Examining how PSMB11 orchestrates T cell development
130
0
0
Texte intégral
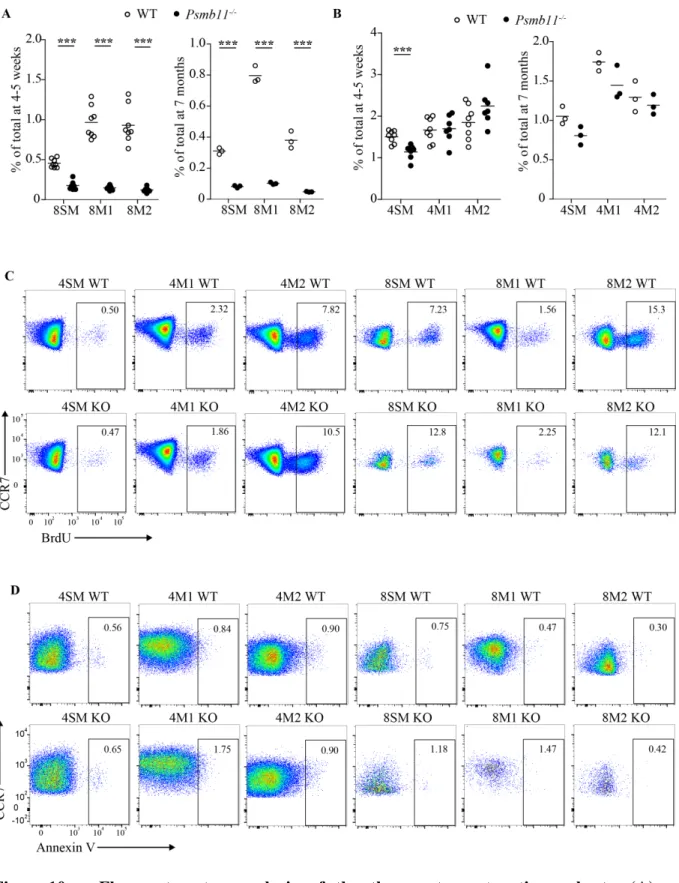
Figure
+7
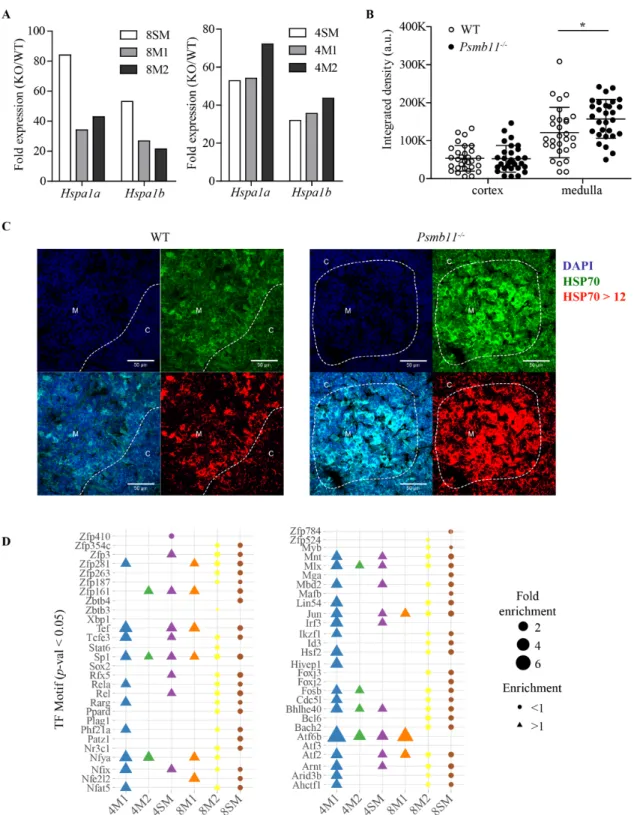
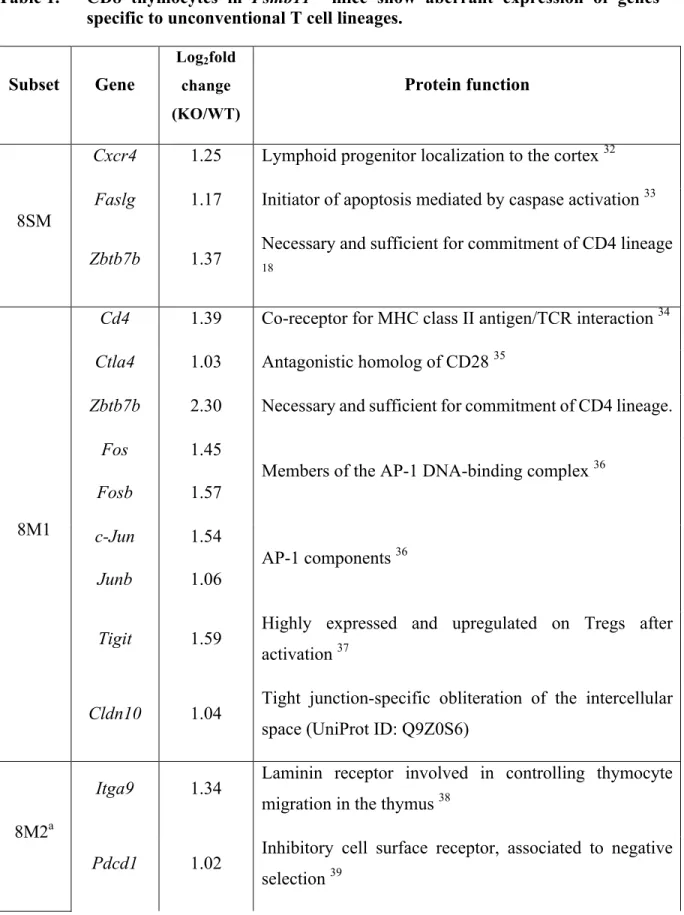

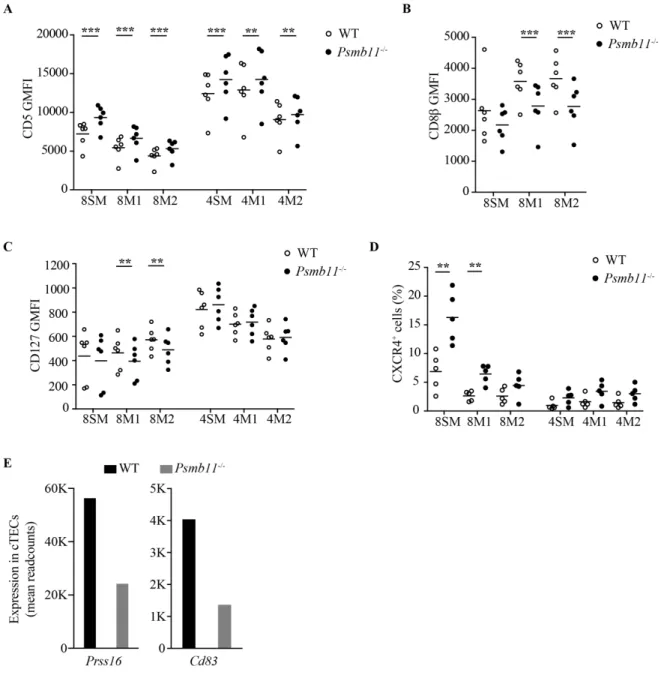
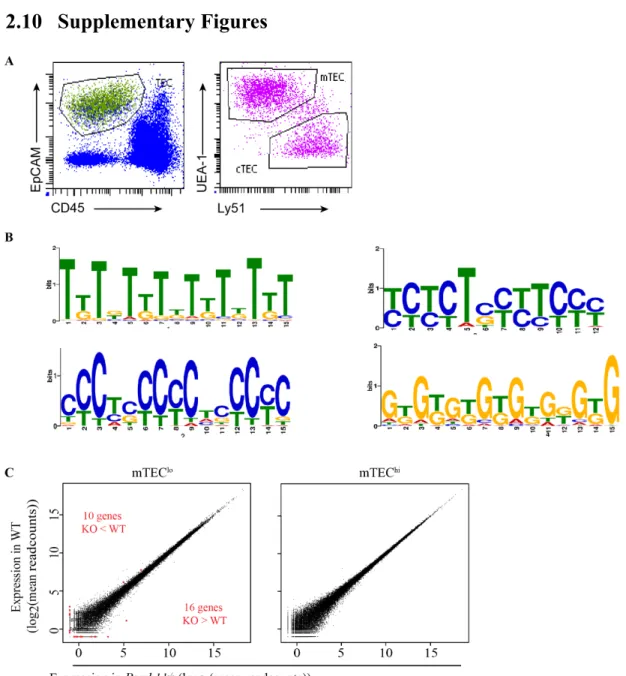
Documents relatifs
