Identification et caractérisation d'une PR10 impliquée
dans la synthèse de l'isobutylthiazole chez la tomate
Mémoire
Jérémie Ghironzi
Maîtrise en biologie végétale - avec mémoire
Maître ès sciences (M. Sc.)
Identification et caractérisation d'une PR10
impliquée dans la synthèse de
l'isobutylthiazole chez la tomate
Mémoire
Jérémie Ghironzi
Sous la direction de :
Résumé
L’isobutylthiazole est un composé volatil à l’arôme de verdure important pour la flaveur de la tomate. D’après sa structure moléculaire, il serait synthétisé à partir d’un composé azoté issu de la leucine et d’un composé soufré issu de la cystéine. Cependant, cette hypothèse n’a pas été confirmée et la voie de synthèse de l’isobutylthiazole reste méconnue. Ce projet avait donc pour objectif d’apporter de nouvelles connaissances sur le sentier du composé volatil. Un locus de 5 Mb sur le chromosome 4 a été ciblé pour son impact sur la production de l’isobutylthiazole. Une cartographie de précision a permis de réduire ce QTL à une région de 25 kb permettant ainsi d’identifier un gène candidat, appelé isobutylthiazole synthase 1 (IBST1). Le gène IBTS1 code pour une protéine de la famille des pathogénesis-related 10 (PR10), qui sont connues pour jouer des rôles divers chez les végétaux. La fonction du gène IBTS1 dans la synthèse de l’isobutylthiazole a été confirmée par transgénèse. L’ajout de précurseurs potentiels dans la tomate et dans les feuilles de N. benthamania agroinfiltrées a par la suite apporté des connaissances sur le sentier métabolique. Les composés azotés issus de la leucine ne permettent pas la synthèse de l’isobutylthiazole contrairement à certains composés non azotés issus du même acide aminé. Un enrichissement en leucine et en cystéine marqués a par ailleurs permis de confirmer que ces derniers sont les précurseurs de l’isobutylthiazole et que l’azote provient de la cystéine et non de la leucine. Un composé issu de la condensation entre l’isovaleraldéhyde, un composé non-azoté dérivé de la leucine, et la cystéine a été identifié dans la tomate, l’acide isobutylthiazolidine-4-carboxylique. Un enrichissement en acide isobutylthiazolidine-4-carboxylique a été réalisé dans les feuilles de N. benthamania exprimant IBTS1 et FMO, une Flavin monooxygénase responsable de la synthèse des composés azotés volatils chez la tomate. La combinaison des deux protéines avec le substrat a permis la production d’isobutylthiazole et des autres composés azotés volatils issus de la leucine. Cette étude a donc permis d’identifier un gène responsable de la synthèse de l’isobutylthiazole dans la tomate et d’améliorer les connaissances sur le sentier des composés volatils azotés issus de la leucine.
Abstract
Isobutylthiazole, which has a green aroma, is an important volatile in tomato that contributes positively to the flavor of the fruits. The biosynthetic pathway of this compound is still largely unknown. The most logical hypothesis is that isobutylthiazole results from the combination of a nitrogenous compound derived from leucine and a sulfurous compound derived from cysteine. This thesis goal was to verify this hypothesis and to improve our knowledge of the isobutylthiazole metabolic pathway. A locus of 5Mb on chromosome 4 was targeted for its high impact on the production of isobutylthiazole. Fine mapping allowed defining the locus to a smaller region of 25 kilobases containing only a single gene expressed in the fruit. The gene was named isobutylthiazole synthase 1 (IBTS1) and codes for a small protein belonging to the PR-10 group (Pathogenesis-related), a family of proteins with diverse functions. The role of IBTS1 in isobutylthiazole synthesis was confirmed by overexpressing and silencing the gene. Feeding experiments further allowed the identification of potential precursors in the metabolic pathway. Nitrogenous compounds derived from leucine did not result in higher isobutylthiazole, in contrast to the non-nitrogenous leucine-derived compounds. Labelled amino acid feeding experiment confirmed that leucine and cysteine are precursors of isobutylthiazole, and that nitrogen comes from cysteine instead of the leucine. A compound resulting from the condensation of isovaleraldehyde (leucine-derived) and cysteine was identified in tomato, isobutylthiazolidine-4-carboxylic acid. Feeding of isobutylthiazolidine-4-carboxylic acid was realized in agroinfiltrated N. benthamania leaves expressing IBTS1 and FMO, a flavin monooxygenase responsible for the synthesis of nitrogenous volatiles compounds in tomato. The combination of FMO, IBTS1 and the compound resulted in the production of isobutylthiazole and the other nitrogenous volatiles derived from leucine (isovaleronitrile, 3-methylbutanal oxime, 1-nitro-3-methylbutane). This study allowed to identify a new gene involved in the synthesis of isobutylthiazole and to expand knowledge of the synthesis of nitrogenous volatiles.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... viii
Liste des abréviations, sigles et acronymes ... ix
Remerciements ... xi
Introduction ... 1
Chapitre 1 Revue de littérature ... 3
1.1 La flaveur de la tomate ... 3
1.1.1 La flaveur ... 3
1.1.2 Les différents rôles des composés volatils ... 5
1.2 Les sentiers des composés volatils chez la tomate ... 6
1.2.1 Les dérivés d’acides gras ... 7
1.2.2 Les dérivés de terpènes ... 9
1.2.3 Les dérivés d’acides aminés ... 12
1.3 Identification de locus associés à l’isobutylthiazole ... 17
1.4 Hypothèses et objectifs ... 20
Chapitre 2 Matériel et méthode ... 21
2.1 Matériel biologique ... 21
2.2 Extraction des volatils ... 21
2.3 qPCR-RT ... 21
2.4 Plantes transgéniques ... 22
2.5 Production de protéine et purification ... 23
2.6 Activité enzymatique in vitro ... 23
2.7 Activité par enrichissement de substrat in vitro ... 24
2.8 Agroinfiltration ... 24
2.9 GC&GC-MS ... 24
2.10 Synthèse chimique ... 24
2.11 Analyses statistiques ... 25
3.1 Identification d’un gène candidat à l’aide des lignées d’introgression ... 26
3.2 Étude de la relation entre l’isobutylthiazole et l’expression d’IBTS1 ... 30
3.3 Confirmation de la fonction du gène IBTS1 ... 37
3.4 Caractérisation de la protéine IBTS1 ... 45
3.4.1 Test in vitro ... 45
3.4.2 Test d’enrichissement ... 46
Chapitre 4 Discussion ... 54
4.1 IBTS1 et la famille des PR10 ... 54
4.2 Le sentier métabolique des composés volatils azotés ... 56
4.3 Rôle de la FMO dans la synthèse de l’isobutylthiazole et des composés volatils azotés ... 58
4.4 La voie de biosynthèse de l’isobutylthiazole ... 60
Conclusion ... 65
Liste des figures
Figure 1 : Sentier métabolique des composés volatils issus des acides gras chez S.
lycopersicum.
Figure 2 : Sentier métabolique des composés volatils issus des terpènes chez S.
lycopersicum.
Figure 3 : Sentier métabolique des composés volatils issus de la phénylalanine chez S.
lycopersicum.
Figure 4 : Sentier métabolique des composés volatils issus de la leucine et de l’isoleucine chez S. lycopersicum.
Figure 5 : Utilisation de S. lycopersicum et de S. pennellii pour établir une cartographie de précision obtenue entre ces deux espèces.
Figure 6 : Émission d’isobutylthiazole et expression du gène IBTS1 dans les fruits des lignées d’introgression.
Figure 7 : Les huit stades de maturité du fruit de tomate (cultivar M82).
Figure 8: Quantité d’isobutylthiazole émise par les fruits du cultivar M82 aux huit stades de maturité.
Figure 9 : Quantité des composés volatils issus de la leucine émis dans la tomate du cultivar M82 aux huit stades de maturité de fruit.
Figure 10 : Quantité des composés volatils issus de l’isoleucine émis dans des fruit de la tomate cv. M82 aux huit stades de maturité de fruit.
Figure 11 : Quantité des composés volatils issus des terpènes émis dans les fruits de la tomate cv. M82 aux huit stades de maturité.
Figure 12 : Quantité des composés volatils issus des acides gras émis dans les fruits de la tomate cv. M82 aux huit stades de maturité.
Figure 13 : Quantité totale des composés volatils émis dans les fruits de la tomate cv. M82 aux huit stades de maturité.
Figure 14 : Expression du gène IBTS1 chez la tomate cv. M82 aux huit stades de maturité du fruit.
Figure 15 : Représentation de la structure tridimensionnelle de la protéine IBTS1.
Figure 16 : Quantité d’isobutylthiazole émis dans les tomates de la lignée témoin Vendor et de la lignée mutante IBTS1 P126S.
Figure 17 : Représentation de la structure tridimensionnelle de la protéine IBTS1 (à gauche) et de sa version mutante P126S (à droite).
Figure 18 : Étapes de production des lignées transgéniques.
Figure 19 : Expression du gène IBTS1 endogène dans les fruits transgéniques.
Figure 20 : A : Expression du gène IBTS1 synthétique ; B : Expression cumulée du gène IBTS1 endogène et synthétique des tomates transgéniques.
Figure 21 : Quantité d’isobutylthiazole émise dans les tomates transgéniques.
Figure 22 : Gel d’électrophorèse des protéines lors de la purification par Poly-Histag de la protéine IBTS1.
Figure 23 : Analyse UPLC-Q-TOF des tomates enrichies en leucine ou en cystéine marquées et non marquées et du témoin isobutylthiazole.
Figure 24 : Contenu en composés volatils après un enrichissement en acide isobutylthiazolidine-4-carboxylique dans les feuilles de N. benthamiana exprimant FMO et IBTS1.
Figure 25 : Contenu en composés volatils après un enrichissement en 2-isobutyl-1,3-thiazolidine dans les feuilles de N. benthamiana exprimant FMO et IBTS1
Figure 26 : Accumulation d’isobutylthiazole et présence du gène IBTS1 dans le clade
Lycopersicon.
Liste des tableaux
Tableau 1 : Séquences nucléiques et d’acides aminés du gène IBTS1 et du gène synthétique.
Tableau 2 : Quantités relatives des composés volatils émis des tomates (cv. M82) enrichies avec différents substrats.
Tableau 3 : Quantités relatives des composés volatils émis des tomates (cv. Large red cherry) enrichies avec différents substrats.
Tableau 4 : Quantités relatives des composés volatils émis des tomates (IL-4-1 et IL-12-2) enrichies avec différents substrats.
Liste des abréviations, sigles et acronymes
TomLoxC: Lipoxygénase 13
13-HPOT: Acide linoléique 13-hydroperoxide
ADH: Alcool deshydrogénase
IPP: Isopentenyl-diphosphate
FPP: Farnesyl diphosphate
GGPP: Géranylgéranyl diphosphate
TPS: Terpènes synthases
CCD1: Caroténoïde clivage dioxygénase 1
CYP (71ou79ou83): Cytochrome p450
FMO: Flavin monooxygénase
AAT: Alcool acyltransférase
QTL : Quantitative trait loci
IL : Lignée d’introgression
CAPS: Cleaved amplified polymophic sequence
IBTS1: Isobutylthiazole synthase 1
qPCR: PCR quantitative
PR10 : Pathogénesis-related 10
RT-qPCR: reverse transcription-PCR quantitative
AS: Antisens
LRC: Large red cherry
SE : erreur standard
UPLC: Ultra Performance Liquide Chromatographie
Remerciements
Premièrement, je tiens à remercier mon directeur de recherche, Charles Goulet pour son encadrement exceptionnel. En plus de m’avoir permis de travailler deux ans sur ce projet, il m’a appris dans les réussites comme dans les échecs à savourer la science et à déguster les tomates.
Puis je souhaite remercier tous les membres du laboratoire, Isabelle Clermont, Louis-Félix Nadeau, Blandine Bulot, Roberto Montoya, Sébastien Isabelle et Sylvain Villot pour leurs aides dévouées et les bons moments passés au laboratoire ou au champ.
Pour finir, je tiens à remercier Kassandra pour son soutien inconditionnel, ma famille et mes amis en France qui malgré la distance sont là pour moi.
Introduction
La tomate (Solanum lycopersicum) colore nos assiettes depuis longtemps. Ce fruit charnu est apprécié pour sa diversité de formes et de couleurs, mais surtout pour son goût unique. La tomate plait également aux agriculteurs pour ses qualités agronomiques. En 2016, la production de tomate fraiche a atteint 177 millions de tonnes, ce qui en fait la première production de légumes frais dans le monde (FAOstat, 2016). Une telle production a été atteinte grâce à des avancées de base en agronomie telles que la fertilisation, la protection des cultures, l’irrigation de précision et aussi grâce à des programmes de sélection efficaces pour développer de nouvelles variétés de tomates. Cette sélection et les avancées technologiques ont permis de multiplier par 2,6 le rendement de la tomate depuis une cinquantaine d’année (FAOstat, 2016).
Cette production intensive en champs ou en serre a occasionné une baisse de la qualité gustative. En effet, les consommateurs ont constaté une diminution de la flaveur des fruits depuis plusieurs années (Fernqvist et Hunter, 2012). Cette flaveur est le résultat d’une interaction entre les sucres, les acides organiques et les composés volatils (Baldwin et al., 2008). La saveur du fruit a longtemps été considérée comme secondaire par les sélectionneurs, car elle ne permettait pas une meilleure rentabilité pour la plupart des producteurs. Les sucres et les acides ont été intégrés dans le processus de sélection, alors que les composés volatils, plus complexes et dispendieux à phénotyper, sont généralement absents des objectifs des programmes de sélection (Yulinget Lindhout, 2007). La tomate produit plus de 400 composés volatils différents et une trentaine d’entre eux sont en quantité suffisante pour avoir un effet sur la flaveur du fruit (Baldwin, 2000). Dans les nouveaux cultivars de tomates, certains de ces composés volatils sont significativement moins accumulés dans le fruit comparativement à des variétés dites « anciennes » (Klee et Tieman, 2018). Parmi eux, l’isobutylthiazole, qui procure un arôme de verdure (céleri, persil ou encore feuille de tomate) et qui est primordial pour l’arôme typique de la tomate, est significativement moins produit dans les nouveaux cultivars. Ce composé est même utilisé dans l’industrie agroalimentaire afin d’intensifier le goût de tomate de certains produits de transformation (Maga et Katz, 1975). C’est un composé qui est unique aux espèces du sous-genre lycopersicon et qui s’accumule uniquement dans le fruit lors de sa maturation.
La tomate est le sujet de nombreuses études scientifiques, car il s’agit d’une plante modèle pour les fruits climactériques et d’une ressource importante dans la sécurité alimentaire. De
nombreux outils sont ainsi disponibles, tels que des banques de semences pour les plantes sauvages apparentées à la tomate domestique, des lignées d’introgression et une grande variété de cultivars et de plantes mutantes. Des outils génétiques majeurs sont également disponibles, comme le séquençage du génome (950Mb) de S. lycopersicum (génome complet), la tomate commune, ainsi que trois espèces sauvages apparentées, S. pennellii, S. lycopersicoides et S. pimpinellifolium (génome partiel) (The Tomato Genome Consortium, 2012). De plus, les génomes d’espèces diverses du genre Solanum ont été séquencés, c’est le cas de S. melongena et de S. tuberosum. L’étude des voies de biosynthèse des composés volatils chez la tomate est facilitée par l’accessibilité de différentes lignées d’intérêt, les données de séquençage disponibles et les nombreuses publications scientifiques sur le sujet. En pratique, la découverte d’un ou plusieurs gènes responsables de la synthèse de l’isobutylthiazole permettrait aux sélectionneurs de développer de nouvelles variétés avec une accumulation d’isobutylthiazole conservée au fil des croisements. C’est dans cette perspective que s’inscrit ce mémoire, ce qui a pour but d’améliorer nos connaissances sur la synthèse de l’isobutylthiazole.
Chapitre 1 Revue de littérature
La tomate est appréciée des consommateurs, notamment pour sa flaveur. Malheureusement, ce caractère est de plus en plus sujet à des plaintes pour les nouveaux cultivars de tomate. En effet, une étude a montré que les nouveaux cultivars sont moins savoureux et que certains composés volatils importants pour l’arôme sont parallèlement moins accumulés dans le fruit (Klee et Tieman, 2018). Une étude de la synthèse de ces composés volatils pourrait permettre de retrouver la saveur tant appréciée des consommateurs. Les paragraphes qui suivent dressent une synthèse des connaissances sur la flaveur des tomates et leurs les composés volatils, notamment ceux qui sont importants pour l’arôme.
1.1 La flaveur de la tomate
1.1.1 La flaveur
La flaveur particulière de la tomate résulte d’une interaction entre les sucres, les acides organiques et les composés volatils. Le goût (sucré, acide, salé, umami ou amer) d’un fruit est perçu par la langue, principalement par des cellules sensorielles qui possèdent des parois avec de nombreux chimiorécepteurs. Ces cellules sont reliées à des neurones qui transmettent l’information jusqu’au cortex cérébral. Par exemple, les protéines T1R sont des chimiorécepteurs localisés sur les cellules sensorielles qui enclenchent un signal électrochimique jusqu’au cerveau lorsqu’ils sont en contact avec des sucres (Li et al., 2002). Pour la perception de l’acidité, des chimiorécepteurs à protons H+ et à ions provoquent une dépolarisation de la cellule pour ainsi transmettre un signal (Da Conceicao Neta et al., 2007). L’arôme de l’aliment ingéré dérive quant à lui de la perception des composés volatils. La tomate, par exemple, en produit plus de 400 différents (Petró-Turza, 1986). Chaque composé volatil procure un arôme particulier (exemples : verdure, bois, floral) et l’ensemble de ces arômes procure l’arôme typique du fruit. L’intensité de la flaveur perçue est corrélée à la quantité de sucres et d’acides organiques. Dans la tomate, il est nécessaire d’avoir un équilibre entre les sucres et les acides pour avoir la flaveur typique de l’aliment même si les composés volatils sont en quantité suffisante. Entre autres, un manque d’acidité ou de sucres provoque une diminution de la flaveur (Stampanoni, 1994).
L’arôme est perçu différemment pour les sucres ou les acides. Il est détecté au niveau de la cavité nasale par voie olfactive ou par voie rétronasale. La voie olfactive permet de sentir
les odeurs par le nez tandis que pour la voie rétronasale, les composés volatils atteignent la cavité nasale lors de la mastication. Les composés volatils qui procurent l’arôme sont détectés dans la cavité nasale par l'épithélium olfactif (situé dans la partie supérieure) qui transmet l’information au bulbe olfactif puis au cerveau. L’épithélium olfactif chez l’humain est composé de 10 millions de neurones olfactifs et chaque neurone ne possède qu’un type de récepteur. Environ 400 récepteurs différents ont été répertoriés dans l’épithélium olfactif humain et chaque récepteur peut reconnaître plusieurs composés volatils différents qui donnent chacun un signal avec une intensité différente (Choi et Han, 2015). Cet ensemble de récepteurs permet à l’Homme de différencier plus d’un milliard d’odeurs différentes (Bushdid et al., 2016). L’odeur caractéristique de la tomate est induite par la détection d’une trentaine de composés volatils. La perception de la flaveur peut également être modifiée par des éléments externes. L’apparence, le toucher, la température de l’aliment et les souvenirs peuvent influencer la flaveur perçue par le cerveau (Verhagen et Engelen, 2006)
Plusieurs facteurs permettent l’identification d’un composé volatil par les neurones olfactifs. Dans un premier temps, il faut que les cellules olfactives expriment un récepteur protéique qui puisse reconnaître la molécule et transmettre un signal au cerveau. Par exemple, une étude a montré qu’une variation génétique mineure (polymorphisme d'un seul nucléotide) était présente dans la population pour le gène OR5A1, un récepteur olfactif pour la β-ionone (odeur de violette/framboise). Les personnes avec la mutation ne pouvaient pas détecter la β-ionone et donc l’odeur de violette ou de framboise (Jaeger et al., 2013). Dans un second temps, il faut que l’épithélium olfactif reçoive suffisamment de molécules pour que le cerveau transforme l’information en flaveur. Il existe un seuil de détection (ng/L) propre à chaque molécule, déterminé en pratique par un panel de personnes qui sentent une gamme de dilutions d’un composé volatil dans l’eau, jusqu’à un seuil de concentration la flaveur du composé n’est plus détectée. Par exemple, l’isobutylthiazole a un seuil situé entre 2 et 3,5 ng/L alors que pour la β-ionone, ce seuil est d’environ 0,007 ng/L. Dans la tomate ces composés sont présents à des concentrations d’environ 36 ng/L et 4 ng/L respectivement (Baldwin et al., 2000). Ces deux composés ont donc un impact dans la flaveur de la tomate.
Seulement une trentaine des 400 composés volatils produits par le fruit sont au-delà du seuil de détection (Buttery et al., 1989). Cependant, il semblerait que certains composés volatils ont un impact indirect sur la flaveur bien qu’ils soient émis en dessous du seuil de détection (Tieman et al., 2012). Les composés volatils de la tomate peuvent être divisés en trois
familles selon la nature de leurs précurseurs, soient les composés issus des acides gras, ceux issus des terpènes et ceux issus des acides aminés.
1.1.2 Les différents rôles des composés volatils
Les fruits font partie de l’alimentation de nombreux animaux, ils sont une source importante d’énergie, de minéraux, de vitamines et d’antioxydants (Willcox et al., 2010). La production de fruits est donc essentielle pour de nombreuses espèces animales frugivores. Mais les frugivores sont tout aussi importants pour les plantes. Parce qu’ils permettent l’endozoochorie, soit la dissémination des graines grâce à l’ingestion des fruits. Le frugivore, lorsqu’il se nourrit, ingère le fruit et ainsi transporte les graines dans l’environnement favorisant la colonisation du milieu par la plante (Sorensen, 1983). Les plantes ont développé des stratégies d’attraction pour leurs fruits, avec notamment la production de pigments rouges comme les carotènes pour rendre les fruits plus visibles ou encore la production de composés volatils pour augmenter la qualité gustative ou l’odeur du fruit. La tomate, avec sa couleur vive et sa production de centaines de composés volatils, fournit un exemple frappant de l’accumulation de ces molécules dans le fruit.
Un autre rôle connu des composés volatils chez les plantes est l’attraction des pollinisateurs. Contrairement à la tomate, de nombreuses plantes sont zoogames, c’est-à-dire que les animaux sont essentiels pour leur pollinisation. Pour attirer les pollinisateurs, la fleur produit des composés volatils qui vont être identifiés par les cellules olfactives de l’animal. Par exemple, dans la famille des Nicotiana, certaines espèces sont pollinisées par des papillons de nuit (Ippolito et al., 2004). L’attraction se fait par l’olfaction; les fleurs produisent davantage de composés volatils la nuit afin de se faire reconnaître et polliniser par les papillons nocturnes. Il s’agirait plus spécifiquement, des esters benzéniques et des oximes qui sont produites en grande quantité par la fleur des Nicotiana qui permettrait cette reconnaissance (Raguso et al., 2003). Les fleurs de tomate, sont pollinisées pour leur part par le vent principalement (Lesley, B.J.W., et Lesley, M., 1939) et produisent d’ailleurs peu de composés volatils (Buttery et Ling, 1993).
Les composés volatils sont connus également pour leurs rôles dans la réaction de défense des plantes. Ils peuvent dans un premier temps être nocifs ou répulsifs pour l’agent pathogène ou le ravageur. Par exemple, la fleur d’Arabidopsis thaliana produit de la β-caryophyllène, un composé volatil montre des effets d’inhibition de croissance contre une bactérie pathogène (Pseudomonas syringae) de la fleur (Huang et al., 2012). Le second
rôle des composés volatils dans la défense des plantes est indirect, via l’attraction de prédateurs. Un composé volatil avec un effet indirect a été découvert récemment chez
Oenothera biennis (Noge et Tamogami, 2018). Lorsque la plante est attaquée par le
coléoptère nuisible Altica oleracea, elle produit davantage de composés volatils. Le composé le plus synthétisé lors de l’infestation est l’isovaleronitrile, un composé issu de la leucine qui n’a aucun effet direct sur le coléoptère. Cependant, l’isovaleronitrile a un effet attractif pour Zicrona caerulea, un prédateur de l’A. oleracea. La punaise Z. caerulea, pour se nourrir, localise les plantes infestées par l’A. oleracea qui émettent de l’isovaleronitrile. L’émission d’isovaleronitrile permet donc aux plantes O. biennis de se défendre indirectement contre le ravageur (Noge et Tamogami, 2018).
Les composés volatils sont en outre des moyens de communication entre les plantes. Lors d’un stress biotique ou abiotique, la plante peut partager ce stress en produisant des composés volatils, qui peuvent être détectés par les plantes avoisinantes et engendrer des changements physiologiques en prévision d’une attaque éventuel ou d’un stress. Un des composés volatils les plus connus dans la communication plante-plante est le méthyl jasmonate qui est produit en plus grande quantité lors de certains stress. Ce composé volatil est détecté par d’autres plantes relativement proches qui de surcroit peuvent appartenir à des familles différentes. Dans une étude, il a été montré que le méthyl jasmonate produit par Artemisia tridentata de la famille des Asteraceae a augmenté par proximité l’expression d’un inhibiteur de protéase, une protéine de défense dans S. lycopersicum (Farmer et Ryan, 1990). Cette communication inter ou intra espèce rend les plantes plus adaptables à différente force de pression telle que la sécheresse, la salinité, les pathogènes ou encore les herbivores.
Les composés volatils ont ainsi une importance agronomique non négligeable. Ils permettent aux plantes d’attirer des pollinisateurs, de se défendre contre des stress abiotiques ou biotiques, et de communiquer avec les plantes avoisinantes pour parer ces stress. L’étude des composés volatils a donc une portée appliquée claire associée à l’amélioration de la qualité gustative, de la résistance aux stress et de la rentabilité via une meilleure pollinisation.
1.2 Les sentiers des composés volatils chez la tomate
Les composés volatils ont un fort impact sur la flaveur mais sont complexes à phénotyper et donc difficiles à inclure dans les critères de sélection. Ce désintérêt pour la flaveur a mené
à une baisse de production des composés volatils dans les nouveaux cultivars. Or l’identification des gènes responsables de la synthèse de composés volatils permet d’inclure plus facilement l’arôme dans le processus de sélection. Depuis l’apparition des appareils de quantification pour les composés volatils comme le chromatographe à phase gazeuse dans les années 1950, l’étude des composés volatils a été facilitée. Ces études ont contribué aux connaissances actuelles sur certaines voies métaboliques. Les trois voies métaboliques majeures pour les composés volatils, à savoir les dérivés d’acides gras, les dérivés de terpènes et les dérivés d’acides aminés, sont présentées ci-dessous.
1.2.1
Les dérivés d’acides gras
Les acides gras sont des molécules essentielles à toute forme de vie. Ils permettent principalement la formation de molécules structurales comme les phospholipides, mais jouent également un rôle dans les réactions de défense et l’imperméabilité des structures cellulaires. Un acide gras est une chaine aliphatique de 4 à 38 carbones, linéaire ou ramifié, munie d’un groupe carboxyle. Les acides gras sont des précurseurs de nombreuses molécules du métabolisme primaire, mais aussi du métabolisme secondaire tel que les composés volatils. Chez la tomate, une vingtaine de composés volatils sont issus des acides gras, dont certains montrent un impact positif sur la flaveur : (E)-2-pentenal (C5), 1-penten-3-one (C5), Hexanal (C6), Hexanol (C6), (Z)-3-Hexenal (C6), (Z)-3-Hexenol (C6), (E)-2-Heptenal (C7), 1-Octen-3-one (C8) et (E),(E)-2,4-decadienal (C10) (Tieman et al., 2017). Ces composés donnent des arômes différents selon leurs nombres de carbones. Par exemple, les molécules à 5 carbones (C5) procurent un arôme âcre ou noisette alors que les C6 procurent plutôt un arôme de verdure ou d’herbe coupée. Ces composés volatils permettent une meilleure flaveur des fruits et certains sont même impliqués dans les réactions de défense des plantes. La production de ces composés volatils se ferait lors de la destruction des tissus de feuille ou du fruit (Baldwin et al., 2000). Plus précisément, ils seraient produits à partir d’acides gras issus des membranes des thylakoïdes (Yashroy, 1987), qui elles sont constituées principalement d’acides linoléiques et linoléniques (C18). Ces deux acides gras sont des précurseurs connus ou potentiels des composés volatils chez la tomate (Figure 1). La première étape du sentier pour ces composés est l’ajout d’un groupe hydropéroxide (-OOH) sur l’acide linoléique ou linolénique via l’action d’une lipoxygénase. Par exemple, chez la tomate, la lipoxygénase 13 (TomLoxC) oxyde l’acide linoléique (18:2) sur le carbone 13 pour produire de l’acide linoléique 13-hydropéroxide (13-HPOT) (Chen et al., 2004). S’en suit une fragmentation de la molécule au niveau du carbone
porteur du groupe -OOH qui produit un aldéhyde à 6 carbones et un acide carboxylique à 12 carbones. Cette réaction est catalysée par une autre enzyme appelée hydropéroxidase
lyase (HPO-lyase) (Suurmeijer et al., 2000). L’Hexanal est ainsi formé, qui peut ensuite être
converti en hexanol par l’alcool deshydrogénase 2 (ADH2) (Speirs et al., 1998). La formation du (Z)-3-Hexenal/(Z)-3-Hexenol suit les mêmes étapes en partant cette fois-ci de l’acide linolénique (18:3). Le (E)-2-Hexenal est quant à lui produit par simple isomérisation, spontanée ou enzymatique du (Z)-3-Hexenal.
Si le sentier des composés à 6 carbones est bien connu, ce n’est pas le cas pour les composés à 5, 7, 8 et 10 carbones pour lesquels on ignore encore certaines étapes de synthèse. Chez le soya, les composés volatils C5 seraient produits à partir de 13-HPOT produit par une lipoxygénase. La forme ferreuse de la lipoxygénase permettrait une réduction du 13-HPOT et un clivage au carbone 13 pour produire un alcène à 5 carbones et un cétoacide à 13 carbones (Salch et al., 1995). Par la suite, une alcool déshydrogénase permet la formation du pentenol puis la production de 1-penten-3-one hypothétiquement par une autre ADH (Gardner et al., 1996). Par extrapolation les composés à 5 carbones chez la tomate pourraient également être synthétisés à partir de 13-HPOT produit par TomLoxC, mais il semblerait que la suite du sentier soit catalysée par d’autres enzymes que celles identifiées pour les C6. Dans le sentier des composés volatils issus des acides gras, seulement 3 familles d’enzymes sont possiblement responsables de la synthèse d’une vingtaine de composés volatils dans la tomate : les lipoxygénases, les hydropéroxidase lyases et les alcool déhydrogénases.
Figure 1 : Sentier métabolique des composés volatils issus des acides gras chez S.
lycopersicum. La zone en verte représente la voie de synthèse des C5 connue chez Glycine max L.. Lox : lipoxygénase, ADH : alcool déshydrogénase, HPO-lyase :
hydropéroxidase lyase.
1.2.2 Les dérivés de terpènes
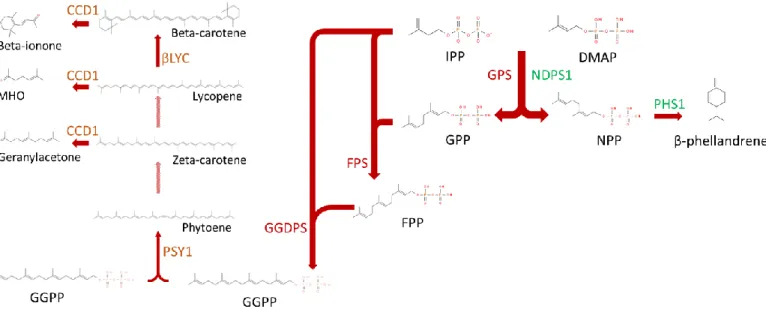
La famille des terpènes est la plus grande famille de composés organiques chez les plantes. Ils sont les précurseurs d’une grande variété de composés volatils comme les ionones, le linalool ou encore les phellandrènes, mais aussi de pigments comme le lycopène ou la β-carotène. Les terpènes sont utilisés dans de nombreuses industries pour leurs fragrances, leurs flaveurs, leurs colorations et leurs effets pesticides ou médicinaux. Chez les plantes, ces composés permettent une attraction des pollinisateurs et des frugivores, des communications plante-plante, et une protection contre des stress environnementaux comme les UV-B, les insectes ou les maladies (Gershenzon et Dudareva, 2007). Les terpènes sont classés selon leur nombre de doubles isoprènes (C5) ou leur nombre de terpène (C10) représentés ainsi (C10)n. Ainsi on retrouve les hémi- (C10)½, les mono- (C10), les sesqui- (C10)1½, les di- (C10)2, les sester- (C10)2½, les tri- (C10)3, les tétra-
(C10)4 et les poly- ((C10)n avec n>4) terpènes. Ces différentes molécules sont synthétisées à partir d’une réaction de condensation entre l’isopentényl-diphosphate (IPP) et un second diphosphate à x prényl (diméthylallyl diphosphate DMAP x=1 ; géranyl diphosphate GPP x=2 ; farnesyl diphosphate FPP x=3 ; géranylgéranyl diphosphate GGPP x=4). Certains terpènes ne sont pas produits à partir d’IPP, mais plutôt à partir de deux diphosphates à x prenyl, par exemple les squalènes (triterpène) qui sont produits à partir de deux FPP. Une fois la condensation effectuée, la molécule perd son groupe diphosphate pour donner un alcène à x isoprènes (Breitmaier, 2006). La molécule peut ensuite subir diverses modifications, comme des isomérisations, des oxydations, des clivages, des estérifications, des cyclisations ou des carboxylations. Par exemple, la famille des monoterpènes inclut des hydrocarbures comme le myrcène ou l’ocimène selon l’isomérisation, des alcools (ex : géraniol), des aldéhydes (ex : géranial), etc. La production de composés volatils à partir des terpènes est très diversifiée et implique de nombreuses protéines. La formation des prényls diphosphates se fait par une prényl diphosphate synthase appelée également prényltransférase. Ainsi, le géranyl diphosphate est synthétisé suite à la condensation de l’IPP et du DMAP par la géranyl diphosphate synthase (GPS). Selon la stéréochimie du prényl diphosphate produit par l’enzyme, la protéine sera classée comme une cis- ou une trans- prényltransférase. Une cis-prényltransférase produira du néryl diphosphate tandis qu’une trans-prényltransférase produira du géranyl diphosphate. Ensuite, les terpènes synthases (TPS) permettront la production de terpènes à partir des prényl diphosphates (Leon et al., 2013).
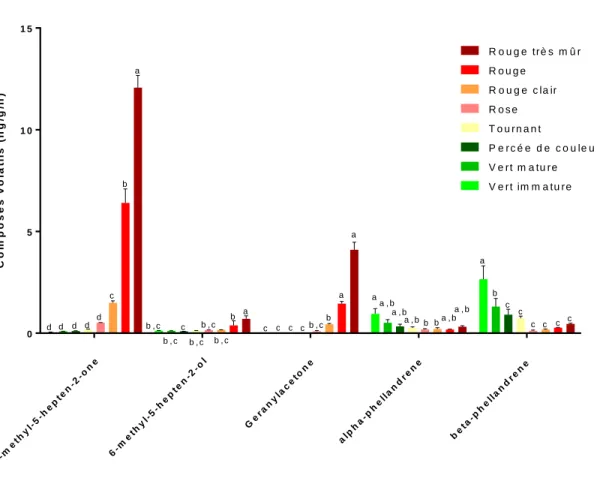
Dans le génome de la tomate comestible (S. lycopersicum), 40 gènes TPS ont été répertoriés, ce qui a permis d’élucider certaines voies métaboliques (Figure 2). Par exemple, la production du monoterpène β-phellandrène dans les trichromes commence par l’action d’une cis-prényltransférase appelé néryl diphosphate synthase 1 (NDPS1), qui catalyse la condensation de l’IPP et du DMAP pour former du néryl diphosphate. La protéine TPS20, ou PHS1 pour phellandrène synthase 1, permet à partir du néryl diphosphate la synthèse de la β-phellandrène (Falara et al., 2011). Les composés volatils terpénoïdes sont généralement des hémi-, des mono- et des sesqui- terpènes, parce que les terpènes avec plus de 3 isoprènes ont une masse molaire trop élevée pour être volatils. Chez la tomate, ces composés volatils sont retrouvés en faible quantité. Par contre, la tomate contient plusieurs composés produits à partir des caroténoïdes (tetra-terpènes,) dont la β-ionone (β-carotène), le géranylacetone (ζ-carotène) et le 6-méthyl-5-hepten-2-one (lycopène). Ces
composés sont reconnus pour faire partie des 30 composés volatils les plus importants pour obtenir la flaveur typique de la tomate (Tieman et al., 2017). Le tetra-terpène précurseur est synthétisé par la protéine phytoène synthase (PSY), une prényltransférase qui catalyse la réaction de condensation de deux GGPP pour produire du phytoène (Fray et Grierson, 1993). Le phytène subit des déshydrogénations (au niveau du carbone 12, 13, 20 et 21) et une isomérisation par deux réactions enzymatiques pour donner la ζ-carotène. Ensuite, par l’action de plusieurs enzymes, la ζ-carotène est déshydrogénée (sur le carbone 8, 9, 24 et 25) puis isomérisée pour donner du lycopène. Ce dernier composé peut être cyclisé aux deux extrémités par une lycopène bêta-cyclase (βLYC) pour produire la β-carotène (Ronen et al., 2000). Les carotènes et les lycopènes sont des pigments présents en grande quantité dans le fruit, qui leurs donnent sa couleur rouge. Cette couleur du fruit permet aux frugivores d’identifier visuellement les fruits. Ces composés colorés permettent également la synthèse des composés volatils essentiels identifiés précédemment. L’action d’une caroténoïde clivage dioxygénase 1 (CCD1) clive les tétra-terpènes pour produire des composés volatils aux arômes floraux ou fruités. La CCD1 clive la ζ-carotène au niveau de la double liaison covalente entre le carbone 6 et 7 et produit le géranylacétone, un composé volatil à l’arôme fruité et floral (rose, tropicale, magnolia, poire). La CCD1 clive le lycopène au niveau de la double liaison covalente entre le carbone 10 et 11 et donne le 6-méthyl-5-heptèn-2-one (MHO), un composé volatil à flaveur fruitée (pomme, citron, banane). CCD1 clive également la β-carotène au niveau de la double liaison covalente entre le carbone 9 et 10, ce qui aboutit à la β-ionone, un composé à flaveur fruitée (sucré, framboise) (Vogel et al., 2008). Les composés dérivés des terpènes jouent un rôle important dans l’attraction du fruit, autant visuellement que gustativement, par le biais de molécules très diversifiées.
Figure 2 : Sentier métabolique des composés volatils issus des terpènes chez S.
lycopersicum. Les acronymes en couleurs représentent des enzymes du sentier, ceux en
vert des enzymes de la voie de synthèse de la phellandrène, ceux en orange les enzymes impliqués dans le sentier des caroténoïdes et ceux en rouge des enzymes non spécifiques. Les flèches pleines sont des réactions caractérisées dans la tomate tandis que les flèches partielles sont hypothétiques. IPP : Isopentényl-Diphosphate, DAMP : diméthylallyl diphosphate, GPP : géranyl diphosphate, NPP : néryl diphosphate, FPP : farnésyl diphosphate, GGPP : géranylgéranyl diphosphate, MHO : 6-méthyl-5-hepten-2-one, GPS : géranyl diphosphate synthase, FPS : farnésyl diphosphate synthase, GGDPS : géranylgéranyl diphosphate synthase, NDPS1 : néryl diphosphate synthase, PHS1 : phéllandrène synthase, PSY1 : phytoène synthase, βLYC : lycopène beta-cyclase, CCD1 : caroténoïde clivage dioxygénase.
1.2.3
Les dérivés d’acides aminés
La troisième famille de composés volatils est issue des acides aminés. Les acides aminés sont connus pour être les constituants des protéines, mais ont aussi d’autres rôles pour les organismes vivants. Ils sont aussi une source de nutriments potentiels par anabolisme (C, H, O, N et S) (Braun et al., 2015) et les précurseurs de composés des métabolismes primaire
et secondaire qui interviennent dans le développement, la croissance, la reproduction et la réponse aux stress (Maeda et Dudareva, 2012). Les acides aminés ont une grande diversité de structures et de compositions chimiques et certains sont précurseurs de composés volatils. Dans la tomate, une dizaine des 30 composés volatils importants pour l’arôme sont issus de 3 acides aminés, soient la phénylalanine, la leucine et l’isoleucine.
1.2.3.1 Composés volatils issus de la phénylalanine
La phénylalanine est un acide aminé qui possède un cycle aromatique. Il s’agit d’un précurseur de composés volatils d’intérêt dans plusieurs plantes, retrouvés principalement dans les fleurs et les fruits. Dans la fleur du pétunia, par exemple, de nombreux composés volatils dérivés de la phénylalanine sont produits et sont impliqués dans l’attraction des pollinisateurs (Verdonk et al., 2005). Dans le fruit de la tomate, le 2-phényléthanol, le 1-nitro-2-phényléthane et le phénylacétaldéhyde sont issus de la phénylalanine et procurent un arôme floral (rose, jacinthe) important pour l’arôme caractéristique de la tomate (Figure 3). La phénylalanine peut être décarboxylée par une aromatique amino acide décarboxylase (AADC) pour produire du phényléthylamine. À partir de ce composé aminé, la formation de phénylacétaldéhyde se fait par une enzyme encore inconnue qui réalise une substitution du groupe amine par un groupe aldéhyde; il s’agirait hypothétiquement d’une amine oxidase (Tieman et al., 2006). Puis une autre enzyme, la phénylacétaldéhyde réductase (PAR) convertit le groupe aldéhyde en alcool pour produire du 2-phényléthanol (Tieman et al., 2007).
La voie métabolique pour les composés azotés issus de la phénylalanine n’est pas encore établie dans la tomate. En revanche, dans le peuplier, la voie de biosynthèse est connue. La synthèse du 1-nitro-2-phényléthane se fait via la formation du phénylacétaldoxime. En effet, un cytochrome P450 mono-oxygénases de la famille des CYP79 dans le peuplier a été caractérisé, cette enzyme convertie la phénylalanine en phénylacétaldoxime ou encore la leucine en 3-Méthylbutanal oxime (Irmisch et al., 2013). Par la suite, les composés aldoximes formés à partir des acides aminés permettent la formation de composés volatils avec un groupe nitrile ou nitro comme le 1-nitro-2-phényléthane (Schmidt et al., 2014). La conversion d’un aldoxime en nitrile par un autre cytochrome a été caractérisée dans certaines espèces et l’enzyme en question est une CYP71 (Vetter, 2000). Dans la tomate, aucune protéine CYP79 ou CYP71 ne semble être impliquée dans la synthèse des composés aldoximes, nitro et nitrile. Cependant, au sein du laboratoire une protéine a été identifiée comme importante dans la synthèse des composés azotés issus des acides
aminés. Cette protéine code pour une Flavin monooxygénase (FMO), une famille de protéine qui permet l’oxydation d’un azote ou d’un soufre. Cette protéine a été trouvée sur le chromosome 12 et permet la synthèse du phénylacétaldoxime, du 1-nitro-2-phényléthane, du 2-phénylacétonitrile et des composés azotés issus de la leucine. L’activité enzymatique et le substrat de la protéine ne sont pas encore caractérisés.
Figure 3 : Sentier métabolique des composés volatils issus de la phénylalanine chez
S. lycopersicum. Les flèches complètes représentent une réaction enzymatique
caractérisée pour la tomate, contrairement aux flèches partielles synonymes d’une réaction non caractérisée ou inconnue. AACD: aromatique amino acide décarboxylase, PAR: phénylacétaldéhyde réductase, FMO: Flavin monooxygénase.
1.2.3.2 Composés volatils issus de la leucine et de l’isoleucine
La leucine et l’isoleucine sont des isomères et sont proches chimiquement. La leucine a une ramification de type 3-méthylbutane alors que l’isoleucine a une ramification en 2-méthylbutane. Les composés volatils produits à partir de la leucine ou de l’isoleucine sont très répandus chez les végétaux et procurent des arômes très diversifiés. Dans la tomate, 9 composés volatils sont issus de la leucine et de l’isoleucine. L’acide isovalerique, le 3-méthylbutyl acétate, le 3-méthylbutanol, l’isovaleraldéhyde, l’isovaleronitrile, le 1-nitro-3-méthylbutane et l’isobutylthiazole sont issus de la leucine (Figure 4, A), tandis que l’isoleucine est le précurseur du méthylbutanal, du méthylbutanol et de l’acétate de
2-méthylbutyle (Figure 4, B). Les sentiers de ces 2 acides aminés sont similaires. La leucine, ou l’isoleucine, est d’abord transformée en cétoacide par une amino acide à chaine ramifiée
transaminase (BCAT), qui catalyse l’interconversion entre l’acide aminé et le cétoacide
(Taylor et al., 2004). Cette enzyme a été bien caractérisée dans le melon (Gepstein et al., 2010) et dans la tomate où des gènes candidats ont été identifiés et confirmés (Maloney, et al., 2010). Par la suite, le cétoacide est décarboxylé pour donner un aldéhyde par l’action d’une keto-acide décarboxylase (BCKA) (Kochevenko et al., 2012). La formation d’aldéhyde peut également se faire par une seconde voie métabolique à partir de l’acide aminé. L’acide aminé peut être potentiellement décarboxylé par une amino acide décarboxylase pour donner un composé amine qui peut ensuite permettre l’interconversion en aldéhyde. Cette seconde voie de synthèse a été observée uniquement pour la phénylalanine dans la tomate avec la caractérisation de la protéine Aromatique amino acide décarboxylase (AADC) (Tieman et al., 2006). Les composés volatils alcools, que ce soit pour le sentier des terpènes, des acides gras ou des acides aminés, sont synthétisés à partir d’aldéhydes par une alcool déshydrogénase (ADH). Cette réaction à double sens entre les alcools et les aldéhydes est en partie effectuée par ADH2. Chez la tomate, deux ADH ont été répertoriés : ADH1 et ADH2. ADH1 s’exprime dans les graines et le pollen alors que ADH2 est exprimé dans le fruit (Longhurst et al., 1990). Cependant il semblerait que des enzymes homologues à activité similaire soient impliqués, car un transgène antisens d’ADH2 ne permet pas de diminuer complètement tous les alcools (Pres et al., 1999). À partir des alcools volatils (issus de différents précurseurs) et d’un acyl-CoA, une enzyme appelée alcool acyltransférase (AAT) produit des esters volatils (Goulet et al., 2015). Ces esters ont des qualités de fragrance et gustative intéressante. Par exemple, à partir du 3-methylbutanol issu de la leucine (arôme de whisky) et de l’acétyl CoA, l’AAT catalyse une réaction de condensation pour produire dans la tomate du 3-méthylbutyl acétate (arôme de banane). De même, pour le sentier de l’isoleucine, la production de l’acétate de méthylbutyle se fait à partir du 2-méthylbutanol. Ces esters produits par l’AAT peuvent être reconvertis en alcool par une autre protéine, une carboxylestérase (CXE) (Goulet et al., 2012).
Dans le sentier de la leucine, trois composés volatils azotés sont importants pour l’arôme de la tomate : l’isovaleronitrile, le 1-nitro-3-méthylbutane et l’isobutylthiazole. L’isovaleronitrile et le 1-nitro-3-méthylbutane ont un homologue dans le sentier de la phénylalanine avec, respectivement, le 2-phénylacétonitrile et le 1-nitro-2-phényléthane. Dans le sentier de la phénylalanine, ces composés azotés semblent être corrélés au
phénylacétaldoxime et il se pourrait alors que le 3-méthylbutyraldoxime soit impliqué dans la synthèse de l’isovaleronitrile et du 1-nitro-3-méthylbutane. Dans la littérature sur la tomate, aucun lien entre une enzyme et ces composés azotés n’a été recensé. Cependant, l’enzyme FMO mentionnée précédemment permet la synthèse de l’isovaleronitrile, du 3-methylbutyraldoxime, du 1-nitro-3-méthylbutane et de l’isobutylthiazole. Il semblerait que FMO permet la synthèse des composés azotés issus de divers acides aminés. Cependant, aucun thiazole ne semble être issu de la phénylalanine.
L’isobutylthiazole est un composé volatil important pour l’arôme de la tomate avec sa flaveur de verdure de type persil, céleri ou encore feuille de tomate. L’isobutylthiazole est une molécule avec un groupe thiazole, c’est-à-dire un cycle à 5 atomes avec un soufre en position 1 et un azote en position 3 (Figure 4, A). Ce composé volatil a été découvert pour la première fois chez la tomate (S. lycopersicum) en 1969 et à ce jour, l’isobutylthiazole a été détecté uniquement chez le sous-groupe lycopersicon (Viani, R. et al., 1969). Cependant, d’autres thiazoles ont été identifiés dans la tomate, soient le 2-propylthiazole, le 2-sec-butylthiazole, le 2-isopropyl-4-méthylthiazole et le benzothiazole (Petró-Turza, 1986). L’isobutylthiazole semble être issu de la leucine, étant donné sa structure de type 3-methylbutane. Une expérience d’enrichissement en leucine marquée effectuée dans la tomate semblerait montrer que la leucine est le précurseur de l’isobutylthiazole. De plus, des corrélations ont été faites entre l’isobutylthiazole et des composés volatils issus de la leucine tels que l’isovaleronitrile, le 3-méthylbutanal et le 3-méthylbutanol (Buttery et Ling, 1993). Cependant l’autre partie de sa structure contient un soufre et des atomes de carbone supplémentaires, et il semblerait donc qu’un autre précurseur soit essentiel à sa synthèse. Plusieurs hypothèses dans la littérature convergent vers la cystéine comme second précurseur. En effet, de par la structure de la cystéine, il semblerait que la cystéine ou un dérivé soit le précurseur de l’isobutylthiazole (Schutte, 1974). Des composés volatils thiazoliques ont été recensés dans différents aliments comme le bœuf, le café, les arachides et les pommes de terre, mais ces composés ont été identifiés seulement après une cuisson donc produits de façon non enzymatique (Maga et Katz, 1975).
Il semblerait que des températures élevées permettent la formation de thiazole à partir d’une réaction de condensation entre les sucres et la cystéine (ou cystéamine), ou entre les aldéhydes et la cystéine (ou cystéamine). Une autre hypothèse avancée est la formation de thiazole par dégradation thermique de la thiamine (Schutte, 1974). Des thiazoles ont été détectés également dans des plantes de la famille des Brassicacées, comme Barbarea
vulgaris ou Arabidopsis thaliana. Ces thiazoles sont issus de la dégradation d’un
glucosinolate conjugué avec un acide aminé, cette dégradation forme un isothiocyanate puis un thiazole (Bednarek, 2012) (Agerbirk et Olsen, 2015). Cependant, les glucosinolates sont connus pour être produits presque exclusivement par les Brassicacées et aucun glucosinolate n’a été détecté chez les Solanacées (Rodman, 1991). Beaucoup d’inconnues persistent au sujet de la synthèse de l’isobutylthiazole. La découverte d’un ou plusieurs gènes clés du sentier permettrait potentiellement d’en savoir plus sur cette voie de synthèse.
Figure 4 : Sentier métabolique des composés volatils issus de la leucine (A) et de l’isoleucine (B) chez S. lycopersicum. Les flèches complètes représentent une réaction enzymatique caractérisée pour la tomate, contrairement aux flèches partielles synonymes d’une réaction non caractérisée ou inconnue. BCAT : amino acide à chaine ramifiée
transaminase, ADC : amino acide décarboxylase, KDC : keto-acide décarboxylase, ADH : alcool déshydrogénase, AAT : alcool acyltransférase, CXE : carboxylestérase, CYP71 : cytochrome P450 famille 71, CYP79 : cytochrome P450 famille 79.
1.3
Identification de locus associés à l’isobutylthiazole
L’essor de la biologie moléculaire a permis d’identifier plus facilement les régions chromosomiques associées à un caractère particulier. Plus spécifiquement, un suivi de la transmission génétique est possible grâce aux marqueurs moléculaires. Ces marqueurs
permettent d’identifier des portions du génome et d’établir des cartes génétiques, constituées de plusieurs marqueurs localisés sur l’ensemble du génome dans un ordre connu et avec une distance inter-marqueur calculée selon le taux de recombinaison. Chez la tomate, des cartes génétiques sont notamment disponibles pour des populations de lignées d’introgression créées avec des espèces apparentées à S. lycopersicum. Une lignée d’introgression consiste en un croisement entre deux lignées différentes et dont la progéniture est par la suite rétrocroisée avec l’un des deux parents. Le parent qui sert pour le rétrocroisement est le parent dit de référence; il s’agit ici de S. lycopersicum, la tomate commune. Le pool génétique du parent de référence est ainsi conservé en grande partie tandis seulement une fraction du génome du second parent est conservé. Les lignées obtenues diffèrent dans leur génétique, ayant conservé des fractions génomiques distinctes du second parent. Ainsi, les différences phénotypiques des lignées obtenues sont dues aux différentes sections génomiques du second parent. Pour l’isobutylthiazole, l’étude des lignées d’introgression est intéressante si les deux espèces parentales montrent des différences au niveau de leurs productions du composé volatil. En effet, ces lignées pourraient amener à l’identification d’un QTL, corrélés à la production de l’isobutylthiazole.
L’étude phénotypique de différente lignées d’introgression entre S. lycopersicum et des espèces apparentées : S. pennellii (Tieman et al., 2006), S. habrochaites (Mathieu et al., 2009) et S. pimpinellifolium (Rambla et al., 2017) a permis d’identifier plusieurs QTL corrélés à l’émission de composés volatils. Similairement, une population de lignées issues d’un croisement entre une tomate cerise et une tomate de plus gros calibre a été utilisée pour identifier des QTL associés à la flaveur (Saliba-Colombani et al., 2001). Pour l’isobutylthiazole, plusieurs QTL ont été identifiés. Les premiers, sur les chromosomes 2, 4 et 6 ont été corrélés à l’accumulation d’isobutylthiazole dans l’étude de Saliba-Colombani (2001). Des QTL ont été mis en évidence par une lignée de rétrocroisement entre les cultivars Cervil (S. lycopersicum var. cerasiforme) une petite tomate cerise à l’arôme intense et Levovil (S. lycopersicum) une grosse tomate avec un arôme commun de tomate. Le cultivar Levovil produit significativement plus d’isobutylthiazole que la tomate cerise Cervil, une différence corrélée à des régions spécifiques des chromosomes 2, 4 et 6 (Saliba-Colombani, V. et al., 2001). Des lignées d’introgressions entre S. pennellii et S.
lycopersicum utilisées par Tieman D.M. et al. (2006) ont permis d’identifier plusieurs QTL
reliés à des composés volatils. Le parent utilisé, S. pennellii, est une espèce apparentée à la tomate commune qui ne produit pas d’isobutylthiazole. Les données obtenues dans cette
étude ont permis d’identifier deux QTL pour l’isobutylthiazole sur les chromosomes 4 et 12 (Tieman et al., 2006). Le QTL du chromosome 4 a été confirmé par une autre étude portant sur une lignée d’introgression entre S. lycopersicum et S. habrochaites. S. habrochaites une espèce apparentée à S. pennellii produit des petits fruits verts au profil aromatique différent de la tomate commune. En particulier, l’isobutylthiazole est 10 fois plus accumulé dans la tomate commune que dans son espèce apparentée. L’étude a relevé une corrélation entre l’accumulation d’isobutylthiazole dans le fruit et la même région du chromosome 4 que dans l’étude de Saliba-Colombani (2001). Cependant, les QTL des chromosomes 2 et 6 n’ont pas été mis en évidence dans cette étude alors qu’une région du chromosome 5 induit une réduction de l’isobutylthiazole chez S. habrochaites (Mathieu et al., 2009). Enfin une troisième étude a montré une corrélation entre la production d’isobutylthiazole et trois nouvelles régions qui se situent sur les chromosomes 1, 3 et 9. Cette fois-ci, le matériel végétal utilisé provenait d’une population issue d’un croisement entre deux espèces produisant de l’isobutylthiazole, S. lycopersicum et S. pimpinellifolium. Cette dernière est une espèce proche de la tomate commune qui produit des petits fruits rouges comestibles (Rambla et al., 2017).
D’autres études ont adopté une approche différente pour identifier des locus. Au lieu d’utiliser des populations issues d’un croisement, ces études se sont basées sur la variabilité au sein d’une population de cultivars diversifiés. En utilisant des données de séquençage massif, deux études en particulier ont mené à l’identification de plus d’une centaine de QTL reliés à des composés volatils. La première étude, avec 174 cultivars différents a mis en évidence 4 locus pour la synthèse de l’isobutylthiazole situés sur les chromosomes 2, 6, 8 et 10 (Zhang et al., 2015). La seconde, avec une population de 398 cultivars a permis de corréler deux régions situées sur les chromosomes 5 et 11 avec l’accumulation d’isobutylthiazole (Tieman et al., 2017).
En somme, toutes ces études ont mené à l’identification de régions génomiques affectant l’accumulation de l’isobutylthiazole sur tous les chromosomes sauf sur le chromosome 7. Les QTL des chromosomes 2, 5 et 6 ont été identifiés dans deux études différentes et celui du chromosome 4 dans 3 études. Il y a donc plusieurs facteurs qui influencent l’accumulation du composé volatil et il n’est pas étonnant d’identifier plusieurs QTL pour un seul composé volatil étant donné qu’il peut y avoir plusieurs gènes clés impliqués dans la synthèse ou la régulation du sentier. De surcroit, l’isobutylthiazole est issu de la leucine et il se pourrait que certains QTL soient corrélés à la teneur en cet acide aminé dans la plante. Malgré
l’identification de tous ces QTL, l’identité des enzymes impliquées dans la régulation du contenu en isobutylthiazole reste inconnue.
1.4 Hypothèses et objectifs
Parmi les nombreux composés volatils produits par la tomate, l’isobutylthiazole est l’un des plus importants pour sa flaveur. Or, ce composé est significativement moins abondant dans les nouveaux cultivars. Afin de conserver des niveaux d’isobutylthiazole acceptables dans le fruit, il est nécessaire d’en savoir davantage sur sa biosynthèse. Pour l’instant, les connaissances sur le sentier métabolique de l’isobutylthiazole se résument aux sentiers de la leucine, avec laquelle il est corrélé étroitement. En s’appuyant sur la littérature, deux hypothèses ont été formulées pour ce projet :
1- Une région du chromosome 4 contient un gène clé du sentier de synthèse de l’isobutylthiazole chez la tomate.
2- L’enzyme responsable de la synthèse d’isobutylthiazole chez la tomate catalyse une réaction à partir d’un dérivé de la leucine et d’un dérivé de la cystéine.
Les objectifs poursuivis pour tester ces hypothèses ont été les suivants :
• Identifier un gène candidat impliqué dans la synthèse d’isobutylthiazole par cartographie génétique de précision.
• Vérifier la fonction de ce gène candidat dans la plante.
• Caractériser la protéine identifiée par des tests enzymatiques.
• Élucider les précurseurs de l’isobutylthiazole et les étapes potentielles du sentier métabolique menant à la synthèse de ce composé.
Chapitre 2 Matériel et méthode
2.1 Matériel biologique
Les lignées d’introgression IL4-1 et IL12-2 utilisées pour les travaux proviennent d’une population dérivée d’un croisement entre S. lycopersicum et S. pennellii (Eshed et Zamir, 1995). Les lignées pour la cartographie de précision ont été réalisées par rétrocroisement avec le parent M82 et en utilisant des marqueurs de type InDel et CAPS pour identifier les nouveaux recombinants. Une fois fixées, les lignées ont été cultivées dans le champ (de juin à octobre 2017, 2018 et 2019) ou la serre de l’Université Laval (Québec), au moins trois plants ont été plantés à trois emplacements différents et placés de façon aléatoire.
Les différents stades de maturité du fruit ont été sélectionnés ainsi : vert immature : coloration du fruit 100% vert avec des graines non viables ; vert mature : coloration du fruit 100% vert avec des graines viables ; Murissant : coloration du fruit 0 à 10% de jaune ou d’orange ou de rouge ; Tournant : coloration du fruit 10 à 30% de jaune ou d’orange ou de rouge ; Rose : coloration du fruit 30 à 60% de jaune ou d’orange ou de rouge ; Rouge clair : coloration du fruit 60 à 90% de jaune ou d’orange ou de rouge ; Rouge : fruit ferme et coloration de plus de 90% de jaune ou d’orange ou de rouge ; Rouge très mûr : fruit légèrement mou et coloration 100% de jaune ou d’orange ou de rouge (figure 5, p.29).
2.2 Extraction des volatils
L’extraction des composés volatils s’est faite à partir de 100g de tomate coupée en dés d’1 cm³ (issu d’au moins 3 fruits) placés dans un tube en verre. Le système utilisé permettait de créer dans le tube un flux d’air d’une extrémité à l’autre. Sur la sortie d’air du tube était placée une résine HayeSep Q qui capte les composés volatils. Après une heure d’incubation, les composés volatils étaient récupérés de la résine par une élution de chlorure de méthylène. Un standard interne était ajouté au solvant, le nonyl acetate. Pour les tests sur le tabac, les feuilles étaient roulées puis coupées en fines lamelles d’environ 1 mm de largeur.
2.3 qPCR-RT
Les extractions d’ARN ont été réalisées à partir de 220 mg de tomate broyée et congelée (à -80°C) sur les échantillons ayant servi aussi à l’analyse des volatils. L’extraction d’ARN était effectuée en selon les directives du protocole de Biobasic pour le kit d’extraction pour plante
avec une EZ-10 spin column. Une étape de digestion à la DNAse, puis une seconde purification, ont aussi été ajoutées. Les échantillons étaient dilués pour obtenir une concentration finale de 20 ng/μL. Les amplifications PCR en temps réel (qPCR) ont été réalisées avec le qScript One-Step qRT-PCR Kit de Quantabio (101414-172). Pour les lignées d’introgression et les différents stades, les amorces (GGGCTTGAAAGGTAAGTTGAG/TCAATGGCTTCAAACACTTCC) ont été créées afin de ne pas amplifier d’ADN génomique en plaçant une des deux amorces sur la jonction des deux introns. L’expression quantifiée d’IBTS1 endogène (Solyc00g060810) dans les plantes transgéniques a été réalisée avec une amorce dans la région 3’ UTR et une amorce dans
la séquence codante du gène (TGGGTTTTCTCATTGGTGTG/
CAATTCCTAGAAGGAGAGGGAGT). L’expression d’IBTS1 synthétique a été quantifiée avec des amorces propres à la séquence modifiée du gène (GGAAGCGTTATCAGGTGGAAG/CTTGTAAAGGAAAGTCCAGGTTG).
Gène IBTS1
ATG GGC TTG AAA GGT AAG TTG AGT GCT TCT ATA GAA ATA AAT TGT GGA GGA CAC TTA ATG TAT GAT TAT TTA TAC ACC AAA CCT CAT CTT ATA CCC AAC ATA ACA CCT AGC AAG ATA CTG AGT TTT GAG TTT ATC GAA GGC GAA ATC ATA AAG GTT GGT TCA GTT ATT AGG TGG AAA TAT AAC GAT GAT GGA AAA GAT AAG ATT GTA GAG GAA GTG TTT GAA GCC ATT GAT CCT GAA AGT AAA TCA ATC ACT TCG AAA ATA ATT GGC GGA GAT ATG TTA GAG CTG TAT GAT TCC TTC ACT TTT ATT TCG TCA AGT GAT CAG CAA TGG GCT ACA TGG ACA TTT CTG TAC AAA AAG AAA ACT GAA GAA ACT CCG GAA CCT CTT GCT CTA TTG GGT TTT CTC ATT GGT GTG ACA AAA GAT ATA GAG AAT CAC CTT CTC AAG TAA
Gène synthétique d’IBTS1
ATG GGA CTT AAA GGA AAA CTT TCT GCT TCT ATT GAA ATT AAC TGC GGT GGT
CAT CTC ATG TAC GAC TAC CTC TAC ACT AAG CCA CAC CTC ATC CCC AAC ATT
ACA CCT TCT AAA ATC CTC TCT TTC GAG TTC ATT GAG GGA GAA ATT ATC AAA GTT GGA AGC GTT ATC AGG TGG AAG TAC AAC GAC GAC GGA AAA GAT AAG ATC
GTG GAG GAG GTG TTC GAA GCA ATC GAT CCT GAG TCA AAG TCA ATC ACC TCT AAG ATC ATC GGT GGA GAT ATG CTT GAA CTT TAT GAT TCT TTT ACT TTT ATC AGC TCT TCT GAC CAG CAG TGG GCA ACC TGG ACT TTC CTT TAC AAG AAA AAA
ACT GAG GAG ACA CCT GAG CCT CTT GCT CTC CTT GGA TTC CTT ATC GGA GTT
ACC AAG GAT ATC GAG AAC CAC TTG CTT AAA TAA
Séquence d’acides aminés d’IBTS1
MGLKGKLSASIEINCGGHLMYDYLYTKPHLIPNITPSKILSFEFIEGEIIKVGSVIRWKYNDDGK DKIVEEVFEAIDPESKSITSKIIGGDMLELYDSFTFISSSDQQWATWTFLYKKKTEETPEPLALL GFLIGVTKDIENHLLK
Tableau 1 : Séquences nucléiques et d’acides aminés du gène IBTS1 et du gène synthétique.
2.4 Plantes transgéniques
La construction génétique pour les plantes transgéniques surexprimant la protéine d’intérêt a été développée artificiellement à partir du génome de référence. Les codons du gène ont été optimisés pour éviter l’induction de silencing tout en conservant la séquence d’acides aminés. La construction génétique a été insérée dans le plasmide pHK (Mülleder et al.,
2016). La construction antisens a été réalisée à partir du clonage de la séquence du gène d’intérêt issu de S. lycopersicum, cv. M82. La construction a été ensuite insérée dans le plasmide pHKAS. L’expression des deux constructions génétiques était régulée par le promoteur constitutif FMV 35S. S. lycopersicum cv. Large Red Cherry a été transformé par
Agrobacterium tumefaciens ABI à partir des cotylédons selon le protocole de Sun, H.J. et
ses collaborateurs (2006). La kanamycine a servi d’agent de sélection, à une concentration de 50mg/L. Une analyse quantitative des composés volatils et de l’expression du gène d’intérêt a été effectuée comme décrit précédemment.
2.5 Production de protéine et purification
La séquence codante d’IBTS1 a été amplifiée par PCR à partir de l’ADNc de S. lycopersicum
cv. M82, puis insérée dans le plasmide pDEST17 et pDEST15 avec le système Gateway.
Les plasmides ont été électroporés dans la souche d’Escherichia coli BL21 DE3 pour expression hétérologue. La culture bactérienne a été produite dans du milieu Luria-Bertani maintenu sous agitation à 28°C et avec comme antibiotique la carbenicillin à 100 μg/mL. Une fois la densité optique de 600 nm obtenue, l’expression de la protéine a été induite par ajout d’isopropyl β-D-1-thiogalactopyranoside à une concentration finale de 0,2 nM pendant une nuit à 28°C. Une lyse cellulaire a été effectuée par des cycles gel/dégel et l’ajout de 0,05% (v/v) de Triton-X-100 contenant du PMSF à 1% (p/v). Deux approches de purification ont été testé. Une purification « His-tag » a été réalisée avec les billes Ni-NTA agarose de Qiagen (Qiagen, 2003). La quantité d’imidazole dans la solution de lavage a été augmentée au fil des lavages (de 20 mM à 50 mM) jusqu’à l’élution avec une solution de 500 mM. Pour la purification par une glutathione S-transferase (GST) la fixation s’est faite sur des billes enveloppées de glutathion (Maity, R. et al., 2013). Après fixation, les billes ont été lavées avec une succession de tampons Tris-HCl pH 8,0 et PBS pH 7,0. L’élution a été réalisée à partir d’une solution de glutathion réduit à 10 mM. L’élution finale a été conservée à 80°C dans une solution de glycérol 25% (v/v).
2.6 Activité enzymatique in vitro
L’activité enzymatique de la protéine IBTS1 a été mesurée in vitro dans des tampons phosphate à pH 4 et 7. Les substrats ajoutés étaient à une concentration de 0,05 à 0,1 mM. Les produits ont été analysés en Headspace sur un GCMS (voir partie 2-19 GC&GC-MS) (Bais et al., 2003) (Samanani et al., 2004).