DEVELOPPEMEI'lI' FLORAL ET EMBRYOLOGIE
D'OCHNA
ATROPURPUREADC.
AESTRACT
PAUZE, Fernand. Floral Development and Embryo1o
gy
of Ochna atropur-pureaDe.
Department of Biology. Doctor of Philosophy. The inflor-escence, organogenesis, histogenesis, vascularization and embryology ofQ.
atropurpurea DC. are described. The 'centripetal' character of the sequential development of the androecial primordia is established for this species of the family Ochnaceae. The taxonomie significance of this morphological. observation is discussed in relation to the diagnostic importance attached· by Cronquist (1968) to the centripetal or centrifugaI character of the sequential initiation of the staminalprimordia~ in his phylogenetic classification of the dicotyledonso The validity of a universal application of this diagnostic criterium at the ordinal rank or at a higher level of classification is rejected. The conventional floral theories are not considered as satisfactorily explaining the morphological nature of the flower in
Q.
atropurpureaDe.
A new chromosome number (2n=
48) is reported. Fertilization by auto-pollination is demonstrated in this species. The conclusions reached by Chiarugi and Francini (1930) regarding partherlogenesis and ultimate propagation of the species through adventive embryony are rejected.Biology Dept.
PH.D.
RESUME
PAUZE~ Fernand. Développement floral et embryologie dlOchna
atropur-purea De. Département de Biologie. Doctor of Philosophy. Llinflo-rescence, l'organogénèse, llhistogénèse, la vascularisation et llem-bryologie d'O. atropurpurea DC. sont décrites. Le caractère du déve-loppement séquentiel Icentripète
' des primordiums de l'androcée est établi pour cette espèce de la famille Ochnaceaeo La signification taxonomique de cette observation morphologique est discutée en rapport avec 11importance diagnostique accordée par Cronquist (1968) au carac-tère séquentiel centripète ou centrifuge de 11 initiation staminale, dans sa classification phylogénétique des dicotylédones. La validité de l'app1ication universelle d1un tel critère diagnostiq~e, au niveau
ordinal ou à un rang de classification plus élevé est rejetée. Les théories florales conventionnelles sont considérées comme ne pouvant expliquer de façon satisfaisante la nature morphologique de la fleur
dl~. atropurpurea De. Un nouveau compte chromosomique (2n
=
48) est rapporté. La fécondation par auto-pollinisation est démontrée chez cette espèce. Les conclusions de Chiarugi et Francini (1930) quantà la parthénogénèse et à la propagation de l'espèce ultimement par embryonie adventive sont rejetées.
Aotlt, 1972.
UNE THESE
PRESENTEE A LA FACULTE
DES ETUDB3 GRADUES:; Er DE LA RECHERCHE: DE L'\UNIVERSITE MCGILL
COMME EXIGENCE PARTIELLE POUR LE DIPLOME DE "DOCTOR OF PHILOSOPHY" par FERNAND PAUZE
o
Fernand Pauze 19'73claircissements pour l'étude de la vascularisation;
3- c'est la permière fois que le développement inflorescentiel et floral est décrit chez une espèce de la famille des Ochna-~;
4- ces descriptions portent atteinte au système de classification phylogénétique de Cronquist (1968), qui divise les dicotylédo-nes en deux embranchements sur la base postulée de la centri-pétie ou de la centrifugie du développement séquentiel des pri-mordiums staminaux;
5- ces descriptions démontrent que les théories florales conven-tionnelles ne peuvent expliquer de faç"on satisfaisante la fleur d'Ochna atropurpurea
OC.
6- un nouveau nombre chromosomique est rapporté pour cette espè-ce déjà étudiée du point de vue embryologique, sous les noms d'O. multiflora OC. et d'~. serrulata Walp. en synonymie dl~.
atropurpurea OC.
7- certaines données embryologiques publiées dans la littérature sont démontrées comme étant erronées.
Remerciements
Nous tenons à exprimer notre profonde gratitude aux Profes-seurs du Département de Biologie de l'Université McGill, les docteurs Muriel V. Roscoe et Rolf Sattler, pour avoir proposé en 1965 les su-jets d'étude de cette thèse et pour leur constant intér@t concernant le succès de ces travaux. Le docteur
M.V.
Roscoe, qui devait trop h§tivement prendre sa retraite, a suggéré l'étude du développementCe dernier a inspiré l'auteur par ses écrits,
ses conseils et son encouragement.
Notre profonde reconnaissance est acquise aux autorités de
la Faculté des Sciences de l'Agriculture et de l'Alimentation de l'U-niversité Laval, ainsi qu'aux collègues du Département d'Agrobiologie et de Phytotechnie pour leur encouragement dans la poursuite de cette
recherche.
Nous adressons nos vifs remerciements au Professeur Jacques
Miège, directeur des Conservatoire et Jardin Botanique, Ville de Ge-nève , ainsi qu'à ses collaborateurs les docteurs S. Vautier et CoBoE. Bonner pour le contrele de l'identité de l'espèce à l'étude, dans les herbiers et la littérature. Nous les remercions également pour nous avoir fourni gracieusement la documentation pertinente et pour la con-servation dans leurs herbiers du spécimen que nous leur avons soumis.
Nous sommes reconnaissants au directeur des serres de
l'Her-barium de la Cité de Westmount {Province de Québec} pour avoir géné-reusement fourni quelques plants d'Ochna atropurpurea De. aux fins de cette étude.
Sur le plan de l'assistance technique, nous ne saurions trop remercier monsieur Jean
D.
Brisson, étudiant de deuxième cyclepour sa constance dans la poursuite de la revue bibliographique, pour son ingéniosité à dégager l'auteur de nombreuses sous-t~ches qui
au-raient nécessairement affecté le cours de cette recherche. Nous lui
devons le montage final des planches de cette thèse et leur dévelop-pement photographique. Notre profonde gratitude lui est acquise pour
son indéfectible collaborationo
Nous remercions également monsieur W. Hsiung, étudiant de deuxième cycle pour son assistance dans la préparation des cou-pes histologiques, monsieur Maurice Desmarais, professeur au dépar-tement d'Agrobiologie, pour le calcul au système APL/360 des taux de corrélations rapportés dans cette thèse, madame Louiselle Thériault, technicienne de laboratoire, pour la préparation des écrasements sur lames, servant au dénombrements des chromosomes et madame Francine Mondy pour la dactylographie de cette thèse.
Notre gratitude s'adresse au Conseil de la Recherche Agri-cole de la Province de ~ébec qui a permis à l'auteur, par ses
oc-trois annuels sous le numéro LA-67-299, de se procurer l'équipement, la matériel et l'assistance nécessaire à la poursuite de cette recher-che.
Finalement, l'auteur adresse un merci tout spécial à son épou-se pour son dévouement et son soutien moral, ainsi qu'à épou-ses deux fils pour leur patience et leurs encouragements.
AVANI'-PROPOS REMERCIEMENTS INTRODUCTION
MATERIEL El' MBI'HODES
1. Identification et origine de l'espèce 2. Méthodes
LE DEVELOPPEMENT FLORAL
1. Les théories florales
2. Concepts histogéniques floraux
3. Terminologie et symboles descriptifs 4. Organographie de l'inflorescence 4.1. Organographie du végétal
4.2. Modalité d'expression florale: la phénologie
4.3. Modalité d'expression florale: les inflorescences
4.3.1. Organographie d'une inflorescence de renfort
4.3.2. Expression florale d'une inflorescence de renfort
4.3.3. Type d'inflorescence: représentation d'une "aire inflorescentielle minimale"
5. Organogénèse florale
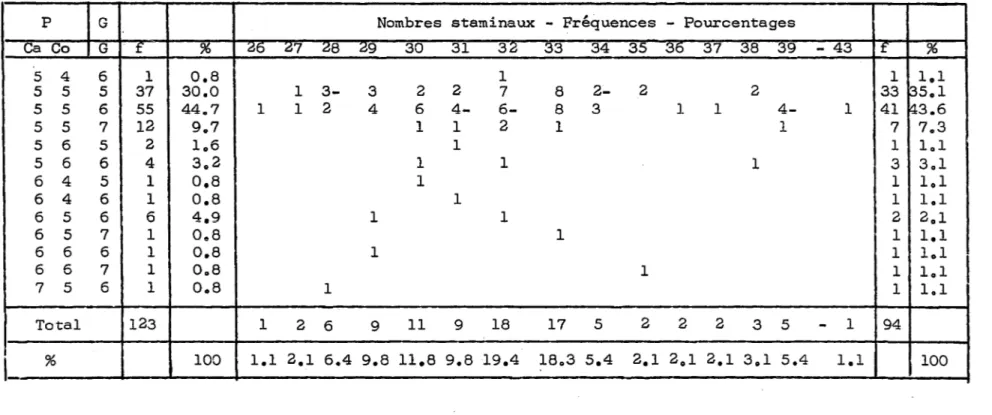
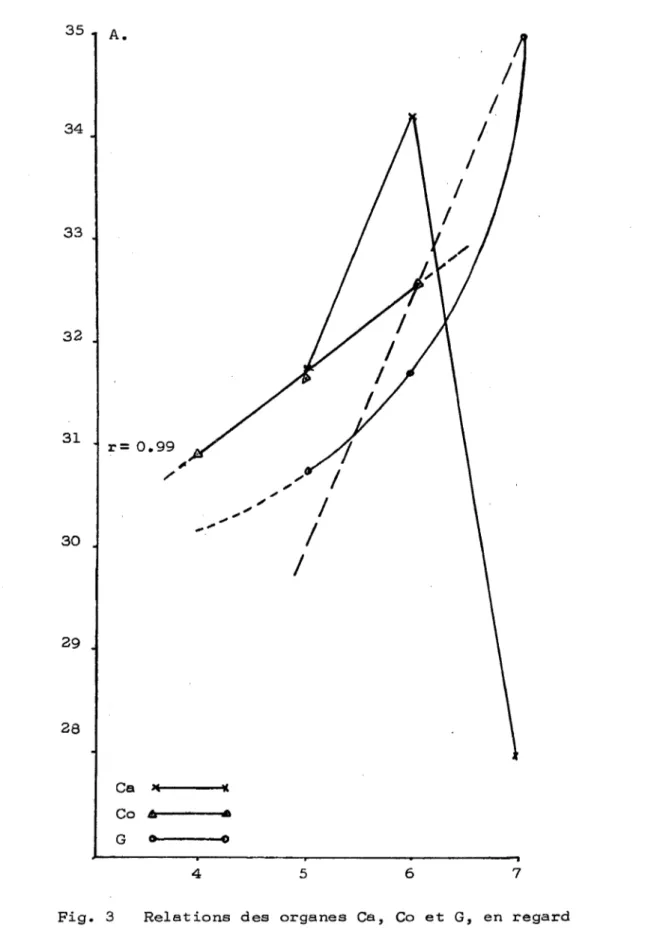
5.1. Description de l'organogénèse florale 5.1.1. Développement du périanthe i i i 1 10 10 13 18 18 26 32 36 36 39 41 42 44 47 63 63 63
5.1.2. Développement de l'androcée 65 5.1.3. Développement du gynécée 69 5.1.4. Variations dans la composition florale 73
5.2. Histogénèse florale 92
5.2.1. Apex inflorescentiel et membres périanthaires 92 5.2.2. Apex floral et membres de l'androcée 96 5.2.3. Apex floral et membres du gynécée 99 5.3. Apex floral et vascularisation 103
DISCUSSION 135
HABRYOLOGIE 165
1. Androsporogénèse, nombre chromosomique pollinisation et fécondation
1.1. Androsporogénèse
1.2. Nombre chromosom.iql.l'~ de 11 espèce 1.3. Pollinisation et fécondation
~. Gynosporogénèse, aposporie goniale
et aposporie somatique 2.1. Gynosporogénèse
2.2. Aposporie goniale et aposporie somatique
3. Gamétogénèse - Développement du gamétophyte - Anomalies
3.1. Gamétogénèse et développement du gamétophyte
3.2. Anomalies du développement d'un gamétophyte 167 167 169 171 172 172 174 175 175 177
4.1. Fécondation, amphimixie, apogamie 4.2. Les embryons et les graines
DISCUSSION
SOMMAIRE DES DISCUSSIONS ET CONCLUSIONS
BIBLIOGRAPHIE
181 186 190 200 227LNI'RODUCTION
Depuis son établissement par de Candolle, en 1811, la famil-le des Ochnaceae, d'origine presqufexclusivement pantropicafamil-le (Kanis,
1968), a fait l'objet de plusieurs travaux taxonomiques et phylogéné-tiques basés sur un nombre limité de caractères morphologiques, ana-tomiques et embryologiques. Parmi ceux-ci, les caractères exomorphi-ques floraux et ceux de la graine albuminée ou exalbuminée ont joué un rele déterminant dans la délimitation des cadres naturels de la famille. De fait, c'est à par·tir des particularités externes de la fleur et du fruit que de Candolle (1811, 1824) établit les principaux caractères de la famille originale composée, d'abord, des genres Ochna, Gomphia (= Ouratea), Walkera (1811), auxquels i l ajoute éven-tuellement les genres Elvasia et Castela ("Prodromusll 1824). Dans sa différenciation des genres Ochna et Gomphia, i l dis·tingue déjà les caractères suivants:
- Pétales 5-10. Etamines nombreuses, filets filiformes persistants, anthères linéaires ou ovales à double sillon et déhiscentes du som-met à la base. Bourgeons écailleux. Fleurs sortant sous les feuil-les, sur le bois de l'année précédente •••• 0 . 0 • • 0 0 • • • • • • • • • • Ochna
- Pétales 5. Etamines 10, filets subnuls, anthères longues, dressées en pyramide et déhiscentes par 2 pores apicauxo Grappes issues du
sommet des rameaux feuillés 0 0 • • • • • • • • • • • • • • • • • • • 0 0 . 0 . 0 0 • • Gomphia
Engler, dans son traité général des Ochnaceae (1874), fait la somme et l'épuration des travaux de ses prédécesseurs depuis de Candolle (1811, 1824), Planchon (1846-1847), Endlicher (1840), Bentham et Hooker (1862), Le Maout et Decaisne (1868), et Baillon (1873).
(1968, p. 2-7) et Farron (1968, p. 181-183) pour un relevé histori-que plus détaillé sur la taxonomie des Ochnaceae et des Ourateeae respectivement. C'est à Engler (1874) que revient le mérite de
réflé-ter les limites modernes de la famille. Il divise celle-ci en deux
groupes auxquels il n'accorde aucun statut formel; i l désigne ces grou-pes: série IIAlbuminosae" et série "Ex:albuminosae", en se basant sur la présence ou l'absence d'albumen dans la graineo Dans leur "Traité gé-néral de Botanique" (1876), Le Maout et Decaisne font, à grands traits, un relevé de l'état des connaissances acquises sur la feuille, la fleur à tous ses niveaux d'organes, le fruit, la graine et l'embryon. Ils résument ces données factuelles comme suit: "Sépales 4-5. Pétales autant que de sépales, ou en nombre dm/hIe. Etamines en nombre double ou multiple de celui des pétales. Anthères à déhiscence apicale. Car-pelles 4-5, ou plus, réunis par la base du style gynobasique, l-ovulés.
Fruit charnu. Graines peu ou point albuminées. Embryon droit. Tige
ligneuse. Feuilles alternes stipulées." - Dès lors, les caractères exomorphiquesgénéraux de la feo.mille sont bien connus 0 - Gilg reprend
ces caractères dans son traité taxonomique des Ochnaceae, dans le
IIDie natUrlichen Pflanzenfamilien" (1895), selon le système d'Engler
dont il conserve, jusqu'en 192 5, les séries "Albuminosae" et Ex:albu-minosae".
Van Tieghem (1902-1907), déjà familier depuis son "Traité
de Botanique" (1891) avec certains caractères inflorescentiels et flo-raux des principaux genres connus, apporte à la famille une nouvelle dimension sur le plan morphologique et une contribution très élaborée sur le plan taxonomique. Gr8ce à son sens aigu de l'observation, i l met en évidence de nombreux caractères anatomiques et morphologiques
1
3
de la tige et de la feuille (1902 .. 9,9) ainsi que de la graine et de l'embryon (1902 h) qui~ semble-t-il, ont échappé à ses prédécesseurso A l'aide des caractères tirés surtout de la graine et de l'embryon, i l propose une classification excessivement complexe de la famille (1902 g) qui révèle son concept étroit des taxons et son inorthodoxie dans le traitement des problèmes nomenclaturaux o Les auteurs subséquents lui préfèrent alors le système de Gilg pour la compréhension de la famille. Toutefois, Farron (1968) signale que les observations anatomiques et morphologiques de van Tieghem sont demeurées très utiles pour la déter-mination des relations interspécifiqueso
Decker (1966) rapporte une vingtaine de travaux, s'échelon-nant de 1889 à 1959, portant presqueexclusivement sur l'anatomie du bois de un ou de plusieurs genres de la famille des Ochnaceae. Pritzel
(1898) et Martin (1946) étudient l'anatomie de la graine, Cousin (1953) s'attache à llanatomie de la feuille,
Baum
(1951) étudie l'apocarpie du fruit d'Ochna multiflora DC., Erdtman (1952) travaille sur le pollen. A l'exception des travaux de Decker (1966) sur l'anatomie du bois des Luxemburgieae et des familles alliées, la morphologie des Ochnaceae est davantage étudiée, par ricochet, par l'ensemble des auteurs sui-vants: Sinnott et Bailey (1915), sur l'anatomie nodale et la morpholo-gie des stipules; Vestal (1937), Dickison (1967), respectivement sur l'anatomie du bois des Hype:r'icaceae et des Dilleniaceae; Wilson (1937), Corner (1946), sur la phylogénèse et sur la centrifugie des étamines respectivement; Schofield (1968) sur l'anatomie pétiolaire desGuttif~ et des familles alliées. C'est sur une partie ou sur la
majorité de ces travaux que les systématiciens se sont basés pour re-grouper, coordonner et relier entre eux les éléments offrant le plus d'affinités naturelles.
En 1925, dans le "Die natUrlichen Pflanzenfamilien:i Gilg place les Ochnaceae parmi les familles considérées les plus primitives dans le grand plexus des Parietales d'Engler (1909). Corner (1946) con-sidère ce plexus comme un regroupement vague de différentes familles qui n'ont aucun caractère en commun, pas m&me la placentation (la
"supposée" placentation pariétale, selon Dickison, 1967), et que ce groupe n'a de naturel que la "concaténation". Lawrence (1951) signale également que l'ordre des Parietales n'est pas phylogénétique et qu'il faut s'attendre à ce que les familles soient réparties dans plusieurs ordres. Un relevé des principaux systèmes de classification~ ainsi que
l'examen des travaux de Vestal (1937), Lawrence (1951), Kanis (1968),
et d'autres, révèlent que les Ochnaceae ont été placées, selon les auteurs, dans les ordres suivants: Geraniales (Bentham et Hooker, 1862), Guttales (Hallier, 1905, 1912), Guttiferales (Bessey, 1915; Wettstein, 1924, 1935; Rendle, 1925; Pulle, Compendium, 1937; Melchior, 1964), Parietales (Gilg, 1925; Lawrence, 1951; Chadefaud et Bmberger, 1960), Theales (Hutchinson, 1926; Takhtajan, 1959, 1969; Cronquist, 1957, 1965, 1968; Thorne, 1968), Clusiales (Pulle, Compendium, ed. 3, 1952), Ochnales (Hutchinson, 1959).
Bien que Hutchinson (1959) soit le seul à élever la famille au rang ordinal, i l ressort de cette liste que deux ordres prévalent actuellement: celui des Guttiferales (Melchior, 1964) et celui de Theales (Cronquist, 1968; Takhtajan, 1969). Le système de Thorne
(1968) n'est connu que par un synoptique. Ces deux ordres, constitués "sensiblement" des m&nes familles, réflètent en substance l'ancien sous-ordre des Theineae d'Engler élevé au rang ordinal sous deux noms diffé-rIants, selon les conceptions phylogénétiques différentes des auteurs.
5
(Pour les Theales, seul le nom de Cronquist sera retenu dans la pré-sente étude. Cronquist, 1968, reconna~t qu'il suit Takhtajan dans son
arrangement synoptique des sous-classes des Magnoliatae). - Dans chacun des deux ordres, la famille des Ochnaceae est placée parmi les familles considérée les'plus primitives et, è l'intérieur même de la famille, le genre Ochna est considéré le plus primitif.
Selon Melchior, l'ordre des Guttiferales est naturel mais plastique. Au niveau des sous-ordres et des familles, les recoupements et les caractères communs sont si nombreux qu'une étroite corrélation parentale ne peut faire de doute. Pour ces raisons, i l devient impos-sible d'établir des délimitations nettes entre les différents sous-ordres. De par les nombreux caractères primitifs qui le caractérisent, cet ordre est très près, et de façon très directe, des Magnoliales. -Un déyeloppement parallèle de ces deux ordres est alors postulé. Basé principalement sur la présence ou l'absence d'endosperme ainsi que sur les caractères de l'axe floral et des organes qu'il porte, l'ordre est divisé en 4 sous-ordres et 16 familles. Chacun des sous-ordres compor-te les familles suivancompor-tes:
1- Dillenieae (seul sous-ordre où les familles sont réparties d'après la centrifugie ou la centripétie des étamines):
a- étamines centrifuges: Dilleniaceae, Paeoniaceae, Crossosomataceae b- étamines centripètes: Eucryphiaceae, Medusagynaceae,
Actinidia-ceae
2- Ochnineae: Ochnaceae, Dioncophyllaceae, Strasburgeriaceae, Diptero-carpaceae
3- Theineae: Theaceae, Caryocaraceae, Marcgraviaceae, Quiinaceae, Guttiferae
4- Ancistrocladineae: Ancistrocladaceae
Les Guttifera.les de Melchior sont donc reliées aux Magnoliales par sui-te de leur degré très élevé de similitude et ces deux ordres sont
in-terprétés, sur la base des caractères morphologiques et phylogénétiques connus, comme ayant évolué parallèlement dans le grand complexe rana-léen dlEngler. Ce complexe est généralement considéré primitif au sein
d'un système de classification de conception plus ou moins naturelle (phylogénétique).
En contraste, les Theales de Cronquist sont nettement sépa-rées du complexé ranalé"èn et davantage reliées au complexe parfois
appelé guttiféraléen. Dans son arrangement synoptique, Cronquist (1968) divise les Magnoliatae (classe dicotylédone) en 6 sous-classes don~ la
séquence phylogénétique est la suivante: Magnoliidae (Renales sensu Engler), Hamamelidae, Caryophyllidae, Dilleniidae, Rosidae, et
Asteri-~. Il dérive les Theales des Dilleniales, ordre de base de la
sous-classe Dilleniidae. Il dérive également les Ochnaceae des Dillenia-~, hypothèse que Dickison (1967) supporte. Cronquist (1968, p. 191) suggère que "le seul caractère par lequel les Dilleniales diffèrent des Magnoliidae c'est la séquence centrifuge du développement des étamines
"
chez les Dilleniales (traduction libre). Il indique clairement (1968, p. 195) que chez les Theales, "les étamines sont nombreuses et centri-fuges, etc •• o" Il importe donc, pour fins de comparaison dans la dis-cussion de la présente étude, de signaler les familles constituantes des Dilleniales aussi bien que des Thealeso
- Dilleniales: Dilleniaceae, Paeoniaceae, Crossosomateceaeo
- Theales: Ochnaceae, Rhopalocarpaceae, Sarcolaenaceae,
Marcgra-7
viaceae, Quiinaceae, Elatinaceae, Medusagynaceae, Guttiferae.
Le postulat de Cronquist (1968), à l'effet que les familles des Theales ont un androcée à développement séquentiel centrifuge, ré-suIte sans doute, en partie du moins, des travaux de Corner (1946).
Celui-ci rapporte, qu'en autant que connu, l'androcée est centrifuge dans six familles: Hypericaceae, Cistaceae, Dilleniaceae, Bixaceae, Loasaceae; i l postule qu'il pourrait en gtre ainsi chez: Ochnaceae, Caryocaraceae, Guttiferae et Flacourtiaceae. Il ajoute de plus, à l'ap-pui de ce postulat, que l'androcée est centrifuge chez les Opontiales alliées.
,
De l'analyse bibliographique qui précède, il ressort: -- que
les positions taxonomiques et phylogénétiques de la famille des
Ochna-~ ont été établies principalement sur la base des caractères
exomor-phiques floraux, et sur un nombre restreint de caractères anatomiques et morphologiques concernant surtout la tige, la feuille et la graine;
que parmi ces caractères, certains sont di~s primitifs et d'intérgt
scientifique sur le plan phylogénétique; -- que les affinités de la famille, du niveau infra-familial au niveau ordinal, sont incertaines et problématiques ën raison, sans doute, du défaut d'une information suffisante et, comme corollaire, qu1il y a peu d1entente sur la posi-tion phylogénétique de la famille; -- que les postulats de Corner (1946)
et de Cronquist (1968) attendent encore leur vérification dans les ob-servations factuelles, quant au développement centrifuge probable des étamines chez certaines familles des Theales dont les Ochnaceae; --que la confusion actuelle résulte, en partie, du man--que pres--que absolu d'information sur l'ontogénèse et l'histogénèse des structures florales; -- finalement, que l'étude du développement floral chez Ochna
atropur-pure~ DC. pourrait apporter une contribution partielle importante à
l'élucidation de la position phylogénétique de la famille des Ochnaceae. Pour ce qui concerne l'embryologie des Ochnaceae, Davis
(1966) ne cite que cinq auteurs, reliés à six travaux portant exclusi-vement sur le genre Ochna. Les auteurs cités sont: van Tieghem
(1902 c, g), Francini (1928), Chiarugi et Francini (1929) 1930), Chia-rugi (1931), Chikkanniah (1954), et Ghosh (1964). - Les publications de ces deux derniers auteurs ne sont que des "notes" annonçant l'expo-sé éventuel des résultats de leurs études sur les espèces
o.
squarrosa L. etQ.
atropurpurea De. respectivement. - Toutefois, la littérature est encore dépourvue des résultats annoncés et, depuis 1964, aucune autre publication n'a pu 8tre relevée sur l'embryologie d1autres espè-ces ou de genres de la famille. Van Tieghem (1902 c, g) traite de l'anatomie de l'ovule et décrit les caractères morphologiques et de po-sitions des cotyles et de l'embryon dans la graine. Les travaux de Francini (1928), Chiarugi et Francini (1929, 1930) et Chiarugi (1931) ne se rapportent, en substance, qu'à une seule entité pour laquelle deux noms d'espèces furent utilisés,soitQ.
multiflora DC. (Francini, 192 8) et O. serrulata Walp. (Chiarugi et Francini, 1930). La situation est rectifiée dans l'exposé de 1930, et le nomQ.
serrulata Walp. est con-servé. En 1931, Chiarugi ne fait que rapporter les conclusions de la publication de l'année précédente. Quant au nom de l'espèce retenu, i l sera démontré dans la description du matériel utilisé pour la présen-te étude que le nomQ.
serrulata Walp. est réductible à celui dl~. ~purpurea DC. selon la loi de la priorité de désignation.
-Les travaux de Chiarugi et Francini (1930) constituent une étude caryologique et embryologique très poussée, mettant à jour pour
9
la première fois chez une même espèce, selon Chiarugi (1931), les phénomènes de l'aposporie somatique et de l 1 embryologie adventive re-liés par des formes de transition (proliférations sporophytiques). L'espèce est rapportée (1930, 1931) comme pentaplofde et à 2n
=
35 chromosomes. - Ces résultats sont les seuls rapportés pour la famille des Ochnaceae dans les importantes revues de Stebbins (1941) et NYgren(1954, 1967) sur la question de l'apomixie chez les Angiospermes. L1étude embryologique de l'espèce ~. atropurpurea DC. était
déjà amorcée quand llauteur du présent travail commença la traduction de l'imposante publication des Italiens: "Apomissia in Ochna serrulata Walp." (1930, 250 p.).
La
ressemblance des caractères embryologiques du matériel à llétude avec le matériel décrit et illustré par les Italiens était si manifeste qu1elle devait conduire à la connaissance éventuelle de la synonymie de ces deux espèces. Toutefois, certaines différences, probablement significatives, étaient déjà décelées dans notre matériel. Il fut donc décidé de poursuivre cette recherche qui pourrait peut-être révéler des différences plus marquées et conduire à des connaissances nouvelles sur le comportement embryologique de lles-pèce étudiée.En conséquence, les impératifs de la présente recherche dé-coulent clairement de 1 1 analyse bibliographique qui constitue llessence de cette introduction, et l'objet de l'exposé qui va suivre sera alors d'une double nature, soit: le développement floral et l'embryologie d'Ochna atropurpurea DG.
1. Identification et origine de l'espèce.
Le matériel utilisé pour la présente recherche était cultivé, jusqu'à récemment, sous le nom d'Ochna multiflora OC. dans les serres de l'Université McGill, et de la Cité de We~ount (Montréal, Canada).
Le matériel de ces deux serres est d'apparence identique et provient d'une même origine ultime comme i l sera indiqué un peu plus loino
Tel que signalé dans l'introduction, le doute soulevé concernant la similitude de notre espèce avec celle décrite par Chiarugi et Francini (1930) n'était plus permis. Un spécimen portant fruits, fleurs', feuilles et bourgeons est alors envoyé pour fins d'identification au Professeur Jacques Miège, directeur des Conservatoire et Jardin Botani-ques, Ville de Genève. Ce sont les Ors S. Vautier et C.B.B. Bonner qui contr$lent l'identité de l'espèce dans les herbiers et la litté-rature. Ils signalent, dans une communication personnelle reçue en juillet 1969, qu'en ce qui concerne le spécimen vérifié "il ne s'agit pas en effet de l'Ochna multiflora OC. dont les feuilles sont beaucoup plus grandes et plus larges, et dont les inflorescences racémeuses, allongées sont multiflores (12-18)". Voici, en citatIon, le texte de Vautier et Banner qui permet une rectification: ••• "Le nom d'Ochna serrulata Walp. (Repertorium, Tome 5, p. 400, publié en 1845-1846) a été en réalité repris par l'auteur du Diporidium serrulatum Hochstett décrit dans un article de Krauss: "Catalogus plantarum a Dr. Krauss in Colonia capensi et terra natalensi collectarum" publié dans Flora Vol. 27/1, p. 304, en 1844. Cette espèce est citée par Harvey dans Flora capensis de Harvey et Sonder, vol. l, p. 448, en synonymie de l'Ochna atropurpurea OC. publié par A.-P. de Candolle en 1811 dans
I l
les Annales du Muséum d'Histoire Naturelle de Paris, Tome 17, p. 412. Cette espèce, de de Candolle, a donc priorité sur le nom de Hochstett (1844).11
Vautier et Bonner joignent aux renseignements ci-dessus une photocopie d'une reproduction de l'Ochna atropurpurea OC. (Tabula 1392) dessinée par Ann Webster (1962), avec texte et description de Naomi van der Watt. Cette reproduction est tirée de "The Flowering Plants of Africa, Vol. 35, Parts 3 and 4, 196211. Pour ce qui est de la description faite par van der Watt, elle se l i t comme suit (traduit de l'anglais): "Arbuste, 3-5 pieds de hauteur. Branches couvertes de lenticelles soulevées, d'apparence squameuses,et visibles à l'oeil nu. - Feuilles subsessiles ou pétiolées (9-13 mm de long), allant jusqu'à 5 cm de longueur et jusqu'à 2.0 cm de largeur, habituellement ellipti-ques, occasionnellement elliptiques-ovées, elliptiques-oblongues, ellip-tiques-lancéolées, rarement lancéolées; obtuses, arrondies ou s'amin-cissant légèrement à la base, avec bords marginaux
à
dents serrés habi-tuellement triangulaires, dressées et à pointe aigu~, rarement incur-vées; nervure médiane fortement soulevée, nervures latérales légèrement soulevées. Fleurs solitaires à l'apex de rameaux courts, rarement deux. pédicelles 1-1.5 cm de long, articulés à la base ou à environ l mm de la base. Sépales 6-7 mm de long, 3-6 W~ de large, elliptiques,elliptiques-ovés, elliptiques oblongs, arrondis à l'apex, agrandis et de couleur rouge foncé dans le fruit. pétales 0.9-1.3 cm de long, obové avec base cunéiforme. Filaments 2-2.75 mm de long; anthères 'pl us longues que les filaments 2-3.5 mm de long, oblongues ou d' dspec t
linéaire, s'ouvrant par de courts pores terminaux. Ovaire de 5-G carpelles; style 5-7 mm de long; stigmaTe en nombre égal aux carpelleso
Fruits noirs, subglobuleux, 7 mm de long, environ 5 mm en diamètre; 1-2 ou plusieurs drupes, aussi larges que des pois, sur un torus hémi,;-phérique. (Herbier National, Prétoria, No. 29012.) - Planche 1392, F.P.A. Décembre, 1962.11
Vautier et Bonner signalent p~core q~en consultant la c1ef des espèces de Gilg dans l'article 1I0chnaceae africanae" (dans Bot. Jahrbuch f. Syst. Vol. 33, p. 232-236, 1903) et celle de E.P. Philipps (Bothalia, Vol. l, p. 90, 1922), ils arrivent à~. atropurpurea DCo qui se distingue par des lenticelles denses visibles à l'oeil nu, etco, tel que décrit par van der Watt. Ils concluent que l'ensemble des ca-ractères décrits parart s'adapter parfaitement à notre spécimeno
Chiarugi et Francini (1930) décrivent la distribution d'O. atropurpurea DCo et d'~. ~rrulata Walp. comme suit (traduit de
l'ita-lien): "Géographiquement, Ochna atropurpurea est répartie dans l'Afrique Méridionale, de près du Cap de Bonne Espérance (District de Calédonie)
jusqu'à la partie sud-orientale du Transvaal (près de Barbeton). Ochna serrulata est confinée à un angle de la limite orientale du grand ter-ritoire de l'espèce affine. Elle vit dans la forêt littorale du Natal près de la baie du Natal (Durban) où elle fut récoltée pour la première fois par Krauss en 1839. De cette localité, elle fut rapportée à Kew en 1861 par Sanderson, résident anglais à Durban et correspondant de W. Hooker. C'est de Kew que l'espèce fut diffusée à travers le monde sous le nom erroné de Ochna mul tiflora".
De cette dernière traduction, il ressort que l'espèce italien-ne correspond bien à la citation antérieure de V~utier et Bonner, que les espèces de Florence et de Montréal proviennent toutes deux de Kew et, en dernière analyse, de l'Afrique du Sud. Les noms d'espèces ~.
13
mu1 tif10ra DC. (Montréal) et Q.. serru1ata Wa1p. (Florence) sont donc réductibles à Ochna atropurpurea DC.
Dans une lettre du 24 octobre 1969, le directeur du Conser-vatoire de Botanique de Ville de Genève, le professeur Jacques Miège, assure l'auteur du présent exposé que le spécimen reçu pour fins d1i-dentification est conservé dans les collections de Genève et désigne: 1 plante cultivée, collecteur Fernand J. Pauzé. Un autre spécimen est déposé à 11Herbier p. Louis-Marie o.c.s.o. de l'Université Lava10
L'iden·t:ité de l'espèce à 1 l étude est donc rétablie, ce qui aura des implications dans la seconde partie de cette thèse, soit llembryologie.
2- Méthodes.
Les méthodes utilisées pour la réalisation de ce travail so~t
variées en raison même des aspects différents traités. Du c~té floral,
ce sont 1lorganogénèse, l'histogénèse et l'étude de la vascularisation.
Du c8té embryologique, i l s'agit: a) de l'étude du développement mor-phologique des microspores et des mégaspores jusqu'à la formation des embryons dont le développement structural est vérifié dans les grai-nes, b) de la vérification de la germination des grains de pollen !lin vitro" et de leur comportement dans les styles et les ovules, c) de la germination des graines pou.r le dénombrement chromosomiqlle dans les racines. Les techniques utilisées varient selon les sujets traités.
1- Concernant llorganogénèse florale, le matériel est pré-paré selon la techniqu~ de Sattler (1968).
2- Pour l'histogénèse des axes inflorescentiels et floraux, les procédés habituels de l'inçlusion à la paraffine et de la microto-misation sont utilisés (Johansen, 1940; Sass, 19(Y1). Toutefois, da.ns
ce cas, le tétrahydrofurane (THF) remplace l'alcool butylique tertiai-re (T.B.A.) pour la déshydratation u Les petits axes floraux sont préa-lablement insérés dans de petits blocs rectangulaires de tige de chou-fleur, de 3 à. 5 mm en diamètre, et colorés à l'éosine pour fins de ma-nipulation et d'orientation de la coupe au microtome. Le matériel est sectionné ent~e 7 et 10 microns d'épaisseur selon la dureté des maté-riaux. Une fois étalées et séchées sur lame de verre, les sections sont reCO<.lvertes de "Cello2dinll à. 1.5% pour prévenir leur décollement. Les combinaisons de colorants utilisées sont: safranine - Fast Green (Johansen, 1940, p. 80-82), safranine - bleu d'aniline (Gurr, 1965,
pu 305), acide tannique - chlorure ferrique - safranine (Poster, 193~,
p. 91). Cette dernière combinaison s'avère supérieure pour la colora-tion des parois cellulaires des tissus méristématiques de notre maté-riel. Cependant, dans le cours du travail, elle n1est adoptée que tar-divement et sur une faible quantité de matériel u
3- L~ vascularisation est étudiée dans les coupes
anatomi-ques et par l'éclaircissement d'axes floraux entiers selon la techni-que de Rodin et Devis (1967).
4- Pour l'étude embryologique, les fixateurs employés sont variés et tels que décrits par Johansen (1940: pu 39-45): la solution de Carnoy; les solutions alcool - acide acétique glacial dans les proportions 1:1, 3:1; la solution ac:i.de chromique - acide acétique glacial - formaline (CRAF) de Navashin. Comme colorant, l'hématoxy-line régressive de Heidenhain est d'abord utilisée. Mais en raisün de la présence d'une très grande quantité de plastes amylifères prenant fortement le colorant dans les sacs embryonnaires matures, ainsi que la forte chromophylie des synergides pour l'hématoxyline, l'examen des
15
structures devient impraticable et une quantité appréciable de maté-riel et de temps fut perdue à truvers les tentatives d1amélioration de la technique, mais sans su~cès appréciables. L'hématoxyline de
Harris ou celle. de Delafield (Johansen, 1940) ont alors remplacé la précédente avantageusemen·t~ à condition d'éviter la surcoloration et
d10btenir une différenciation assez poussée du colorant avant le mon-tage final des lames. Le matériel est sectionné habituellement à 18
microns d'épaisseur, à cause des dimensions des cellules du sac embryon-naire qui varient entre 15 et 25 microns d'épaisseur pour les cellules de 1 'appareil oosphère~ ou davantage pour les antipodes o
5- Pour la culture "in vitro" des grains de pollen, la tech-nique utilisée est celle de Brewbaker et Kwack (1963), et pour llétude du comportement des tubes polliniques dans les styles et les ovules la technique de Martin (1958) est adoptée permettant les observations au moyen de la fluorescence.
6- Dans le cas de la germination des graines pour le dénom-brement chromosomique ~ans les racines, les drupes matures sont récol-tées directement sur les frui ts et n'2 doi vent pas subir de périodes de dessèchement marquées, sans quoi la germination ne peut sleffectuerg Les graines sont libérées de leur péricarpe par dissection et déposées sur buvard humide dans des bo~tes de plastique transparent. Ces bot-tes sont placées en lumière dans un germoir mai ntenu à la température constante de 250 Cu Les racines atteignent 1 cm et plus de longueur entre 4 et 7 jours de germination. La méthode suivie pour le comptage des chromosomes est celle de Salesses (1967) pour les arbres fruitiers à noyaux et à nombre chromosomique élevé, mais légèrement modifiéeo Comme prétraitement, le présent auteur utilise une solution aqueuse
de colchicine à 085% durant 4 heures. Le réactif de Schiff ou bien
le Peulgen sont employés comme colorants pour une période de 8 à 12
heures à 4 o C. La période d'hydrolyse à l'acide chlorhydrique IN à
600 C d'une durée de 10 minutes donne ici les meilleurs résultats.
La macération est effectuée en deux étapes, 2 heures dans une solu-tion de pectinase à 20% suivie d'une autre période de 2 heures dans une solution à 0.1% de cellulase (Worthington Biochemical COrp. Free-hold, New Jersey, rU/mg). Cette deuxième étape est une autre modifi-cation apportée à la méthode de Salesses, en raison des difficultés rencontrées dans l'étalement des cellules en couche monocellulaire.
La
libération d'une certaine quantité de cellulose des paroiscellu-laires permet des aplatissements convenables des grosses cellules à
cytoplasme très dense qui ne se réalisent autremen't qu'avec
difficul-tés.
7- PoUr l'étude et la photographie du matériel aim,i prépa-ré par les différentes techniques, les systèmes optiques suivants sont
u'tilisés. L'épiscopie pour l 'organogénèse selon la technique de Sattler
(1968) à l'aide d'un condensateur pour lumière incidente SU.L photomi-croscope Zeiss avec objectifs à c8nes d'immersion de Leitz (4.0X et 6.5X) adaptés pour le condensateur Zeiss. Afin de tirer parti adpqua-tement de l'objectif 4.0X, le présent auteur a ajouté dans le
porte-filtre une lentille achromatique (grac;i.euseté du département d~
physi-que de la Faculté des Sciences de l'Université Laval) ayant une dis-tance focale de plus ou moins 10 cm. L'addition de cette lentille a permis un apport lumineux convenable pour l'examen et la photogrùphie d'inflorescences trop grosses pour l'objectif 6.5X. - Dans le cas des sections faiblement colorées et difficiles à résoudre en diascopie,
17
l'auteur s'est largement servi de l'optique du contraste de phase ordinaire, et celui du contraste de phase interférentiel - différen-tiel avec condensateur de Nomarski. - Finalement, pour l'étude de la fluorescence des tubes polliniques dans les styles et les ovules, l'emploi d'un microscope Wild M-2Q équip~é à cet effet s'est avéré efficace.
LE DBVELOPPEMBNT FLORAL
1. Les théories florales.
La morphologie florale a toujours été et demeure la pierre d '.achoppement de la morphologie végétaleo Aussi, la question perpé-tuelle: "qu'est-ce qu'une "fleur"?" demeure-t-elle toujours d'actuali-té. C'est cette question que différents concepteurs et théoriciens tentent d'élucider de façon lia priori", par suite de l'absence de té-moins fossilisés, seuls détenteurs authentiques possibles de l'histoire du développement évolutif des fleurs d'Angiospermeso En raison de cette absence de fossiles, les connaissances actuelles sur le dévelop-pement évolutif floral reposent presqu'entièrement sur les comparai-sons qui peuvent. etre tirées des études morphologiques~ anatomiques,
ontogéniques et histogéniques des types floraux en existence. Ces travaux ont fait l'objet de plusieurs revisions (exemples: Bancroft, 1935; Arber 1937, 1950; Poster et Gifford, 1959, Eames~ 1961; Bsau, 1965) et sont principalement reliés à la théorie florale dite "clas-siquell qui est influencée depuis ses origines par l'approche concep-tuelle idéaliste de Goethe (1790) tel que signalé par les historiens de la question florale. - Selon cette théorie, la fleur dans son ensemble est considérée comme un axe feuillé modifié: le réceptacle est une tige modifiée; les organes floraux sont des feuilles modifiées; les étamines et les carpelles sont des sporophylles (micro- et mégas-porophylles). Donc l'axe floral et ses appendices sont qualitative-ment et fondaqualitative-mentalequalitative-ment les homologues d'un axe végétatif et de ses
organes feuillés appendiculaires.
Cette théorie, selon Barnard (1961),
a
été modernisée
par
~es écoles de I.W. Bailey et ses associés, de AuJ. Eames et d'autreso19
Elle est également connuesous le nom de théorie ranaléenne du fait que selon ses promoteurs, les types floraux les plus primitifs sont trouvés dans les Magnoliaceae, les Ranunculaceae et les familles al-liées des ordres Magnoliales et Ranales chez les dicotylédones, et dans les Butomaceae et les falnilles alliées chez les monocotylédones. Tou-te spéculation sur la nature probable des proangiospermes envisage un type de gymnosperme du groupe des cycadophytes possédant des strobiles bisexués avec mégasporophylles en positIon distale, des microsporophyl-les en position sous-distale et, en position proximale, des feuilmicrosporophyl-les s'tériles qui seraient éventuellement devenues des pièces périanthaireso Les sépales et les pétales de la fleur primitive sont alors considé-rés des organes appendiculaires homologues des feuilles stérileso Les étamines et les carpelles primitifs sont des feuilles modifiées por-tant des sporanges, mais sans distinctions de filament et d'anthère, ou d'ovaire, de style et de stigrna o
Beiley et ses collaborateurs (Beiley et Smith, 1942; Bailey et Nast, 1943; Bailey et Smith, 1943) ont décrit les étamines primiti-ves de familles des Renales dont les Winteraceae et les
f;!ylnantandra-~. Ces étamines dans leurs formes les moins modifiées sont dorsi-ventrales, larges et foliacées, non différenciées en anthère, filament et connectif, et portent des sporanges enrobés par paires dans les tis-sus de leur surface abaxiale (ex. Degeneria). Selon Canr.ight (1952)
et Moseley (1958) les étamines sont moins foliacées chez les
Magnnlia-~ et les Nympheaceae respectivement. Les carpelles sensément les
plus primitifs sont rapportés par Beiley et Swamy (1951). Chez Dege-neria (Bailey et Smith, 1942) et Drimys piperita (Bailey et Nast 1943b),
tance des bords marginaux; les surfaces stigmatiques
poils glandulaires sur les bords libres marginaux. Le carpelle devient
éventuellement condupliqué avec bords marginaux coalescents, conservant au sommet une région stigmatique réceptrice du pollen. Dans
l'évolu-tion subséquente du carpelle, les ovules se forment sur les bords
mar-ginaux ép9issis en région placentaire. De ces caractères primitifs sont dérivés les gynécées composés,et d'autres formes de placentation, passant de la placentation laminaire à la placentation marginale, et de celle-ci aux placentations pariétales et axiles. (Pour une revue et
une discussiOn complète, voir Barnard, 1961; Eames 1961).
Sommairement, la fleur primitive ranaléenne possède un périanthe composé de plusieurs sépales et pétales libres, de nombreux étamines et carpelles libres, organes disposés en spirale sur un axe allongé. Cette théorie florale dans laquelle les catégories d'organes de la tige feuillée et de la tige florale sont "essentiellement
ll
homo-logues est à la base de divers systèmes de classification, ceux de
Bentham et Hooker (1862-67), de Bessey (1897, 1907), de Hallier
(1905-1912) et de Hutchinson (1926, 1929, 1969). Elle est également large-ment acceptée "à défaut" d'autres théories dégagées de
l'essentialis-me qui conduit au subjectivisl'essentialis-me au niveau des observations factuelles.
Cette théorie et son concept de l'homologie générale sont carrément rejetés par les uns et fortement mis en doute par d1autres. Ainsi, pour Grégoire (1938), le réceptacle floral n'est pas réductible
à un apex végétatif et le carpelle est irréductible à une feuille;
la fleur est donc "sui generisll • - Thompson nie l'existence du
l'ovule.
La fleur est avant tout un axe sporogène portant des parties florales qui ne sont pas des entités distinctes mais seulement des extensions locales du réceptacle. I l ne reconnatt donc pas d'entités appendiculaires de type foliaire. Troll (1939) retient le concept idéaliste de l'homologie générale, mais i l développe le "concept de type" sur la base duquel l'uniformité des plantes supérieures est vi-sualisée, c'est-à-dire "l'Urpflanze" de Goethe, dont l'existence com-me doctrine al~tonome de la forme est ici prouvée à nouveau et devient le centre méthodologique de toute étude spéciale. La morphologie est redéfinie sur une base typologique avec le résultat évident, selonAr-ber (1937), que l'approche typologique met de l'avant la méthode de la comparaison comme principe-guide. Cette forme de pensée, Arber la dé-finit .comme la morphologie du "Gestalt".
D'au·tres concepts sont avancés dans le but de réduire la di-versité des formes végétales à un nombre restreint de catégories d'ap-plication universelle. Ainsi, Zi~~ermann a proposé la théorie du
té-lome. D'après cette théorie, les plantes vasculaires les plus primiti-ves sont composées entièrement: de systèmes de télomes, c'est-à-dire d'un axe dichotomique dont les portions terminales ultimes peuvent ~tre
fertiles et porter des sporanges, ou bien être stériles et constituer des phyllofdes. Ces télomes qU'ils soient fertiles ou stériles peu-vent ~tre groupés en faisceaux'o Zimmerman et certains autres
morpho-logistes ont étendu l'applicatLon de ce concept au delà. du niveau des fougères pour. interpréter les organes foliaires et reproducteurs, même chez les angiospermes (ex. Wilson, 1937, dans l'interprétation des fascicules d'étamines chez certaines familles des Parietales et
des Malvales). Selon Mason (1957, po 93) "Si le complexe télome
-mésome est significatif dans les interprétations de la morphologie, i l
ne peut @tre utilisé dans aucun problème de comparaison diagnostique qui emploie la logique au-dessus du niveau de la différenciatio~ méris-tématique de l'organisation d'un système. Le télome est comme la
cel-lule une structure omniprésente. B::ant une caractéristique d'ordre
général, i l ne peut donc servir de diagnostique à des cas particuliers. Il (Traduct:l.0n littérale.) - En conséquence les morphologistes ne semblent guère justifiés d'étendre l'application de la théorie dtl té lome aux
structures des fleurs d'Angiospermes.
Cependant la théorie du télome fait époque en ce qU'elle
ins-pire d'autres concepts et théories selon lesq..lels les micro- et les
mégasporanges sont essentiellement des strU'::tures axiales depuis l'ori-gine, et non des structures d'origine foliaire (ou du moins pas néces-sairement). - Ainsi, Lam (1960) propose d,eux concepts pou.r tenir
comp-te de la diversité de position des sporanges dans le gynécée: la
sta-chyosporie et la phyllosporie. Si l'ovule naS:t sur une stru.::ture
fo-liacée (homologue d'une feuille ou phyllome) c'est la phyllosporie;
si au contraire l'ovtlle nô3.S:t Stlr un axe floral (homologue d'une tige
ou caulome), .c'est la stachyosporie. - Lam considère que la stc.lchyos-porie et la phyllosstc.lchyos-porie ont évolué indépendamment à part:i r d ' üu.-:ê l:r:cs hypothétiques gymnospermiques (mésocormophytes). Toutefois, ce SyS-tème mixte conduit à des difficultés parce qu'il ne peut tenir cümpte de toutes les situations plus ou moins intennédiaireso
Melville (1962, 1963) considère que l'unité de base de la
fleur est le gonophylle, c'est-à-dire une feuille avec branche
23
ne porte que des sporanges m~les, ou bien seulement des sporanges fe-melles, l'ensemble phyllome - branche fertile est désigné androphylle ou gynophylle respectivement o Dsns le cas où les sporanges m~les et femelles sont présents à la fois sur la branche fertile, llensemble phyllome - branche fertile se nomme androgynophyllea- Cependant lors-que la branche fertile est distinctement séparée de loël. structure
fo-liacée, cette structure est appelée tégophylle. - Cette théorie est considérée comme dépourvue de la base essentielle du concept et ne peut donc devenir un outil constructif po~r la synth~se et l'analyse
ce
de quoi que" soi t en botanique qui ne soit pas immédiatement d<9scrip-tif (voir Croizat, 1962, po 804, pour une analyse critique) 0
Finalement, pour Meeuse (1966), chez les Angiospermes les ovules naissent sur des axes ou leurs homologues. - Ainsi, Meeuse si-gnale (p. 127) que les ovules naissent essentiellemenot de cup,.lles, lesquelles dérivent de la fusion de télomes ou syntélomes protocauli-naires phyllodiques indépendamment des trophophylles; la cupule n'est donc pas une feuille. La structure ovulaire est un gynoclade o Les sporanges m§les naissent sur des rameaux fertiles appelés androclades. Les autres structures sont des énations stériles cauliro~ires~
I l ressort de 11 examen de ces théories que la perpétuelle controverse tourne a'.ltour des poinots suivants:
1- les anc@tres hypothétiques remontent à 11ère paléozo~que et sont postulés à partir des ptéridospermes (Glossopteris, pour Melvil-le) ou à partir des gymnospermes (Cycadopsida, pour la théorie classique, et pour Meeuse);
2- les organes floraux sont des structures transformées, homologw~s
(caulome) et la feuille (phyllome);
3- le concept de l'homologie relève d'un essentialisme idéologiqu~,
c'est-à-dire que, selon la théorie classique, les organes floraux sont 'essenti.ellement' ou 'fondamentalement' homolo~oo'es d'un phyl-lame ou d'un caulome;
4- le concept de l'homologie est d'ordre typologique et ramène les différentes structures à des modèles définis (Troll);
5- les sporanges sont homologues des feuilles selon la théorie clas-sique et selon celle de Melville (théorie des gonophylles), ou bien ils sont caulinaires, selon Meeuse;
6- la nature du réceptacle floral, et celle des organes constituant l'androcée et le gynécée sont les plus controverséeso
(Four des exposés détaillés et des discussions sur ces théo-ries, l'auteur réfère aux auteurs précités ainsi qu'à Eames, 1961; Barnard, 1961; Puri, 1951, 1952, 1961; et Croizat, 1962; eot autres).
Les étudiants modernes de la morphologie florale (ex.: MerxmUller 1963; Sattler, 1966) constaten~ le peu de p=ogrès accompli depuis plus de 150 ans, en termes d.= solutions finales. Ils recher_ chent les causes des différences profondes d' ino::erprétation et de re-connaissance des structures florales. Sattler (1966), dans sa recher-che d'une approrecher-che plus adéquate de la morphologie comparée, fait res-sortir l'acuité des difficultés reliées aux concepts de l'homologie générale et de l'homologie typolo3ique. Il démontre le danger d'abou-t i r à des raisonnemend'abou-ts circulaires par l'ud'abou-tilisad'abou-tion d .. ~ ces fO:!:1ues conceptuelles d'homologies. MacDonald (1971) rapporte que Forman
(1966) a cristallisé l'usage des homologies typologiques dans la dis-cussion de la cupule des Fagaceae: "Afin de comprendre la nature ou
2S
l'origine évolutive d'un organe, i l faut normalement l'interpréter
en termes d'un organe qui lui est homologue dans un groupe reliéQ
Ce-ci est la procédure normale pour essayer d'interpréter l'inconnu en
termes du connu" (traduit de l'anglais)u
Une telle approche basée sur des degrés de ressemblances
faciles à déte~niner, tels que le mode d'origine, la dimension, la
forme et la fonction~ a servi à établir des relations entre différents
taxons, sans'toutefois permettre de saisir la nature réelle du déve-loppement morphologiq'..le des organes comparés des divers taxons. I I s'ensui t donc que la valeur des degrés de ressemblances, mesurés en termes d'origine, de dimension, de forme et de fonction, n'est pas né-cessairement significative dans le procédé d'établissement des
homolo-gies. Il en résulte que les organes considérés homologues par un
chercheur ne le sont pas nécessairement par d'autres chercheurs (Sat$-1er, 1966; MacDonald, 1971).
Considérant donc d'une part, la faiblesse et les restrictions de la mécanique rationnelle de l'ho~ologie typologique et, d'autre part~ les illogismes a1JXquels conduit l'application de l'homologie générale
qualitative (philosophiq'.le, en termes d'essentialisme), Sattler (1966)
fait la critique des concepts et des théories en eKistence et propose
un concept de "l'homologie semi-quantitative". Ce concept, bien q"..1e plus ou moins philosophique, a pour but de placer la morphologie com-parée sur une base conceptuelle plus adéqu8te et de permettre de tirer,
à partir de comparaisons morphologiques, des déductions plus
significa-tives et moins biaisées phylogénétiquement. Sattler (1967) démontre
l'aspect semi-quantitatif de son concept en développant un modèle
organo-génétiques. Il illustre par ce modèle l'initiation pétalaire chez au moins 7 genres ou espèces offrant des difficultés d'interprétation .. Ce concept relève l'étudiant de la morphologie, ou florale ou générale, du développement de faire face aux questions suivantes: est-ce un
phyllome ou est-ce un caulome?, est-ce un primordium sépalaire, ou pé-talaire, ou staminal?, etc. Par ce concept, l'étudiant peut donc po-ser la question différemment: est-ce lldavantage" un phyllome ou un cau-lome?, etc., sans avoir à établir subjectivement l'homologie de tel ou tel organe comp9r~ et sans avoir à définir ces organes comme fondamen-talement ou essentiellement similaires entre eux. Les écueils des ho-mologies générales et typologiques sont donc ainsi largement prévenus. C'est par l'application intégrale de ce concept que MacDonald (1971) rejette en total~té les diverses théories conventionnelles et propo8e
un modèle pour le développement floral chez les "Amentiferae", ap-plicable à d'autres études comparées. C'est aussi sur la base du con-cept de IIl'homologie semi-quantitative" que l'organogénèse d'Ochna atropurpurea OC. sera considéréeo
2. Concepts histogéniques floraux.
Ce sous-titre est utilisé dans un sens restrictif et i l n'est applicable ici qu'aux méristèmes in.:lorescentiels et floraux. De fait, i l n'entre pas dans le cadre original de cette étude de con-sidérer les diverses étapes de transition depuis l'état méristématique des apex .caulinaires jusqu'à la formation des apex de l'inflorescence et des fleurs.. Les difficultés relatives à ce sujet seront éviden-tes lors de la description de l'inflorescence. (Pour une revue d{·-taillée et une mise au point longuement attendue sur le comportement
21
des apex méristématiques lors de la transition de l'état végétatif à
l'état inflorescentiel ou floral, l'auteur réfère à Gifford et Cor-son, 1971.)
Toutefois, un bref rappel sur la structure et le fonctionne-ment du méristème cau1inaire chez les angiospermes s'imposeo . En effet, un certain nombre de concepts ont été développés par différents bota-nistes concernant les apex végétatifs, depuis les travaux de Naegeli (1845) et ceux de Hofmeister (1851), et ces mêmes concepts furent éven-tuellement apliqués aux apex inflorescentiels et floraux. Ces deux auteurs établirent que le sommet des tiges de diverses ptéridophytes est occupé par une grande cellule pyramidale. Celle-ci se divise dans des plans parallèles à ses faces et donne ainsi naissance à tous les
tissus de la tige o C'est à Naegeli, selon Reed (1942) que l'on doit le terme méristème qu1il définit: "un groupe de cellules capables de se diviser". Plus tard, ces m@mes auteurs appliquent le concept de l'initiale unique aux phamérogames (Hofmeister, 1857; Naegeli, 1878) mais ce point de vue est combattu par Hanstein (1868) et Douliot
(1890). Hanstein propose à la place, la théorie des histogèneso Pour lui, i l existe dans le sommet des tiges trois cellules apicales su-perposées qui donnent naissance, en se divisant, à trois régions dis-tinctes et indépendantes: le dermatogène qui est à l'origine de l'épi-derme de la tige et des feuilles, le périblème gui fournit les tissus corticaux de la tige et (probablement?) les feuilles et,fin.:lle.'l\ent, le plérome qui produit les tissus vasculaires et la mo~lle de la tige.
A l'exception du terme "dermatogène", que l'on retrouve dans la litté-rature moderne selon son acception originale, ces concepts sont main-tenant abandonnés o
En 19211, :;chlll.i.dt prnpo:;e uIIe dn:lcri.ptillfl t:(lp()!Jr(lplt.i.ql((~ du méristème apical comme étant constitué de deux zones n~ttement d.i.st
inc-tes. Les cellules apicales les plus superficielles se divisent unique-ment dans le sens anticline de telle sorte q~'elles se disposent en une ou plusieurs assises régulières, caractéristiques de la "tunica"g Toutefois, lors de l'initiation d'une feuille, i l se produit des mi-toses périclines dans les assises tunicales, sauf dans l'assise la plus externe. Les cellules sous-jacentes à la "tunica" se divisent dans tous les sens: anticline, péricline, oblique; elles constituent une zone dont les cellules ne présentent pas de disposition particuliè-re. Cette zone, c'est le "corpus".
Le rcSle de ces deux zones est évident: la "tunica" assure la croissance en surface de l'apex, tandis que le· "corpus" réalise la croissance en volume. La "tunica" est à l'origine des tissus dermi-ques et le "corpusll fournit les tissus médullaires au centre de la tige ainsi que les tissus cortica~~ placés entre le derme et la région
médullaire centrale.
Bien que jugée partiellement ou entièrement insuffisante par les auteurs actuels et selon les écoles de pensées, la théorie de Schmidt n'a pas été complètement délaissée. Gifford (1954) dresse une liste d'angiospermes étudiées depuis 1924 dont les apex présentent la répartition de leurs cellules en une "tunicall et un IIcorpusll •
Cette répartition topographique des cellules apicales dans les apex végétatifs a été retracée dans le méristème inflorescentiel ou floral par plusieurs auteurs (ex.: Newman, 1936; Satina et Blakes-lee, 1941; ~eeve, 1943; Beke, 1947; Tepfer, 1953; Barnard, 1960; Ber-nier, 1964; et d1autres).
29
Cependant, la nature et la circonscription dl~ ces deux ~ones
au niveau floral (ou inflorescentiel) semblent avoir soulevé certaines difficultés. Ainsi, Grégoire (1938) reconna~t également deux zones. La première zone ne constitue qu1un manchon superficiel de quelques cellules d1épaisseur; mais son étendue est importa~te en ce qu1elle recouvre la totalité de la surface apicale. Les cellules de ce man-chon sont très méristématiques et se divisent principalement de façon anticline. Sous ce manchon, se trouvent des cellules largement va-cuolisées constituant la région médullaire du x'éceptacle floral (ou inflorescentiel); c1est le 2orte-méristème. Grégoire rejette toute idée d'homologie du manchon et du porte-méristème avec la "tunica" et le "corpus" de l'apex végétatif: "le réceptacle floral n'est pas ré-ductible à un apex végétatif "selon ses termes.
Plus tard, Lawalrée (1948) remarque que le "corpus" du mé-ristème reproducteur d'un capitule en développement est d1abord un massif lenticulaire biconvexe. I l constate que les divisions
anticli-nes y sont largement prédominantes et qu'éventuellement le "corpusll
devient étalé en nappe sous la "tunica". Pour cet auteur, le "corpus" ne correspond donc plus exactement au concept que Schmidt avait élaboré.
Selon Bernier (1964), malgré les réserves exprimées sur la nature des zones méristématiques florales,certains auteurs continuent l'emploi des termes "tunica" et IIcorpusli. De plu.:;, i l ajoute que la
con~eption de Grégoire (1938) est actuellement admise; elle se retrou-ve sous des formes diretrou-verses dans les travaux de B~vat (1952), Lance
(1958), Popham et Chan (1952), Wetmore, Gifford et Green (1959), Gif-ford et Tepper (1961). Il faudrait ajouter à cette lis.t;e~ Bernier
D'après les considérations de Lawalrée (1948) et celles de Bernier (1964), il faut s'attendre à ce que les méristèmes reproduc-teurs présentent une variété structurale beaucoup 'plus grande que les méristèmes végétatifs, en raison même de la diversité florale et
in-florescentielle en existence.
Cepen9ant, queHqUesoient les types morphologiques produits par les apex reproducteurs soit, une fleur terminale unique, une grappe définie, une ombelle ou autres, ils ont tous en commun la caractéristique d'une croissance limitée, par suite de l'utilisation complète de la surface apicale pour l'édification de fleurs ou de diverses pièces florales. - Une telle situation n'existe pas chez les apex végétatifs où l'initiation foliaire est confinée aux méristèmes organogéniques latéraux et où les activités méristématiques apicales peuvent se poursuivre 'indéfiniment.
Il se dégage de cet exposé des concepts histogéniques flo-raux, que malgré l'affirmation de Bernier (1964) sur l'admission de la conception de Grégoire (1938) d'après la liste d'auteurs qu'il soumet, i l reste encore des doutes. De fait, les étudiants de l'his-togénèse florale font encore face à la question philosophique du "OU bien, O'..l" telle qu'exposée précédemment concernant l'homologie géné-rale de la théorie classique, quant à la présence ou à l'absence des zones "tunica-corpus". En raison de la diversité des inflorescences, ces concepts méritent d'@tre reconsidérés de façon plus empirique.
Dans la poursuite des buts proposés dans l'introduction con-cernant l'étude du développement floral chez Ochna atropurpurea DG., les aspects suivants sont étudiés:
descrip-31
tion succinte de l'inflorescence;
b- la description de llorganogénèse florale depuis le stade de transition du méristème apical reproducteur jusqu1au stade juvé-nile des organes floraux à l'intérieur du rameau inflorescentiel; c- la description de Ilhistogénèse des stades équivalents à ceux de
l'organogénèse florale;
d- la description de la vascularisation florale limitée surtout aux stades juvéniles de llandrocée et du gynécée selon les évidences disponibles. Cette description sera étendue à llensemble de l'axe floral tel que révélé par les techniques d1éclaircissement.
Les observations factuelles tirées de cette étud~ conduiront
à une discussion des aspects traités, insistant surtout sur llandrocée et le gynécéee Le caractère du développement séquentiel de l'androcée
est particulièrement considéré en tant que critère diagnostique de base du postulat de Cronquist (1968) vis-à-vis les positions taxono-mique et phylogénétique de la famille des Ochnaceaeo