:;'~.,,
" '~ \~ ~ ~!. '~"'f.I."I~ '\ xvi\
.
' ",(
,.
o , j -".'
-(
•
,;, , _ ~ _ _ ...-~.~ ... ~ ... 'h ... ff"' 'of(~_ ... '-
,INTIIODUCTION
• 1Dans le ~raitement de certaines maladies très graves, comme par exemple l'anémie aplastiquè ou encore certaines formes de leucémie, la meilleure fa50n d'i.ntervenir
à
l'heure actuelle est d'irradier les,
individus.' Afi~_de prévenir les effets nocifs de l'irradiation, les
\
systè.IJles hématoporétique et lymph?po!étique de. ces patients sont recons-titués
av~c>un~:.reffe ~e.mQe~ie osse~se.
Les donneurs etl~ recev~urs
":. ' , . 1
,
sont choisis' de faço.n' à ce qU,e leurs ce.llule~ lympho!des soient
fdenti-) .. ~ & , ~. t h ,
" ques " '8\1
~ive~u',cfu
système majeurd'hi'8~ocomp8tibi1'ité, et
mutuellement; - . ~ ~" ,} , ,." ,
non réactives aans
un~
réactionmixte',lyrtlpbocyt~~re,
afin,d'év1t:er, , \
, ~ fi ~ " . ~ ..
l'induction ,d'une
~é~~fion
du greffonco~~re,lth6te
chez lesrecev~ur8.
1 • ' ,
. ':rôutefoiç:
~h:z
une°bO~?e
•.~:opor.tibn
despat~ents
(variable selon' lamal~die
etle.c~ntre
de· transplantations)t suite à la' greffe, nous,~l" to'
Il f. ~ 0, '\
nptçns !.'app~rition· de. symptôme~ qui laisse~t st?Pposer le développetnent , d'\lne réaèUon'du gteffon 'contre" l'h6te (GVH).. Dans le prhent travail,
'\ ~ ~ ~ "
nous, allons vérifier s'il est poss!ble
d'indui~e
uue réaction" du• , ~ , ~ ' , " /,l , , , ' " •
gtef~pn c~ntre
l'hôteco~tre
des anU,gènes mineurs 'dIhi~.tbcomp~t1bf~1té,·
>
.. ___ . & Jce qui pourrait expliquer c~phénomène.
,
'..
.l' 1\ 1
Pour ce faire, nous.reverrons les conSéquence' '$ur
ia
répo~s. tœmu~'
.. 1 • •
nitaille et sur l'organisme en
~énéral,
d;"une réaction GVH1ndu~:~~
çontre,1 "II:' ~., "
.
\..
'" ~ , .c"'*.:.'_ ~~ ______ -'_, .. 1.'
\'" ,",.
, -,1
1
1
,
.,
.', > • 1-1
" ·f'
11
<,l' .
..r
! .
..
, 't
c ' , '.
des
~ntigènes
majeursd'histocomp~bilité.
Par la suite, nousexamine-rons l'impor~ance .des différents systèmes mineurs d'histocompatibilité darts la réponse immunitaire. Nous en profiterons pour dégager les indi-"
.
,.,cations expérimentales"','qui permettent d'attribuer un rôle possible à ces antigènes daons l'inG!uction d'une réaction du greffon contre l'hôte.
Cl est ce derni~f point qùi sera eXPloré "par les diverses ej(périen-ces 'décrites dans cette thèse. Ces dernières nous permettront d'une . yart, de démontrer qu'une différence au piveau d'un ~eul ou de plusieurs
antigènest:inetirS
d'histocompatibilit~
est.s:ffisante pour induire une réaction GVH et d'autre part d'en déterminer les px:incipales conséquences.," ,
•
"
..
"''''''.e"
.
,,
' , ,t
• -~-,.--
-'
"...,.-t...
-~---CHAPITRE 1 ilREVUE BIBLIOGRAPHIQUE
.'
1.1 HistorigueIl faut remonter jusqu'au début du !Xe siècle pour ~etrouver d.ns '\
la littérature, l'origine de la'réactl~ du g~effon con~re l'hôte~
, ",
'0 symbolisée par les lettres GVH (de l'anglais, ~raft-versus-Host). C'~st
~ 1 ..
en effe~ Murphy en 1916 qui en greffànt des morceaux de rate provenant
dJ~ poule~ adplte sur la membrane ChQriOallanto~dle~è d'em~ry~s de
,
poulets a observé chez les receveu~s une splénomégalie et la formation de nodules sur la membrane même et, à l'intérieur 'de la
r~te.
Au même moment,'Danchakoff (1916) a dé~Ontré que la nature du ~issu greffé.
jouait 06, rôle important étant ~onné que seulement
la
rate. le foie et, ~
la moelle osseuse p~rmi'tous les tissus qu'il·a utilisés' provenant de donneurs isogénéiques. conduisaient à ce phénomène chez,.le receveur.
Ce n'est que vers les années 1950 qu'il a pu être démontré que les' cellules lymphofdes étaient
1e~
responsables du': phénOlllène (Dempster,1953; B1l1iDghalllf>e't coll. J, 1955,1956, Trentin, 1956 ~19S7 .1958; B1l1ingh.am
et Bre,nt. 1957; Simonsen. 1953.1957). En effet. ces chercheurs en vou~
'13nt étud~er le comportément de cellules lympho!des comme àn~igènet se sont,apérçus que, dans certaines conditions expérimentales. ces cellules
"
peuvent réagir immunologlquement contre des tissus de l '.hôte provoquant ainsi l'induction de la réact'ion GVH.
(
,~t
(.
4
Ce phénomène a par la suite été dé,crit chez plusieurs espèç:es ani-males ~ais il est particulièrement mieux documenté chez le poulet et la sourh.
Toutefois à cette époque. ,les conséquences de cette réaction étaient
'"
~éfinies
coîmne étant le retardd~
la cr9issance, l'émaciation, lasplé-nomégalie, l'hépatomégalie, la diarrhée' et l'an~mie. de même qu'une atrophie des ganglions lymphatiques.
De plus, ces cherçheuis ont essayé plusieurs méthodes afin de mesu-rer l'intensité de la réaction GVH. Le plus simplè a été de suivre la mortalité des animaux.> Certains autres comme Russell (1960) comparaient le poids des animaux expériment~ux à celui des animaux témoins. La pre-mière méthode utilisant~la splénomégalie comme principal critère~e
mesu-f '
re ~ été décrite par Simonsen en 1962._' La méthQde est basée sur le fait qae le degré de splénoméga~ie qui se développe-d~ns les premiers 'jours suivant l'induction de la réaction GVH est pr~portionnel au nombre de
p
cellules imm~nocompétentes, utilisées pour induire la réaction.
L~indi-ce est détermine par le quotient du rapport entre le poids de la rate "
,
et celui de l'anim~l en réaction GVH, SU! le même rapport établi avec , .;." des animaux hon injectés ou qui ont re~u des cel1~les ~ syngéné,iques.
Encore d~ nos jours, ce~ indice de splénom~galie est-utilisé dans plusieurs laboratoires malgré le fait que. ce ne soit plus l'outil tout
à fai~ recommandévpour mesurer la réaction GVH.
D'abord Billirtghatn et c:oll,aborateurs . en 1956, puis S~JIlonsen
,.
en 1957, enfai~ant
une revue te ce qui était conhude'l~
GVH ont décFit,
'.
•
,/
/
;/
\.
{
..
;(
l '-
5trois critè~es qui ~evaient toujours être respectés pour' o~tenir cette
réaction. Ces critères sont:
1) la greffe doit contenir des"cellules immunocompétentes
2) le donneur et le receveur doivent être génétiquement diffé~ents
3) l'hôte doit être incapable de rejeter la greffe.
" 1
..
Suite à toutes les expériences de transferts de cellules
immun!ocompé-tentes qui se sont faites durant une quinzaine d'années, au
début,des.an-,
nées 1960, il n'y a plus de doute sur la validité dp concept qu~~n,
gre~-fe peut réagir immunologiquement contre le receveur.
l"
Dans cette première partie de la revue 'bibliographique, nous n'avons
mentionné que les détails historiques les flus pertinents. Cel;'tal~Jc
8U'-teurs ont déjà, pub1i~ des revu? beaucoup plus c,C!mplètes sur ce sujet.
dont Sim9nsen'en 1962, McBride en 1966, Elklns ~n 1971 et Grebe et
Streilen en 1976:
1.2 Les types expérimentaux de r~action GVH
Q ;
Parmi toutes ,les approches expérimèntales employées dans' 1 t an alys'e
du système immunitaire et de sa r~gulation, la réaction GVH s'est
révé-•
•
,
1ée être un modèle reproductible de choix. ~onséquemment, i l en exis~é
plusieurs formes •
'Une desFpremières formes dgcrites a été la maladie de l'avorton ,
(runt disease),' Nous la devoI.1s principalement à ~i1lingham et
col1a-bOrateurs· (195~) qui, en faisant, des expériences pour 'rendre
immunolog,i-r 1 i 1 f' 1 1
!
l' j, 1
'. '1
.f
:.,j ,.
. 1
! " , l, 1 , i 11
!
1•
1
!
'1 •quement tolérantes d,es souris nouv.eaux-nées à certains antjgènes pa~entaux,
6
se sont aperçus que selon la souche des souris utilisées, la
greffe pouvait provoquer la mortalité de 10070 des animawc utilisés. Pour 'obtenir une telle réact ion, des animaux immunologiquement immatures
doivent recevoir des lymphocytes T parentaux. Les conséquences sont ~e
retard dans .la croissance. une alopécie. une destruction de tissus 1ympbo~des et éventuellement la mort.
Une seconde fonne de GVH se nomme la maladie de type. secondaire (secondary disease). L' hôte doit subir une irradiation léthale et i l
t doi t recevoir des lymphocytes T allogénêiques. Les conséquences décri-te A'Bil1 ingham et coll., 1956) semblent similaires à celles énumérées . précédemment, sauJ qu'il s'agit: ici d'un système plus contestable étant
\
donné que les effets de l ' irradiatiôj peuvent s'ajouter aux veritables conséquences de la réaction, pouvant ainsi les modifier.
La troisième forme est celle de l'intoxication parabiotique (para-biosis intoxication) qui est l'établissement d'une circulation sanguine périphérique croisée entre deux organismes immunocompétents et histoin-compatibles. Comme conséquence à ce type particulier de greffe, i l en résulte une anémie ainsi qu'une perte de poids. la destruction de tis-sus lymllho!des et épithéliawc ains!l qu'une amylo'l'dose (MoBride, 1966).
Finalement, la.t;quatrième forme de GVH est celle de la maladie de l'hybride F~ (FI hybrid disease)."" Elle consiste ,en l'injection de lym-phocytes T parentaux dans une lignée hybride YI résùltan t du croisement de parents histoiocompatibles: Les conséquences sont les mêmes que ce1-les décrites précédemment et l'intensité avec laquelle elce1-les se manifestent
dépend du degré d'histoincompatibi1ité ent~e le donneur et le receveur CMcBride, 1966).
{
" 7
Mainten<lnt que nous avons étudié l~s quatre formes expérimentales
de la réaction du greffon contre l' hô~, nous allons discuter de la
cellule qui est capable d'induire une réaction,GVH.
1. 3 Population(s) cellutaire (5) responsable (5) lie l ''induction de la
réaction CVH
c'est à Simonsen en 1962 à qui revient la description de,la
réac-tion du greffon contre 1 'hôte comme étant u.n ph~nomène à médiatiôn cellu-,
laire. Toutefois étant donné que les populations 'lymphoïdes inj ectées
pour obtenir des réactioI\s GVH n'étaient non pas purifiée.s, mais compor-taient plutôt des mélanges de cellules, il élevenai t important de carac-tériser la ou les cellules responsables de l ' induc tion de ce pnén'omène.
Miller en 1961 et le groupe de Martinez en 1962, ont démontré que la thymectomie néonatale de souris les rendait incapables de rej eter
des greffes de peau ou -de tumeur, Ils postulèrenf alots que le thymus
jouait un rôle fondamental dans la pleine expression de réponses immu:" nitaires.
Dalmasso. et collaborateurs en 1962, en voulant déterIlhner si le,
thymus était aussi impliqU'é dans ~a réaction GV'H ont démontré l'
ilnca-pacité de splénocytes de souris th~ectomisées à la naissance à
indui-re des réactions GVH et ceci, en utilisant le modèle de l' hybride FI
suggérant ainsi ~n rôle essentiel aux lymphocytes T dans l" élaboration
de la réaction GVH.
1
,
C'est Yunis et collaborateu,rs en _1964, qui confirmèrent ce point
en rétablissant la capacité de ,souris thymectomisées à la naissançe à
-,
,
.
"
,
(
8
induire une réac t ion G par l'injection de thymocytes. Ils concluent de ce travail qu'une .ce Iule dérivant du thymus serait responsable de
l,
l ' induct ion de la réact on GVH.
expériences de Cantor et collaborateurs en 1970. portant sur le entiel dé différents organes lymphofdes à induire des/ réactions z la souris en ut 'l1sa~t l"e modèle de la maladie de l'avorton, démontrent que les canal thoracique e~ du sang périphérique, étant principalement de ture T, sont environ quatre fois plus effi-caces que les eellules p ovenant de la rate ou de J.a moelle osseuse à
induire des. réactions G expliquée par le fait dans la rate et dans la thoracique.
Cet te différence d'intensité pouvant être les lymphocytes T sont beaucoup plus dilués
~ oelle osseuse que dans le thymus où le canal
-
'\-Argyris en 1974 a r alisé une expérience avec l'aide d' antisérum
.
spécifique, pouvant éliminer les lymphocytes T, B Ou les macrophages. Le potentiel de la ou des populations cellulaires résiduelles
à
induire des réactions GVH était déterminé par l'utilisation du modèle de 1 'hy-bride FI' Les conclusions peuvent se résumer en trois, -points:---
.
a) une populati.on de lymphocytes T est essentielle à l' induc tian de la réaction GVH
b) une population de macrophages pourr,it contribuer à obtenir'une réaction GVH de plus forte intensité
c) les lymphocytes B ne sont pas essentielp pour l'obtention d'une réac t ion GVH.
r
(
~...
(
---~---9
t
Il est maintenant clairement établi que le lymphocyte T joue un
rôle ess.entiel dans l'induction de la ré~ction du gre.ffon contre l'hôte.
La possibil i té de l t exis tence d ~un type de coopération cellulaire
dans le phénomène d'inducti,on de la réaction GVH, comme 11 en existe un
dans la production des antico~ps, a été étudié par Cantor et Asofsky en
1970 et 1972. Ils ont travaillé chez la souris en utilisant ~le modèle
de l'hybride F l' Leurs résultats leur' permettent de conclure à la
parti-cipation dt au moins deux types de lymphocytes T. Le premier qu'ils ont
nommé Tl serait la cel.~ule effectrice du phénomèI\e., C'est une cellule
immature, à vie courte" non circulante et" qui se ratrouve' principalement
gans la rate et le thymus. C'est ...un~cellule très sensfble à l rection
d' un
s~rum
anU-thêta. Le second lymphocyte' nommét
2 s,erait~a 'cellul~
amplificatrice du phénoniène. C'est une cellule plus mature que Tl'
cir:-~ulante
et à vie longue. Elle est moins sensible à l ' effèt d'un sérumanti-thêta que Tl' ._ ~l1e se retrouve principalement dans le sang'
péri-- .... ~ ,
phérique et dans les ganglions, Cette hypothèse a par la suite été
con-firmée par d" aut~es exp~rienc;s (Hilgard,. 1970; Cantor. 1-972).
Plusi"eurs chercheurs travaillant dans le dàmaine de la différ.enc'i/il- _ tion des lymphocytes T ont essayé par la suite d'analyser cet te hypothèse
de coopération cellulaire dans l'induction de la r~action
GVa.
en f.onc-'tion des différents modèles c:lè maturatioit thymique, de hçon à
caracté-riser davantage les deuX populations cellulaires impliquées. Toutefois,
"
i l est très diffièile de compléter la caractérisation. étant
. plusieurs modèles de
~turatl:i.on
existent et qu'aucun ne(faitl'unanimité.
"
/
,
donnë que encore
\
\
(.
10
•
Selon cert,ins auteurs (Zollinger et Potworowski. 1979; Paquette
et coll •• 1981; Potworowski et Fournier. 1982). i l semblerait 'que la
,
1>cellule effectrice Ly T 1- 2,3+ aurait'une forte concentration de
Surface en,antigène Thy-l, alors que la cellule amp+ificatrice soit
, +'
-corticorésistante. Ly T 1 2,3 et possède une faible concentration
de surface en antigène Thy-l, ~ qui pourrait ~pliquer la différence
de sensibilité au sérum anti-Thy-l déjà signalée plus haut. ,
Toutefois. d'autres expériences sont à faire .dans le but de mieux
caractéris~r les antigèn~s de surface de ces deux populations cellulaires.
A plusieurs reprises. nous avons mentionné que pour qu' 11 y ait
une réaction GVH. il' devait obligatoirement y avoir une incompatibilité
,g~nétique entre le donneur et le receveur.
Nous ,al;t.ons maintenant étudier 1 'influen~e
du
système Ilaj eurd'histocompat!pilité dans l~~nduction de la réaction
GVH
apres avoirfait une brève revue de ce dernier.
1.4 Le complexe maJ eur d 'histocompat"iibilité
1.4.1 Introduction
Les systèmes d'histocompatibilité 'des vertébrés • f s~nt définis
1 •
étant 'des -ensembles dê gènes et de i;roduits de g,ènes qui 'rêglssent l'
ac-1
ceptation ou le rejet d'une greffe.
~
C'est
à
Gorer etâ
Sne-ll que nous devons les premières étudesfài-, '
tes dans ce domaine. Peter Gorer en 1937 et 1938 ,et avec quelques
co1-,
laborateurs en 1948, a été le premier
i
décr1ré
un sy,stèm,e cl 'histocom'"::
, ,
(
"
..
pâtibilité chez,la souris. Un peu plus tard, Snell et coll'borateurs
'(1953) ,ont poursuiyi cette ,étude en faisânt des 'expérien~es, de
greffes-~
de peau dans lesquelles ils utilisaient des ~ouris de gCinotyp.és diffé-,
.
..
rents. Suite à ces ,expériences. ils ont'confirmé l'existence d'un
sys-11
, \
tème majeur d'histocompatibilité (MHC) ,chez la souris. Ce système se' 1
situe sur le chromosome numéro i7 et est nommé le complexe H-2.
Aujourd'hlli, nous connaissons l'existence d'un système majeur d'hls-'
•
tocompatibilité pour une douzaine d ,'organismes verdbrés. Chez 1
'hu--'
..
main;
ie
système.
~ajeur d'histocompatibilité est 1ê sysûme HLA (HumanLeUkocyte'Antlgen)'qui est situé sur le chromosome numéro 6.
/ ' ....
Le système majeur d'histocompatibilité le plus étud~é et le miéux
décrit est actuellement le complexe H-2 chez la souris.~
1.4.2· Le complexe H:'2
\,
.
,.. Le polymorphisme du complexe H-2 en fait un système génkique.',
par-I
ticulier, puisque ~ontrairement aux autres genes, chacun de ceux qui
"-compose le système, maje\lr d 'histocompatibilité possède plusieurs
allè-les, éodant ainsi pour plusieurs l?roduits d:tff~rents . . L 'héréditiS de- ce'
sys~ème est co-dominaDt~ c'est-à~dire que l'antigène s'exprt .. si le
'-gène", est présent (Snell. ,1968). '.
La structure du système H-2 (~oir fig~re 1) a eté mlse,en,~videnc~
,
par l'etnp1oi de lignées pures,. de recom~inaI1ts o~ encore' de' ligné~8 \
-1 cungénique's de souris.
\
\ ' / i l '•
1
j
,
, \~.
Jo ' . Figure 1 :.
" ~chéma du ( sprènt. et " , , .. e H-Z system ',.f -" 1981) Korngolcl, /"•
..
" 1!.
"
(,.
j.' ... '~ :;,c
"-""\
~j
'"
,.
.
" '\.
(
\
\ "\.[
[
Q \ \ ...,,, .. ~"'J. ..... 01_...-.. _ _ ... _~ ... _ . _ . _ _ .j.I _ _ _ _ _ _ _ .... WiJl..., _ _ _ . . . . _ _ _ _ . . _ _ _ __ _ _ _ _ 1.
''~"
, c= •••••••••• ••••••••••••• .... 4 , .t.,=
...
...
~... .
.
..".
~.~,
... " ...
,~F
1 #~c=:I
...
,..
'; 1 .~,--
.
· ,."., ... -,.,. t~
... : ...
r-,... ...
• • • • • • • • • • " I l . ..~?~
=)J
="
=.J
...
,..
'...
,....
...
,~...
;...
,
... .
·
l '
" :
1 : •.
Hi
I~
lei'
... 1 1., ....... ..•...•••.• .••. t
1
.
'.
'.
.
,
• • • • • • , . . . f t • • • • • • • • • • 1 '.~ ( , .ff I l • • • • • • • • il!' • • • • • • • • • • • • • • • • • · ...~
...~iIii"
_ _ ..t·!
J-J.\.
"'.
",
1 .1 11
1··
1 1i
~ 1 l ' " '.(
, , ... ~ .. '0""""
14 ',.
, ,Ce çOJDPlexe est traditionnellement décrit .comme étant la' zone de
0.5 cM. comprise entre les extrémités J( et D du chromosome. La partie
K étant à l,' extJ<émité ,centromérique et D à l'extrémité té1omérlque.. A
l ' i~térie'l:1r de ce 8egtDen~~ de~ régions et des sous-régions ont été
dé-~riées.
1 ·Ces régJ,.ons etsoua-ré8ion~ ~ont
définies pormne étantu~
seg-mènt 'de chromosome servant de locus pour 'au moins un' gène d
'histocom-\
.
',
patïbilité. Ell.es \peu.vent être
d~s.~ing,uées
entreelle~
parcrossif-g-over. Nous' ne savons toutefois pas' encore si un locus est le site de
\ '
, \
un ou de plusieurs gènes liés. Dans le complexe traditionnel, nous
comptOns cinq ré.~ions ~ K. I. S, G et D.
1.4.3 La région Tla
Là
région Tla du .génôme de 'la sour.1s est située sur le chrpmosome17. à la droite de l~ régior\. D- (Old et coll.. 1963). Cette portion du
,
-
,-chromosome cont.ient un cer'tain' nombre de gènes poss&dant l'information
t
~écessaire à la synthèse d'antig~nes de surface. Certains de çes antf-'.
,
gèOes...of,frent-'un intérêt particulier étant donne qu'ils semblent être'
~ ~--.._-- ~ . .
présents spéc~fiqueJl)ent à la surface.d'e certaines populations
cellulai-res 1ymphofdes. ceci la.issant supposer qu'ils pou'rFaient êtr;
d~s
él'é-. ;;e};ts importants dans la différenciation de
ce~ derni~res.
C'est la fP• r" ' .. ..
raison pour laquelle plusieurs immu~ologistes
,
incluent cette ,région,
~
dans le complexe majeur ~ 'histocompatibilité.
A l'heure 'actuelle, sept. loci ont été identifiés' dans cette région
(voir ,figure,l) . Quatre d'entre eux' codent pour ~es antigènes
déce1a-.
, .. 'bles sérologiqueDJent; i l s.' agit des loci t,la (OId et coll." 1963),
Qa-~ (Stanton et aoy~e, 1976). Qa-2 (Flaherty 1 1976) et Qa-3 {Flahert),'"
.
,..
, " ,..•
'-15.
.
, ~ li o •'et
coll...
M78a).~~ux' ~~tl:'~S \Od~t
Pour desantigenes imPortants
d~8.
~s cas de rejets de greffes de
:peau
et de tuméUra; 11 .'qi.t des loci...
, ~
H-3l'et H-32 (Boyse et coll., 1~70; F;aherty et B~~~t,' 1973;
Flaherty
'et Wachtel, 1975). Finalement. le p;roc:luit du loci H~2T semble
~pOT-tant dans ·.les :réactions de cytotoxicité à médiation éellulaire (Klein' .
et Chiang. 1978) •. 'D'e p.lus, deU?' autres
lo~i
codant pour les antigèn!!snommés Qa-4 et Qa-5, ont. 'été a~sociés à cette r~gloni m.tis leurs posi-'
.
tio1'ls sur le chromosome ne sont pas encore bien défl~ies (Baaer11.ng et
co,l1., 1979).
.
.
Apr~s ce'tte brève revue des connaisaances sur 'le"complexe maj eur
d' histocompatibilité, nous âllons. J!l_1ritenant entr~pr~ncl~e l'analyse 4u
'potentt~
qUé
possèden~
lesdifférènt~
Ipci de'ce complexe, à induire,
~. . :1" .
une ;éaction du greffon contre I,'hôte.
l ' .. 1
1 ~ ... • •
1.5. Lf.~nogénéti9ue de la réacdon- du' greffon èontre l'bSte
.' 1.5.1
tu
art~:(gènes' ~jeurs··d'h:l.stocompa~lbUit45
• Il'' est établi depuis lortg~e1DPé dans les' syst~~ murin et avlair~
que 'pour. obt'enir une réacqon GVH, i l fa~t que le donneur'~ et le recev~ur
so~ent h:l.s.~o1ncompaU.bles' (Bill1ngham et éoll., 1956; S'imon's.en, 1957
r'
~
Uphoff. 1958).~ 1
Q
Cependant t aucune étude n' a~liit ét' entreprise pour 8~01i: si leé
..
,différentes régions ou différents 1oc:l.S du c9mplexe .8':'2 p~uvaientti QU non
... 1 • • ",.
p_rovoquer des' réactions GVH av,ec plus ou lIloina d'intensité.
Les premier,s
à
f.aire ce genre d' ana1ya'~ ont été E1cliwald et colla-~ .'.porateurs en
~969
ètLengero~a
et Vikl1cky durant ,la iaêâe année. 'Ils, " ,
.
" • t,,1
1
L
·C
("-V
-(
(
, '.
16
ont tait leurs études chez la sour~s en ayant recours au modèle de l~
maladie de l'avorton que nous avons décrit précédemment. Ils ont
ten-•
té d'induire des réactions, GVH chez des nouvea~-nés en leur inj ~ctant
des cellu.les lymphofdes parentales selon différ,entes combinaisons. Ils
ont examiné les conséquences de la réaction ~ calculant dtx jours
après l ' in,duction,
1.'
ind~ce de spl.énom~galie. Cpntrairement auxrésul-tats at tendus, il~ 'concluen t en énon)an t qu'il ne semble pas 'y avoir.
de corrélation entre le nombre d'antigènes H-2 différents et l'intensité
d~ la réaction GVH obtenue. De plus, ils mentionnent que seulement:
certâ1ns,antigènes H-2 sont cap~bles d'induire une telle réaction et
même qu'une greffe (hl celluÎes' xéno'généiques ne provoque qu'une tr,ès"
faible spJ,.énomégalie. Tout éfoia. i,1s critiquent sévèrement leurs
tra-,;
vaux en expliquant que l'ind~ce de spléno~galie n'est peut-être pas
le critère idé~l pour ce genre d'étude génétique •
• 1
.
Ryc~ilkpvà,:t collaborateurs en 1970, en utilisant une technique
~us ,sen~,1b1.~'pour faire des études immunogénétiques. ont fait une
ob-.
,.
,sja~atiQn l , importante à propos de l ' influence des, régions K et D en ut'i-•
li,san t,une, réaction mixte lymphocytaire (RML). Ils ont noté que la région
. K pouvait ~timuler une réaction plus forte que la région D.· Dur,ant la
même année, Démant (1970) a démontré que ce nouveau concept s'appliqu@it
aussi à la réaction GVH, à savoir que la région K pouvait occasionber
u~e
té action' GVH plus intense que larégiO~
-'
Cependant, un peu ~lus tard.. certains chercheurs (Bach et~ (:611 .»
1972a,1972b) en reprenant ce 'genre d '~tud~, ont fait une mi,se au point
,. )
l '
.,
\
17
Rych:ll.kov' o~ encore Démant, pour étudiei l'influence de ...la' région, K,
,.
ils pouv,aient J;"etroùver ~n plus J certai~es autres disparité$ au niveau
de la région comprenant les gènes codant po~r ~ertains 'antigènes Ir.
Ils émettent alors 1 'hypothèse que 1.a différence ant1~énique perç\1e paT
une ~ entre les régions K et D était probablement causée par l'étroite
associati9n entre la région K et certains gènes Ir, donnant comme ré
sul-tat' que la région K prOvoquait une m.eilleure stimulation antigénJque que
.. ,IJ'."
la region D.
,.
Effectivement. en utilisant' de9 comhinaisons génêtiq\J~s p.lus
appro-priées, ils concluent ,que la région K seule
il'
offre pas une meilleure·stimulation antigénique que la région
D.
C~s résultats'Qnt d'ailleursété confirmes par K~ein ~t Park en 1973 qui se sont' aerv:[s de la
l::éac-tion GVH pour faire cette étude immunogenétique.
Les "résultats obtenus d~~a, ces études de l'influence des diffétents
.
antigènes du systèmé H-2 sur l"intensité des réactions;'observées. ont
"
.
...
souvent JQenés à des concl~sio~s contradictoi;res •. Ceci .étant occasionné
par des informations incomplètes'-concernant" ~ie génotype des sour1:S
uti-~i~ees, mais aussi par 1e fait ,que le"s réactions étudiées. différaient
\ '
au n1veliu de leur sensibilité. 0 Finalement, ces discordances' ont empêcha
l~s chercheurs' de faire rapidement une 8ynthè~e de l'influence des
anti-gènes H-2 dans l '-élaboration de réponses immunitair.es .
...
,'Klein et Park en 1.973 ont publié un article dans lequel 11s oht
réussi à faire une première classification des antigènes du systèllJe H-2
en fonction de leur pouvoir à induire des réac tions GVH d' inten$1t~
va-riable.
-(
" l j.
18~ C'est par l'utilisation de souris congéniq , et le modèle de la
maladie de l'avorton qu'ils ont fait cette 6tude. L'intensité de la
réaction
GVH
étant mêsurée selon l'indice de splénomégalie. Lesconc'lu--~
sions de ce travail se résument en c~nq points importants:
a), b)
c)
une différence au n,;1.veau de la région D cause unê faible GVH une différence au niveau de la région- K cause une GVH d'intensité
càDparable à' celle obtenue au ptlint a ,
une différence au niveau de la région codant pour les gènes Ir
provoque une GVH' de très forte intensité
.
.
d) une ~ifférence au niveau des loci Ss et Slp' ca?se. mais d'une
façon non 'c~nstante~ une GVH tle faible intensité
e) une différence de deux régions ou plus provoque une réaction GVH
. d'une intensité beaucoup plus grande q~e celle indurte 'par chacune
des r'égioqs prises indi:viduellement.
D'une
~nière spéculatiye, Ils supposent que les antigèneS" capablesd'.indu1re des 'réactions GVB seraient les mêmes que ceux qui stimulent
une'
RML
et qu'ils ;e retrouveraient à la surface des lymphocytes T.Opplto:va et DéDIant (1973) ont confirmé les résultats de Klein et
Park •. De 'Plus, ils se sont aU$si intéressés à l'influence d'un autre
1'0-cus du complexe H-2 soit le th. Malgr,é le fait qu'une incompat ibilUé ~
génétique au niveau du locus tla puisse provoquer le re.jet de greffe de
peau (Snell et coll., 1971; Démant et Graff, 1973), ils' ont prouvé que
,
r
19
Etant donne que de plus en plus d~ lignées congéniques de souris étaient disponibles, i l devenait alor:s possibl~ de compléter la classi-,fit:ation des antigèRes H-2 selon leur pouvoir de stimulation.
Klein et collaborateurs (1974a,,1974b ,1976~ 1977)>> Klein et Chiang en 1977 et Klein en 1977 ont démontré! que ,des différences au niveau des
'>
antigènes codés par la sous-région I-A étaient plus importantes pour
/ " 0 "'
l'induction de la réact~on GVH et le rejet d'allogreffes qu'une d'ifféren-ce 'au' niveau des antigènes cOdés' par
la
sous-région J;;-C. La sous-région "I-B ne semble pas contribuer à la stillD.llation antigénique lors des rej etsd,'allogreffes ni d'ailleurs dans l'induction de la réaction GVH.
Une observation intéressante copcernant les antlg~nes LAD a été résumée par Klein en 1978b. I l existe ce~taiites indications que toutes les régions et sou'S-régions capables de provoquer une réacti,?n GVlI
;!'
possèdent l ' informat.ion génétique codant p,our au moins un antigène LAD. I:l s'agit de LAD-3 dans la régio~ K, LAD-l et LAD-2 dans les
/sous-ré-, '. Bions I-A et l-C respectivement et dE! LAD ... 4 d~ns la région D. Cettj,e observation semble
vo~loir cQ~duire
à l'idée que les antigènes LAD~
seraien~ ceux r~sponsables dans l'induction de la réaction' GVH.
Tou-tefois à 1 'heure 'actuelle, cette pyp,othèse reste encore à être vérifiée, étant donné que des antigènes ,LAD ont_ été localisés dans la sous-région
Q
,
" l ' .-I-J et la région ,G et que pourtant .. une différence génétique au niveau de ces sites "n'induit pas de réa-ctiQn GVH.
En résumé, à l'intérieur du
~om~!exe
H-2 plusieurs antigènesp.~vent
provoquer l'induction de la réact1.on ,GVH~ Nous allons maintenant étudier si d r autres antigènes -à-l'extérieur du complexe ij-2 pourraient jouer l~même rôle. , \ ' \ 1
i
i,\
20
1. 5.2 Les antigènes mineurs d'histocompatibilité
La preuve était maintenant faite qu'une greffe allogénéique ~ntre un donneur et un receveur impliquant des loci sîtués da~s le Câmplexe majeur d'histocompatibilité conduisait à un rejet et dans le cas de greffes de moelle osseuse aboutissait fréquemment à l'induction cl 'une
~ réac tion GVH.
Certains chercheurs ont poursuivi leurs travaux dans le domaine de l'immunogénétique, mais cette fois-ei en utilisant des donneurs. et des receveurs identiques au niveau du système majeur d'histocompatibilité, afin de déterminer si d'autres antigènes extérieurs à ce dernier pou-vaient être impliqués dans ces phénomènes.
Les premiers trava~x ont porté essentiellement sur deux aspects. Il était déjà connu dans l'induction d'une réaction GVH contre des anti-gènes maj eurs d'histocompatibilité qu'une pré-sensibilisation des don-neurs par les antigènes des receveurs, de même que l'injection de. fortes doses de cellules lymphordes favorisaient un déclenchement rapide de la maladie et raccourcissaient le temps de survie des animau~' (Billingham et Silver, 1960; Billingham et coll., 1962).
Simonsen'';en 1962 décrit une expérience dans laquel~e i l a u tilif!é la pré-senSibilisation des donneurs afin -cl. 'induire une réaction GVH contre plusieurs antigènes mineurs d'histocompatibilité.
En se servant du système murin et du modèle de 1 'hybride ~ l ' i l a
i
tenté d'induire des réactions GVH en utilisant des cellules de ra tes pro-venant de parents pré-sensibilisés ou non aux antigènes de 1 ',hybride FI' Le critère qu'il a utilisé pour évaluer l'influence de lapré-sensibili-i 1" 1 1
1
1-/
....
sation est 'le poids de la rate meauré dix jours après l'injection des cellules. 11 a obten~ qu'en utilisant des donneurs non sensibilisés, le poids de la rate était inchangé éomparativement à des témoins lI.on-greffés • . En utilisant les. dç>nneurs pré-~ensibi1isés le poids de la
f rate était le double du poids des témoins', . Ces. résultats' ont par la
,
.
suite été confirmés chez le rat et le poulet (Elkins et Palm, 1966; Davis et Gole, 1967; Stre.iJ.ein et .Billingham, 1967;' Elkins. 1~70).
En ce qui concerne maintenant la dose, Simonsen en 1962, en utili-"Sant le système murin et le modèle de 1 '~ybrtde Fla réussi à prouver
qu'il obtenait une augmentation de plus en plus m~rquée du poids de la rate à" mesure que la dose de cellules lymphoYdes augmentaitJ Ces résul-tats ont été confirmés plu!? tard par Nisbet et Simonsen en 1967.
Sankowski et Nouza en 1968 et Nouza en 1971, en se servant de ces informations oot donè tenté d'induire des r~actions GVH contre des anti-gènes majeurs d'une ~art et mineurs, d'autre part, en util~sant ~e sys- ~
tème murin et le modèle de 1 l.hybride ~1' Ils ont étudié les éonséquen-ces' de ces réactions en faisant: des lpesures de poids de plusieurs organes .
,
comme la rate, le foie et le thymus à différents t~ps après l'induction. La conclusion de leurs travaux est la suivante: en utilisànt unediffé-~ence au niv,eau des antigènes majeurs~ ils observent de grandés
varta-tions du poids des org~nes des animaux en réaction G\TH comparativement ,
.
aux animaux témoins. Ils obtiennent aussi, quoique avec une intensitébeau~oup plus faible. les mêmes résultats en ut11isànt des disp.ar~tés
génétiques à l'extérieur du complexe H-2 •