L'interrelation entre l'empathie et la régulation
émotionnelle : corrélats neuronaux et autonomiques
Thèse
Josiane Jauniaux
Doctorat en psychologie - recherche et intervention
Philosophiæ doctor (Ph. D.)
L’interrelation entre l’empathie et la régulation
émotionnelle : corrélats neuronaux et autonomiques
Thèse
Josiane Jauniaux
Sous la direction de :
Pr. Philip Jackson, Ph.D., directeur de recherche
Résumé
L’empathie et la régulation émotionnelle sont des processus vitaux au fonctionnement socioémotionnel sain. Le terme « empathie » réfère à la capacité de partager et comprendre les émotions d’autrui. La régulation émotionnelle se définit quant à elle par la capacité de moduler ses propres états émotionnels. En neurosciences, bien qu’il ait été proposé d’un point de vue théorique que l’empathie et la régulation émotionnelle soient intimement liées, ces deux entités sont actuellement étudiés en vase clos. Par ailleurs, la littérature scientifique regorge d’études en neuro-imagerie fonctionnelle examinant les corrélats cérébraux de l’empathie pour la douleur. Or, l’empathie est une fonction sociale polyvalente se déployant dans une vaste étendue d’interactions socioémotionnelles. Tout comme la régulation émotionnelle, la valence émotionnelle demeure rarement examinée dans le contexte de l’empathie et l’interrelation entre la régulation et la valence émotionnelle est inexplorée. La thèse a pour objectif d’examiner la relation entre l’empathie, la régulation émotionnelle et la valence émotionnelle, de même que les corrélats neurophysiologiques sous-jacents. Un premier article de revue narrative décrit les régions cérébrales clés impliquées dans l’empathie pour la douleur, qui incluent notamment le cortex cingulaire antérieur médian et l’insula antérieure. Cet article a aussi permis de dégager de la littérature des facteurs régulant la réponse cérébrale associés à l’empathie pour la douleur. Ceux-ci incluent différents stimuli visuels, dont des membres du corps en douleur ou des expressions faciales de douleur. Ces derniers influencent les processus perceptuels de régulation. Les instructions offertes aux participants constituent un autre facteur pouvant influencer la réponse cérébrale, notamment leur orientation (vers soi ou vers autrui). Celles-ci influencent les processus cognitifs de régulation. Dans le deuxième article, une méta-analyse quantitative des études en neuroimagerie fonctionnelle sur l’observation de la douleur est présentée. Celle-ci révèle un réseau central d’activation associé à l’empathie pour la douleur (cortex cingulaire antérieur médian, insula antérieure). Ce réseau s’activerait indépendamment des processus perceptuels et cognitifs de régulation. On démontre également que différents processus perceptuels sont associés spécifiquement à des activations dans les régions sensorimotrices (observation de membres du corps en douleur) et à des activations dans une région impliquée dans l’imitation d’action et la communication non-verbale (observation d’expressions faciales de douleur). Différents processus cognitifs de régulation sont associés spécifiquement à l’activation d’une région impliquée dans l’intéroception et la conscience de soi (perspective orientée vers soi) ou à l’activation de régions impliquées dans le réseau attentionnel (perspective orientée vers le stimulus). Dans le but d’examiner expérimentalement l’effet de la régulation émotionnelle sur l’empathie, une étude psychophysiologique a été ensuite menée. Un nouveau paradigme expérimental a été développé. Lors du visionnement de vidéos illustrant des scènes d’interactions socioémotionnelles, des participants étaient invités à réguler leurs émotions par la réévaluation cognitive. L’empathie situationnelle, l’activité électrodermale et cardiaque ont été mesurées. Les résultats démontrent que la régulation émotionnelle module l’empathie situationnelle et qu’elle est associée à une augmentation de la variabilité du rythme cardiaque. Ces résultats suggèrent que la régulation émotionnelle est sous-tendue par l’implication du système parasympathique dans l’empathie. Aussi, l’observation d’émotions positives en comparaison à des émotions négatives est associée à moins d’empathie situationnelle en parallèle à une légère augmentation de la variabilité du rythme cardiaque. Ces résultats suggèrent une plus grande implication des processus de régulation lors de
l’empathie pour les émotions positives. En somme, cette étude démontre que les processus de régulation dans l’empathie seraient sous-tendus préférentiellement par le système parasympathique. Elle soulève par ailleurs que la valence émotionnelle est un paramètre important à considérer dans l’étude de l’empathie, puisqu’elle influence différemment les réponses subjectives et autonomiques associées. En plus d’offrir une vision plus intégrative de la relation entre l’empathie et la régulation émotionnelle elle soutient empiriquement leur lien. Les données de celle-ci démontrent que l’empathie est un phénomène dynamique pouvant être influencé par les processus perceptuels et cognitifs de régulation, lesquels influenceront différemment la réponse cérébrale et autonomique. En définitive, cette thèse contribuera à développer des modèles plus nuancés reliant l’empathie et la régulation émotionnelle ainsi que les bases neurophysiologiques qui y sont associées.
Abstract
Empathy and emotion regulation are vital processes for healthy socioemotional functioning. Empathy refers to the ability to share and understand others’ emotions while emotion regulation is defined as by the ability to modulate one owns’ emotional state. It is commonly described, from a theoretical perspective, that empathy and emotion regulation are intimately related. However, empathy and emotion regulation have been largely studied separately. In addition, the vast majority of the neuroscience literature on empathy is based on functional neuroimaging studies of vicarious pain. Empathy is although a versatile social function deployed in a large range of socio-emotional interactions. Like emotion regulation, emotional valence is rarely examined in the context of empathy and the relation between emotion regulation and emotional valence during empathy is currently unexplored. The objective of this thesis was to examine the relation between empathy, emotion regulation, and emotional valence, as well as their underlying neurophysiological correlates. In a narrative review article, the key brain regions involved in pain empathy are described, which includes the median anterior cingulate cortex and the anterior insula. In addition, factors that regulate the brain response during pain empathy were pointed out. These included different visual stimuli that regulate differently perceptual processes, such as body parts being submitted to noxious pain or facial expressions of pain. Instructions offered to the participants is also a factor that cognitively regulate the brain response during pain empathy, such as instructions oriented towards oneself or the other. In a second article, a quantitative meta-analysis on functional neuroimaging studies of pain empathy is presented. This study revealed a core network of activation related to pain empathy (median anterior cingulate cortex, anterior insula), which activates independently from perceptual and cognitive processes of regulation. This study also demonstrated that different perceptual processes distinctively activate sensorimotor regions (observation of limbs in painful situations) and a region involved in action imitation and non-verbal communication (facial expressions of pain). Furthermore, different cognitive regulatory processes distinctively activate a region involved in interoception and self-awareness (self-oriented perspective) and regions involved in the attentional network (perspective oriented towards the stimulus). In order to examine experimentally the effect of emotion regulation on empathy, a psycho-physiological study was then conducted. A new paradigm was developed. During the viewing of short videos depicting socioemotional interactions, participants were invited to regulate their emotions using cognitive reappraisal. Situational empathy was measured. Electrodermal and cardiac activity was gathered. Results showed that emotion regulation can increase or decrease situational empathy and is associated to an increase of the heart rate variability. These results suggest that emotion regulation is underpinned by the parasympathetic system during empathy. Moreover, viewing positive emotions, compared to negative emotions, was associated with less situational empathy and a slight increase of the heart rate variability. These results suggest a greater need in emotion regulation processes during empathy for positive emotions. In sum, this study demonstrates that emotion regulation process during empathy is preferentially underlied by the parasympathetic system. This work also highlights that emotional valence is an important parameter to consider when studying empathy, as it influences the underying subjective and autonomic responses. In addition to offer a more integrative vision of the relation between empathy and emotion regulation and supports their link empirically. The findings demonstrate that empathy is a dynamic phenomenon that can be regulated by
perceptual and cognitive processes. Ultimately, this thesis will contribute more nuanced models of empathy that will consider emotion regulation processes and the underlying neurophysiological basis.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... vi
Liste des figures ... ix
Liste des tableaux ... x
Liste des abréviations et des sigles ... xi
Remerciements ... xiii
Avant-propos ... xv
Introduction ... 1
L’empathie ... 1
Un aperçu historique de ses fondements conceptuels ... 1
Les neurosciences : une conceptualisation moderne ... 5
La composante affective de l’empathie ... 6
La composante cognitive de l’empathie ... 15
Le paradigme de l’observation de la douleur ... 19
Intégration des composantes et processus ... 21
La régulation émotionnelle ... 23
La régulation émotionnelle : une composante distincte dans l’empathie ? ... 23
Le modèle modal des émotions... 25
Le modèle du processus de la régulation émotionnelle ... 26
Les stratégies de régulation émotionnelle ... 27
L’empathie et la régulation émotionnelle : limites théoriques ... 31
La régulation émotionnelle : la laissée pour compte dans l’étude de l’empathie ... 31
La réévaluation cognitive : une stratégie efficace de régulation émotionnelle lors de l’empathie ? ... 34
L’empathie et la valence émotionnelle : limites empiriques ... 34
L’empathie pour les émotions positives ... 35
Les connaissances actuelles peuvent-elles se généraliser à tous les processus impliqués dans l’empathie ? ... 35
Objectifs de la thèse ... 37
Chapitre 1 : La réponse cérébrale à la douleur des autres ... 41
Résumé ... 42
Abstract ... 43
Introduction ... 44
La communication de la douleur ... 45
La représentation de la douleur de soi et des autres ... 45
L’impact des stimuli présentés ... 47
Les facteurs intrapersonnels, interpersonnels et situationnels ... 48
Conclusion ... 52
Remerciements ... 53
Références... 54
Chapitre 2: A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective ... 58
Résumé ... 59
Abstract ... 60
Introduction ... 61
Methods ... 65
Studies and coordinates selection. ... 65
Coordinate-based meta-analysis: ALE ... 72
Results ... 74
Pain empathy ... 74
Visual cues... 77
Visuospatial perspective ... 80
Self/other cognitive perspective taking ... 82
Discussion ... 88
Empathy and its related processes ... 89
An affective representation: at the core of pain empathy ... 90
A somatosensorimotor representations ... 92
Processing social nonverbal communication cues ... 93
Secondary networks implicated during pain empathy ... 94
Visual cues ... 94 Visuospatial perspectives ... 97 Cognitive perspective ... 98 Limitations ... 101 Conclusion ... 101 Acknowledgements... 102 References... 103
Chapitre 3: Emotion regulation of others’ positive and negative emotions is related to distinct patterns of heart rate variability and situational empathy ... 115
Résumé ... 116
Abstract ... 117
Introduction ... 118
Material and Methods ... 118
Participants ... 123
Procedure ... 123
Stimuli... 124
Experimental design ... 124
Positive affect and negative affect scale ... 126
Emotional valence and arousal scales ... 126
Situational empathy scale... 126
Dispositional empathy scale... 127
Autonomic measures ... 127
Heart rate variability analysis ... 128
Respiration rate ... 128
Electrodermal responses ... 129
Statistical analysis ... 130
Manipulation verification ... 131
Results ... 132
Effects of emotion regulation and valence on situational empathy ... 133
Effect of emotion regulation on autonomic responses ... 134
Effect of valence on autonomic responses ... 136
Interaction effect of emotion regulation and valence on autonomic responses ... 137
Discussion ... 137
Emotion regulation impacts situational empathy... 138
Emotion regulation increased cardiac autonbomic regulation during situational empathy .. 138
Emotion regulation of others’ positive vs. negative emotions is related to lesser situational empathy and greater cardiac autonomic regulation ... 142
Strengths and Limitations ... 143
Conclusion ... 145
Acknowledgements... 146
References... 147
Discussion générale ... 147
Résumé et intégration des résultats... 153
Premier article : La réponse cérébrale à la douleur des autres ... 153
Deuxième article : Méta-analyse quantitative sur les études en neuro-imagerie fonctionnelle de l’observation de la douleur : rôle de l’information visuelle et de la perspective de l’observateur. ... 154
Intégration des résultats du premier et deuxième article ... 156
Troisième article : La régulation des émotions lors de l’empathie pour les émotions positives et négatives est associée à des patrons distincts de régulation autonomique cardiaque et d’empathie situationnelle. ... 158
Intégration des résultats du deuxième et troisième article ... 159
Contributions de la thèse ... 165
Limites et perspectives futures ... 168
Questions futures en recherche fondamentale ... 170
Questions futures en recherche clinique et retombées potentielles ... 174
Conclusion générale... 177
Bibliographie ... 179
ANNEXE A: Sondage pour l’étude « A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective » ... 191
ANNEXE B : Descriptions des études incluses dans l’étude « A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective » ... 192
Annexe C: Analyses complémentaires pour l’étude «A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective ». 201 ANNEXE D : Schéma illustrant la tâche expérimentale pour l’étude « Emotion regulation of others’ positive and negative emotions is related to distinctive patterns of heart rate variability and situational empathy »... 209
ANNEXE E : Description des vidéos utilisées pour l’étude « Emotion regulation of others’ positive and negative emotions is related to distinctive patterns of heart rate variability and situational empathy » ... 210
ANNEXE F : Étude de validation du protocole de l’étude « Emotion regulation of others’ positive and negative emotions is related to distinctive patterns of heart rate variability and situational empathy » ... 211
Liste des figures
Chapitre 2: A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective
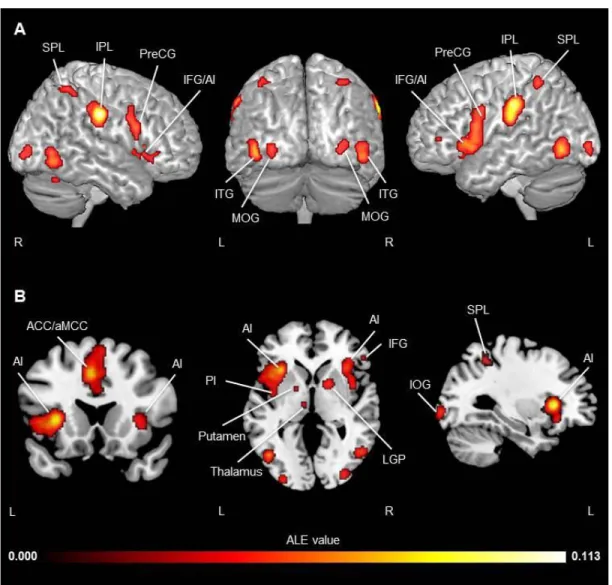
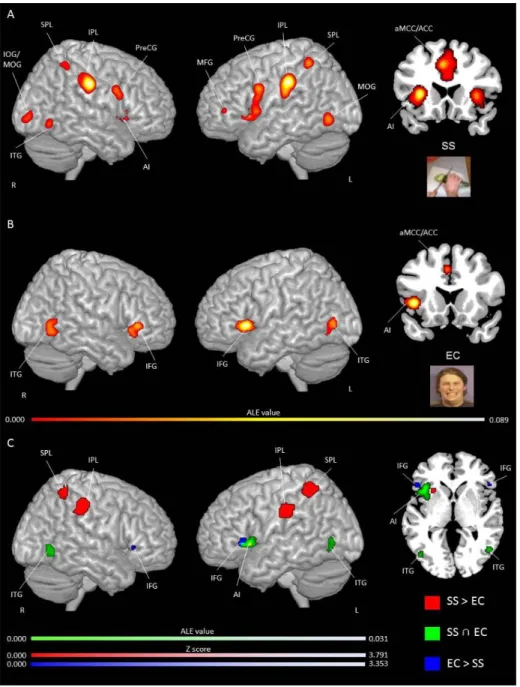
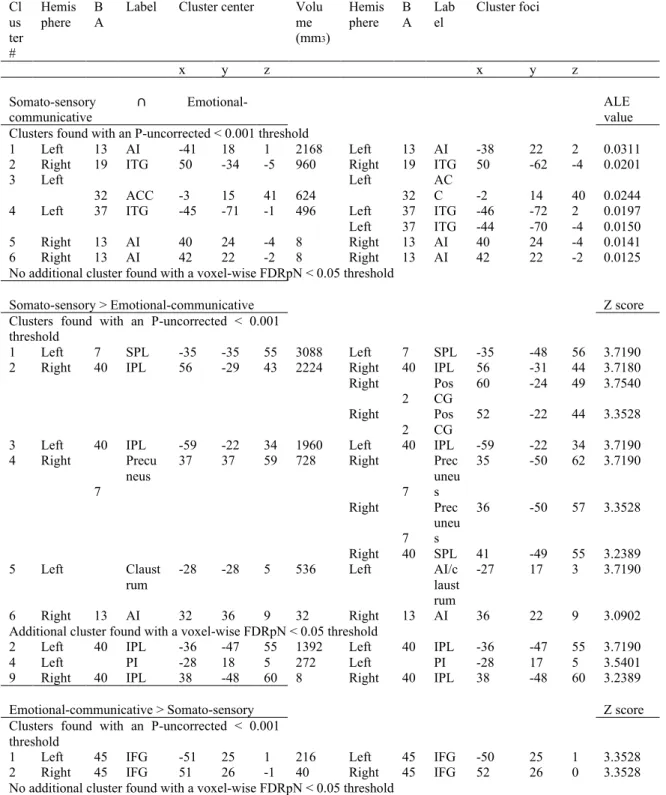
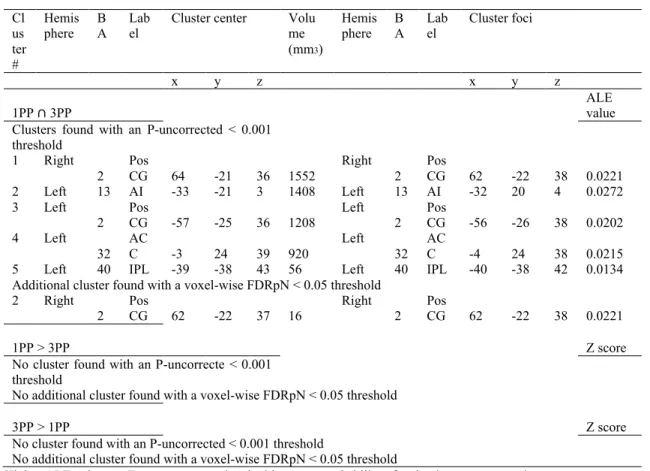
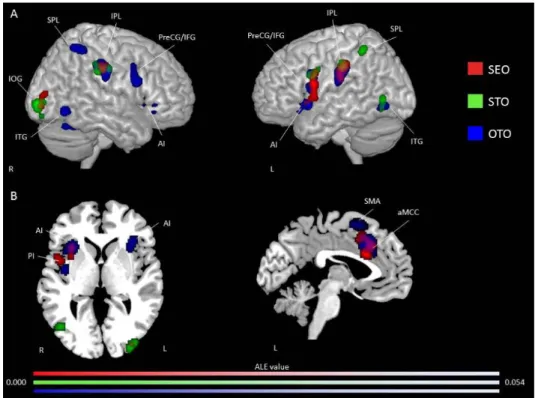
Figure 1. Study selection process………...………...……81 Figure 2. Activation likelihood clusters across all pain empathy experiments for the pain > no pain condition………...89 Figure 3. Activation likelihood clusters for pain empathy experiments using SS or EC visual pain information during a pain > no pain condition………..………...92 Figure 4. Activation likelihood clusters for pain empathy experiments using SS pain information presented from a 1PP or 3PP during a pain > no pain condition……..……...95 Figure 5. Activation likelihood clusters for pain empathy experiments using SS pain information and instructions to adopt either a self-perspective or another person’s perspective, or to focus on the stimuli during a pain > no pain condition………..…………..97 Figure 6. Common and distinct activation likelihood clusters for pain empathy experiments using SS pain information and instructions to adopt either a self-perspective or another person’s perspective, or to focus on the stimuli during the pain > no pain condition………....…98
Chapitre 3: Emotion regulation of others’ positive and negative emotions is related to distinctive patterns of cardiac autonomic regulation and situational empathy
Figure 1. Mean of participants’ ratings on the situational empathy scale during Emotion Regulation and Valence conditions ……….………...…134 Figure 2. Participants’ cardiac indexes on average during Emotion Regulation conditions………..…….………...…..135 Figure 3. Participants’ electrodermal responses on average during Emotion Regulation conditions………..……..………...….135 Figure 4. Participants’ cardiac indexes on average during Valence conditions ………..……..………....….136 Figure 5. Participants’ electrodermal responses on average during Emotion Regulation conditions……….….……..………...….137
Liste des tableaux
Chapitre 3: A meta-analysis of neuroimaging studies on pain empathy: investigating the role of visual information and observers’ perspective
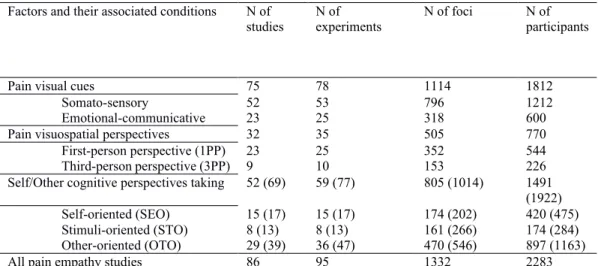
Table 1. List of references included in the meta-analysis and their corresponding factors and conditions………....……...……...…84 Table 2. Description of the sample: number of selected studies, experiments, foci and participants in each factor and their associated conditions………...….86 Table 3. Significant activation likelihood clusters across all pain empathy experiments………...……90 Table 4. Significant activation likelihood clusters for conjunctions and subtractions analyses for the pain visual cue factor…….………...…..93 Table 5. Significant activation likelihood clusters for conjunctions and subtractions analyses for the pain visuospatial perspective factor………...………...96 Table 6. Significant activation likelihood clusters for conjunctions and subtractions analyses for the self/other cognitive perspective taking factor…………...………..…99 Table 7. Significant activation likelihood clusters for conjunctions and subtractions analyses for the self/other cognitive perspective taking factor with studies using an SS or an EC paradigm………..………...….100
Chapitre 4: Emotion regulation of others’ positive and negative emotions is related to distinctive patterns of cardiac autonomic regulation and situational empathy
Liste des abréviations et des sigles
1PP First-person perspective3PP Third-person perspective ALE Activation likelihood estimation
ANS / SNA Autonomic nervous system / Système nerveux autonome BF / LF Basse-Fréquence / Low-Frequency
BOLD Blood-Oxygen-Level-Dependant
EBA Extrastriate body area / Aire extrastrié du corps EC Emotional-communicative pain paradigms
CCA / ACC Cortex cingulaire antérieur / Anterior cingulate cortex CPF Cortex préfrontal
dCCA / dACC Cortex cingulaire antérieur dorsal / Dorsal anterior cingulate cortex FBA Fusiform body area / Aire du corps fusiforme
FDRpNs False discovery rates voxel-wise
FFA Fusiform face area / Aire du visage fusiforme GEE Generalized Estimating Equations
GFI / IFG Gyrus frontal inférieur / Inferior frontal gyrus GFM / MFG Gyrus frontal médian / Medial frontal gyrus GFS / SFG Gyrus frontal supérieur / Superior frontal gyrus GMT / MTG Gyrus médian temporal / Middle temporal gyrus GOI / IOG Gyrus occipital inférieur / Inferior occipital gyrus GOM / MOG Gyrus occipital médian / Middle occipital gyrus GTI / ITG Gyrus temporal inférieur / Inferior temporal gyrus GTS / STG Gyrus temporal supérieur / Superior temporal gyrus
HF High-Frequency
IASP International Association for the Study of Pain IA / AI Insula antérieur / Anterior insula
IRM Imagerie par résonance magnétique
IRMf / fMRI Imagerie par résonance magnétique fonctionnelle / Fonctional magnetic resonance imaging
LF Low-Frequence
LF/HF-ratio Low-/High-Freqyency ratio MNI Montreal Neurological Institute
OFA Occipital face area / Aire du visage occipitale OTO Other-oriented tasks
PANAS Positive Affect and Negative Affect
PGL / LGP Pallidus globulis latéral / Lateral globus pallidus PreCG Gyrus pré-central / Precentral gyrus
PosCG Gyrus post-central / Post-central gyrus RRI Time intervals between each R peak SI Cortex somatosensoriel primaire
SII Cortex somatosensoriel secondaire / Secondary somatosensory cortices SAMS Self-Assessment Manikin Scale
SCRs Skin conductance responses SEO Self-oriented tasks
SMA Supplementary motor area
SS Somato-sensory noxious stimulation paradigms STO Stimuli-oriented tasks
Avec tout mon amour, je dédie cette thèse à ma fille. Quelle chance ai-je d'être ta maman. En espérant pouvoir partager avec toi les émotions qui coloreront ta vie. Sache que tu trouveras au cœur de mon amour l’empathie pour que tu puisses continuellement évoluer.
Remerciements
Cette thèse marque un tournant important dans ma vie. Nombreuses sont les personnes qui m’ont soutenue au cours de mes études. Sans elles, je n’aurais pu être la personne que je suis aujourd’hui et que j’aspirais devenir. Du plus profond de mon cœur, je vous remercie tous chaleureusement.
Je tiens à remercier chaleureusement mon directeur de thèse, Pr Philip Jackson, Ph. D. sans qui ces travaux de recherche n’auraient jamais pu être possibles. Philip, merci d’avoir cru en moi et surtout, de m’avoir soutenue tout au long de ces années, et ce, au-delà de mon parcours en recherche. Malgré mes moments d’incertitude et des projets qui m’ont fait emprunter des détours, j’ai toujours senti ta bienveillance et ta patience. J’ai un immense respect pour ta détermination, ta minutie et le sens critique tant caractéristiques de tes compétences de chercheur. Par ton sens de l’équité et de l’effort au travail, tu sais créer un contexte propice au développement de solides compétences professionnelles en recherche. Merci également pour toutes les possibilités que tu m’as offertes. Un immense merci pour tout !
J’aimerais remercier Pr Pierre Rainville, Ph. D., mon premier mentor qui était aux balbutiements de mon parcours en recherche. Je me souviendrai toujours de quand je t’ai présenté mes premiers résultats de recherche, un simple test-t. Par ta passion pour la recherche et ton approche pédagogique, tu as su éveiller à ce moment cette étincelle qui anime toujours à ce jour ma curiosité intellectuelle et mon intérêt pour la recherche. Merci d’avoir suivi et soutenu l’évolution de mes travaux au cours de ces années, ainsi que pour ta rigueur exceptionnelle et ta générosité. Tu as contribué à façonner la personne que je suis aujourd’hui. Mille mercis.
J’aimerais remercier Pr Carol Hudon, Ph. D. qui a également suivi l’évolution de mes travaux de recherche doctorale. Merci pour vos précieux commentaires lors de mes séminaires. Ceux-ci m’ont permis d’évoluer et ont contribué à développer mon intellect.
Merci à tous les membres du laboratoire qui ont contribué de près ou de loin à mes travaux de recherche, ainsi que pour tous les moments partagés au fil des années. Un merci tout particulier à Sophie Regueiro et à Marie-Hélène Tessier. Merci pour votre précieuse aide et votre amitié. Un merci tout spécial à Sarah-Maude Deschênes qui a su accueillir avec la plus grande zénitude mes angoisses. Merci pour les rires et les anecdotes de mamans partagées. J’aimerais remercier l’ensemble de la population québécoise et canadienne qui a subventionné ces travaux de recherche, sans quoi je n’aurai pu m’y consacrer pleinement. Merci aux Instituts de recherche en santé du Canada, aux Fonds de recherche du Québec – Santé et au Conseil de recherches en sciences naturelles et en génie du Canada.
Un merci tout spécial à mes amies. Merci Élisabeth pour ton amitié depuis le Cégep et qui s’est renforcie depuis que tu habites à Québec. Merci Marie-Ève, ma sœur de cœur, avec qui je passe les étapes les plus importantes de ma vie depuis que nous sommes aux couches. Merci, Catherine J., pour ta soif de l’aventure. Toujours partante pour des périples sans bon sens, j’adore être ta partner intrépide. Merci Pénélope, grande voyageuse, pour toutes les surprises toujours plus inusitées les unes que les autres et pour ton amitié infaillible. Un merci
tout spécial à Jessica L. avec qui j’ai passé à travers le baccalauréat. Merci d’avoir été là pendant ces soirées d’études interminables qui n’en finissaient plus de finir.
Merci Luce et Robert ! Merci de m’avoir accueilli dans votre famille les bras grands ouverts. Merci pour ces nombreux soupers, attentions et plats préparés. Merci d’être des grands-parents toujours prêts à offrir leur amour et leur temps. Votre appui est inestimable.
Merci à mes indestructibles frères, Emmanuel et David. Grandir entourée de frères a certainement forgé mon caractère. Merci d’être toujours les premiers partants pour une pêche sur la glace à moins 30 degrés l’hiver. Merci également à Valérie, ma belle-sœur. Ton sourire et ta générosité font chaud au cœur.
Mes chers parents. La fin de mes études représente en quelque sorte les fruits de vos efforts. J’aimerais vous remercier chaleureusement, puisque vous avez toujours su m’offrir tout ce dont j’avais besoin pour m’épanouir, ainsi que de solides valeurs sur lesquelles je peux m’appuyer pour évoluer. Sans vous, rien n’aurait été possible. Maman, tu m’as comblée de tendresse et d’affection tout au long de mon parcours. Tu m'as toujours soutenue et encouragée durant les moments difficiles et félicitée dans les moments de succès. La fougue qui m’anime continuellement, je te la dois. Papa, tu as su m’enseigner le sens de l’effort, de la responsabilité, de la débrouillardise, l’optimisme et la confiance en soi. La force morale dont tu disposes est inébranlable. Tu as contribué à la personne que je suis aujourd’hui et j’en suis fière. Je suis reconnaissante pour votre amour inconditionnel. J’espère pouvoir offrir à mon tour autant que j’aie reçu. Je vous l’assure, la barre est haute !
À mon amour, Mathieu. Je n’aurais jamais passé à travers mon doctorat sans ton amour inconditionnel et ton soutien. Tu es le gouvernail de notre bateau. Tu sais maintenir la force de mes voiles dans la bonne direction et naviguer avec moi malgré les aléas. Aucun mot n’égale l’immensité de ma gratitude et je souhaite de tout cœur que nos chemins puissent se poursuivre côte à côte encore des années. Que celles-ci soient remplies d’amour, de bonheur et des plus grandes aventures que la vie puisse nous offrir. Merci d’être toi, merci d’être là. Je t’aime.
À ma fille, Aurélie. Je suis sans mot lorsque je pense à l’amour que j’éprouve pour toi. Je donnerais pour toi à mille reprises ma vie. Ton sourire illumine nos vies. Merci pour les dessins et les bisous. Ils me donnent le courage de continuer. Je suis si heureuse d’être ta maman et de te voir grandir. Je t’aime de tout mon cœur.
Avant-propos
Le présent ouvrage constitue une thèse avec insertions d’articles, laquelle se divise en trois chapitres. Une introduction générale est d’abord présentée. Le premier chapitre présente un article de revue narrative qui a été publié dans la revue Douleur et Analgésie en mars 2014. Les chapitres deux et trois présentent des articles empiriques, lesquels découlent de travaux réalisés au cours du doctorat. L’article présenté au deuxième chapitre a été publié dans la revue Social Cognitive and Affective Neuroscience le 8 août 2019. Les résultats des analyses complémentaires à cet article sont présentés en annexe (C). L’article présenté au troisième chapitre a été soumis à la revue PLOS One le 18 septembre 2019 et il est, à l'heure actuelle, en révision. À la suite de modifications majeures, celui-ci a été soumis à nouveau le 21 mars 2020. Une étude pilote en amont à cette étude a été menée et est présentée en annexe (F). Outre la mise en page, les articles insérés dans la thèse sont identiques aux articles publiés ou en processus de révision. La thèse se termine par une discussion générale qui permet de faire l’intégration des résultats présentés aux trois chapitres.
Les trois articles présentés aux chapitres un, deux et trois ont été rédigés sous la supervision du directeur de la thèse, Pr Philip Jackson, Ph. D. Je suis la première auteure sur ces articles. J’ai contribué à chacune des étapes de la réalisation de chaque article. Pour l’article présenté au premier chapitre, j’ai recensé la littérature, déterminé les objectifs et rédigé le manuscrit en collaboration avec Sarah-Maude Deschênes, Dt.P., M.Sc. Pour l’article présenté au deuxième chapitre, j’ai effectué la recension systématique de la littérature, déterminé les objectifs, développé le devis de recherche, assuré la collecte et l’analyse des données, ainsi que rédigé le manuscrit. Pr Pierre Rainville Ph. D. a eu un apport considérable pour cette étude entre autres pour l’identification des objectifs de l’étude, l’élaboration du devis expérimental et la révision du manuscrit. Pr Ali Khatibi Ph. D. a également eu un apport important au niveau de la vérification de la recension de la littérature, l’identification des objectifs de l’étude, la vérification des analyses et la révision du manuscrit. Pour l’article présenté au troisième chapitre, j’ai effectué la recension des écrits, élaboré le devis expérimental, obtenu l’approbation éthique, effectué la collecte, le traitement, l’analyse et l’interprétation des données de même que la rédaction du manuscrit. J’aimerais souligner l’apport de chacun des coauteurs pour cette étude, particulièrement Sophie Regueiro pour son soutien dans la recension des écrits, l’élaboration du devis expérimental et l’acquisition des données, ainsi que Marie-Hélène Tessier pour son soutien au plan statistique et dans la révision du manuscrit. Je remercie Dr Florian Chouchou, Ph. D. pour son soutien dans le traitement et l’analyse des données de même que Dr Alexis Fortin-Côté Ph. D. pour son soutien technique dans le traitement des données. Je remercie Dr Jean Leblond, Ph. D. pour son soutien pour les analyses statistiques. Finalement, je souhaite remercier Dr Stephanie Sutliff, Ph. D. pour son soutien dans l’édition des articles présentés aux deuxième et troisième chapitres.
Introduction
L’introduction exposera d’abord un bref historique des fondements conceptuels de l’empathie. Suivra la présentation de modèles contemporains en neurosciences et certaines limites théoriques seront présentées. Les connaissances actuelles sur les mécanismes neurophysiologiques ainsi que les limites des études expérimentales actuelles seront par la suite présentées. Suivront les objectifs de la thèse.
L’empathie
Un aperçu historique de ses fondements conceptuels
Si le cogito ergo sum (« je pense donc je suis ») de Descartes en est la pierre angulaire, la philosophie moderne ne peut échapper à la deuxième maxime de la théorie kantienne, l’intersubjectivité, « penser du point de vue de n’importe qui d’autre » (Kant) (Jorland et Thirioux, 2008). Selon Kant, il importe d’aller au-delà de la pensée individuelle afin de l’élargir à un point de vue universel, ce à quoi on parvient « en se mettant à la place des autres » (Jorland et Thirioux, 2008). L’étude de l’empathie, ou de l’intersubjectivité, a été longtemps la muse intellectuelle de nombreux philosophes modernes sans qu’aucun terme spécifique soit utilisé (Jorlan et Thirioux, 2008). Einfühlung, terme germanique signifiant ressentir (fühlung) de l’intérieur (ein -), a été introduit pour la première fois par le philosophe allemand Robert Vischer (1847-1933) en 1872 dans sa thèse doctorale « Über
das Optische Formgefühl : Ein Beitrag zur Aesthetik » (« Sur le sentiment de l’optique de
forme : une contribution à l’esthétique »). Celui-ci s’était inspiré des écrits de son père, Friedrich Theodor Vischer (1807-1887), historien, philosophe et théoricien de l’art. Bien que le concept d’empathie puise ses plus anciennes racines idéologiques dans la pensée kantienne, c’est Robert Vischer qui a traduit l’idée en un terme. Celui-ci proposa un champ lexical original et varié qui décrit le sentiment éprouvé au contact des êtres humains. Seul le terme Einfühlung sera retenu par les philosophes subséquents. Einfühlung, qui signifie
into-feeling, a été traduit de l’anglais pour la première fois par Edward Bradford Titchener
(1909) par le terme empathiem soit la faculté de se mettre à la place d’autrui, de percevoir ce qu’il ressent (Larousse, 2003).
Theodore Lipps (1851-1914), un contemporain de Vischer, est la figure principale associée à l’origine du terme Einfühlung. Bien que Lipps ne soit pas le premier à l’utiliser, nombreuses sont les personnes qui le conçoivent comme le fondateur du concept d’empathie comme il a rédigé de nombreux écrits sur sa théorie de l’empathie esthétique. Il soutient dans sa thèse de doctorat « Einfühlung und asthetischer Genuss » (« Empathie et Plaisir Esthétique ») que lors de la perception d’un objet, quel que soit l’objet, le sujet s’empreint de l’affect et de la force dudit objet (Koss, 2006). Le terme Einfühlung s’est rapidement popularisé dans le domaine de la philosophie de l’art. Lipps ne sera pas le seul à parler de l’Einfühlung. Plusieurs de ses contemporains ont proposé des visions similaires, mais aussi alternatives. Les philosophes de l’époque, notamment Karl Groos, Vernon Lee et Jan Rybicki, suggèrent l’existence d’un processus de mimétisme moteur, c’est-à-dire que l’on imite intérieurement les caractéristiques d’un objet externe (Gladstein, 1984). Parallèlement, d’autres, notamment Max Scheler (1874-1928) et Edith Stein (1891-1942), se sont intéressés à l’empathie (ou sympathie à l’époque) comme élément clé aux comportements altruistes. Après l’élaboration et la popularisation des premières théories sur l’Einfühlung à la fin du 19e siècle et au début du 20e siècle, celles-ci ont été largement
critiquées par plusieurs cognitivistes en raison du manque d’objectivité et d’appui empirique (Nowak, 2011). L’étude de l’empathie deviendra alors rapidement obsolète.
L’étude de l’empathie connait un renouveau à la moitié du 20e siècle dans plusieurs
disciplines de la psychologie, notamment à la suite des tragiques événements entourant l’holocauste lors de la Seconde Guerre mondiale (Nowak, 2011). Ce nouveau pan de la littérature marquera la période contemporaine de l’étude de l’empathie (Gladstein, 1984). En effet, de nombreuses théories se seront développées dans le domaine de la psychologie sociale, développementale, cognitive, psychanalytique et humaniste. Notamment, le sociologue Charles H. Cooley (1864-1929) écrit dans son livre « Sympathy or Communion
as an Aspect of Society » que la sympathie, qu’il définit comme le partage de tout état
mental pouvant être communiqué, est cruciale à une société fonctionnelle. Le sociologue George H. Mead (1863-1931) a également contribué à la vision actuelle de l’empathie en introduisant la notion de role-taking, c’est-à-dire qu’afin d’être empathique, il est primordial d’être en mesure de se mettre à la place d’autrui. Le concept de prise de perspective connu des théoriciens actuels puise ainsi ses origines à cette époque. Pour
plusieurs auteurs, l’influence des écrits de Mead aura été cruciale pour les théories subséquentes (Gladstein, 1984). En effet, celles-ci définiront l’empathie comme un processus conscient et actif, plutôt que réactif et projectif, telle que le définissaient les théories de l’esthétique (Gladstein, 1984).
Les théoriciens de la psychologie développementale, tels que Jean Piaget (1896-1980) et James M. Baldwin (1861-1934), ont également contribué à l’étude de l’empathie (Gladstein, 1984). Ceux-ci suggéraient que le développement de l’enfant se caractérise entre autres par le surpassement de l’égocentrisme permettant de voir le monde à travers le point de vue des autres. Selon la vision piagétienne, les premiers signes de la sympathie apparaissent à l’enfance par les réactions affectives de l’enfant lorsqu’il est exposé à la détresse d’autrui et par les changements associés du système nerveux autonome sympathique (Gladstein, 1984). Similairement aux idées de Piaget, Baldwin définissait deux types de sympathie ; une sympathie organique, laquelle est instinctive et immédiate, et une sympathie réflective, laquelle émane en parallèle à la pensée (Gladstein, 1984). Baldwin suggérait également que lors d’une trop grande activation sympathique, une personne peut confondre sa propre souffrance avec celle de l’autre et croire que cette souffrance est sienne. Dès lors, on peut constater que le concept de distinction entre soi et autrui puise ses origines dans les théories de la psychologie développementale (Gladstein, 1984). Elle dépeint l’empathie comme un phénomène à la fois affectif et cognitif qui émane après le stade de l’égocentrisme (Gladstein, 1984).
Parallèlement à celles en psychologie sociale et développementale, plusieurs théories sur l’empathie ont été proposées dans le domaine de la psychologie clinique. Deux racines théoriques peuvent être identifiées et associées aux courants psychanalytique et humaniste issues respectivement de la pensée freudienne et rogérienne (Gladstein, 1984). Sigmund Freud (1856-1939) et Carl Rogers (1902-1987) s’appuieront sur l’observation des comportements des individus et des thérapeutes afin d’élaborer leurs idées. Freud discutera de l’empathie plus indirectement par le concept de l’identification émotionnelle, la première expression du lien émotionnel avec une personne. Il suggérait que l’ego (le soi) s’emprunte des caractéristiques de l’objet, c’est-à-dire la personne qui exprime des émotions, et devient un moyen d’introjection dans l’ego. L’introjection de l’objet dans
l’ego est en quelque sorte une perte de soi dans l’autre (Gladstein, 1984). Freud suggérait que le processus de l’identification expliquait en partie les liens étroits entre les membres d’un groupe et son leader (Gladstein, 1984). Carl Rogers, pour sa part, avait une vision plus cognitive de l’empathie (Gladstein, 1984). Il suggérait que la capacité à prendre la perspective d’une autre personne était cruciale pour un thérapeute. En effet, il accorde une attention particulière à l’attitude empathique. Il nommera cette dernière la compréhension empathique, c’est-à-dire la capacité d’un thérapeute à voir le monde comme le client le perçoit et pouvoir, en retour, exprimer cette perception à son client (Rogers, 1946). Il semblerait ainsi qu’au sein des théories les plus anciennes sur l’empathie en psychologie clinique, une distinction entre l’aspect affectif et cognitif de l’empathie se dessinait déjà.
De la fin du 20e siècle au début du 21e siècle, l’étude de l’empathie connaît une
troisième vague d’intérêt toujours grandissante encore à l’heure actuelle, et ce, dans plusieurs disciplines. Une simple recherche sur PubMed permet de constater l’intérêt quasi exponentiel pour le concept d’empathie. En 1985, 136 articles au total en lien avec ce concept avaient été publiés. En 2018, 1 683 articles ont été publiés. D’un côté, plusieurs philosophes et psychologues s’intéressent à l’empathie dans le contexte de la moralité. De l’autre côté, plusieurs chercheurs en cognition sociale s’intéressent à l’empathie dans le contexte des relations interpersonnelles, plus spécifiquement comme un agent de la compréhension interindividuelle (Goldman, 1995 ; Decety et Ickes, 2009). Il est possible d’identifier deux cadres théoriques distincts issus de la pensée cognitiviste du début du siècle actuel. Le premier englobe les théories entourant la théorie de l’esprit (theory of
mind). Le deuxième regroupe les théories de la simulation de la théorie de l’esprit
(simulation theory of mind) (Zahavi et Overgaard, 2012). Les auteurs de la théorie de l’esprit suggèrent que l’attribution des états mentaux à autrui s’effectue sur la base d’un processus de théorie de l’esprit. Ce dernier se développerait au cours de la petite enfance et se raffinerait tout au long de la vie (Gopnik, 2012). Pour les théoriciens de la simulation, l’empathie est une propriété de l’esprit humain qui utilise son propre esprit comme référence afin de comprendre les états mentaux d’autrui. Elle est une forme d’imitation interne des actions, des émotions et des sensations d’autrui. Certains diront que ce processus est conscient et délibéré (Goldman, 1995). D’autres diront que ce processus est inconscient et implicite (Gallese, 2009). Les théories de la simulation sont proposées
comme étant les plus proches cousines théoriques des théories actuelles en neurosciences sur l’empathie (Stueber, 2006).
Les neurosciences : une conceptualisation moderne
Le cerveau, par ses fonctions fines et complexes, confère à l’humain la capacité de lire les états mentaux d’autrui. L’arrivée des neurosciences, et particulièrement de l’imagerie par résonance magnétique (IRM) fonctionnelle (IRMf) cérébrale, a permis à l’étude de l’empathie d’atteindre un tout autre niveau. L’IRMf est une technique d’imagerie permettant de mesurer in vivo l’activité de régions cérébrales par la détection de changements locaux relatifs au flux sanguin cérébral (Andreelli et Mosbah, 2014). Afin de répondre aux besoins métaboliques liés à l’activation neuronale d’une région donnée lorsque celle-ci est sollicitée, le flux sanguin avoisinant augmente. Cette augmentation modifie le rapport entre l’oxyhémoglobine et la désoxyhémoglobine localement, ce qui occasionne un signal magnétique mesurable, le signal BOLD
(« Blood-Oxygen-Level-Dependant »), par l’appareil IRM (ou « scanner »). Lors de tâches expérimentales, dont
l’objectif est de solliciter certaines fonctions du cerveau, il est ainsi possible de mesurer indirectement l’activité cérébrale en captant le signal BOLD. Par le contraste entre le signal BOLD associé à une condition expérimentale et le signal BOLD associé à une condition contrôle, il est possible de documenter l’activation cérébrale associée spécifiquement à la condition expérimentale et aux fonctions qui y sont associées. Notons que l’IRMf ne permet pas d’inférer un lien direct entre une tâche et une région cérébrale, puisqu’elle mesure l’activité cérébrale de manière indirecte (Andreelli et Mosbah, 2014). Nonobstant, l’IRMf permet d’analyser l’architecture fonctionnelle du cerveau in vivo et procure un aperçu des corrélats d’activation neuronale relatifs à ses fonctions, notamment sociocognitives, telles que l’empathie.
Depuis les toutes premières études en IRMf sur l’empathie (Morrison, Lloyd et Roberts, 2004 ; Singer, Seymour, O’Doherty, Kaube, Dolan, et Frith 2004 ; Jackson et coll., 2005 ; 2006), de nombreuses définitions ont été proposées afin de décrire le phénomène. En fait, Cuff, Brown, Taylor et Howat (2016) en décomptent quarante-trois. Bien que l’empathie soit empreinte d’ambigüité conceptuelle (Zahavi et Overgaard, 2012),
il est généralement admis qu’elle se définit par deux composantes fondamentales, l’une affective et l’autre cognitive (Cuff et coll., 2016). Certains chercheurs accorderont également une importance particulière aux processus de régulation émotionnelle dans leur conceptualisation de l’empathie (Decety et Jackson, 2004 ; Decety et Lamm, 2006 ; Eisenberg et Eggum, 2009 ; de Waal et Preston, 2017). Cette composante demeure toutefois peu élaborée conceptuellement et théoriquement, et encore moins étudiée expérimentalement. Très récemment, des théoriciens du domaine des émotions ont proposé une vision intégrative liant la régulation émotionnelle à l’empathie (Thompson, Uusberg et Gross, 2019). Cette vision est pertinente aux objectifs de cette thèse et sera discutée plus en détails au cours de la présente introduction.
La composante affective de l’empathie
La composante affective de l’empathie, aussi connue par les termes partage
émotionnel (« emotion sharing ») (Decety et Meyer, 2008 ; Decety et Lamm, 2006), partage affectif (« affective sharing ») (Tousignant et coll. 2017), contagion émotionnelle
(« emotional contagion ») (de Waal et Preston, 2017), empathie émotionnelle (« emotional
empathy ») (Shamay-Tsoory, 2011) ou encore expérience partagée (« experience sharing ») (Zaki et Ochsner, 2012), se définit comme « une réponse affective à une autre
personne qui, souvent, mais pas toujours, occasionne le partage de l’état affectif de cette personne » (Decety et Jackson, 2004). La composante affective de l’empathie est un terme général qui englobe un ensemble de processus (Decety et Lamm, 2006 ; de Waal et Preston, 2017). Thompson et coll. (2019) ont proposé deux classes de processus s’inscrivant dans la composante affective de l’empathie : ceux de nature perceptuelle et ceux relatifs au mimétisme. Ces deux classes de processus offrent une nomenclature intéressante qui rejoint plusieurs concepts décrits dans la littérature.
Le corps humain, et tout particulièrement le visage, déploie continuellement des mouvements qui permettent de transmettre et recevoir rapidement des informations sociales. Être en mesure de percevoir efficacement les actions, les émotions et les intentions d’autrui a une valeur adaptative importante (Peelen et Downing, 2007). L’empathie serait tributaire de la capacité à détecter les signaux socioémotionnels (Coll et coll., 2017 ; Decety
et Jackson, 2004 ; de Waal et Preston, 2017 ; Prochazkova et Kret, 2017 ; Thompson et coll., 2019). Dans le contexte de la communication de la douleur, par exemple, les gestes réflexes en réponse à une expérience douloureuse permettent d’informer un interlocuteur de la présence d’un état interne (Hadjistavropoulos et coll., 2011). Il est démontré que les signaux non verbaux, notamment les expressions faciales et les mouvements corporels, sont des indices de communication particulièrement efficaces afin d’informer autrui d’un état interne (Adolphs, 2002 ; Balconi et Bortolotti, 2013 ; Balconi et Pozzoli, 2007). En effet, les expressions faciales sont traitées implicitement, dès les stades précoces du traitement visuel des visages, et sont particulièrement saillantes comparativement à d’autres types d’informations visuelles (Fox et coll., 2000 ; Peelen et Downing, 2007). D’un point de vue développemental, il a été démontré que dès les premières années de vie, les bambins ont la capacité de percevoir les expressions faciales et de leur associer une émotion (Decety et Michalska, 2013 ; Tousignant et coll., 2017). Ceci suggère une capacité quasi innée de percevoir ce type d’indices socioémotionnels. Bien que la perception des expressions faciales ait largement été étudiée (Vuilleumier et Pourtois, 2007), l’étude de la perception des mouvements émotionnels du corps a reçu beaucoup moins d’intérêt. Quelques études indiquent que la perception des mouvements émotionnels du corps peut induire un biais attentionnel et moduler l’activité des régions cérébrales assurant le traitement visuel précoce de l’information (Peelen et Downing, 2007). Par exemple, une étude chez des patients présentant une héminégligence spatiale démontre que l’observation d’images illustrant des postures émotionnelles présentées dans l’hémichamp visuel controlatéral lésionné réduit le biais attentionnel vers ces mêmes images présentées dans l’hémichamp visuel lésionnel ipsilatéral (Tamietto, Geminiano, Genero et de Gelder, 2007). En somme, les expressions faciales et les mouvements corporels évoquant des émotions sont traités rapidement et tôt dans le processus de traitement visuel. Ceux-ci sont primordiaux à la communication sociale (Tamietto et de Gelder, 2010).
La perception des émotions exprimées par les visages ou par les mouvements du corps serait sous-tendue par l’activation de régions impliquées dans les processus précoces de la perception du visage et du corps et dans les émotions (Johnson, 2005 ; Peelen et Downing, 2007). Il existerait par ailleurs une dichotomie neuro-anatomique fonctionnelle entre la perception du visage, du corps dans son ensemble et des parties du corps dans les régions
occipito-temporales (pour une revue, voir Peelen et Downing, 2007). Les études mesurant l’activité électrique chez des sujets épileptiques suggèrent que le cortex visuel ventral, le sulcus temporal supérieur et le lobule pariétal inférieur seraient sélectifs à la perception des mains alors que l’aire extrastriée du corps (extrastriate body area; EBA) serait sélective à la perception du corps dans sa globalité (sans tête) (Peelen et Downing, 2007). Les études IRMf chez l’humain appuient également la fonction sélective de l’EBA dans la perception du corps (Downing, Jiang, Shuman et Kanwisher, 2001). L’aire occipitale du visage (occipital face area; OFA) serait également une région clé impliquée dans la perception des visages (Peelen et Downing, 2007). De récentes études en IRMf démontrent que la perception du corps et du visage activerait spécifiquement des régions dans le gyrus fusiforme. L’aire fusiforme du corps (fusiform body area; FBA) serait sélective à la perception du corps et de ses membres et l’aire fusiforme du visage (fusiform face area; FFA) serait sélective à la perception des visages (Peelen et Downing, 2007). Finalement, des régions sous-corticales, telles que le noyau pulvinar, l’amygdale, le striatum ventral et l’insula, seraient impliquées dans la perception de mouvements et des expressions faciales. Ces régions assureraient le traitement de la charge émotionnelle associée à ces indices sociaux (Johnson, 2005 ; Tamietto et de Gelder, 2010). En somme, l’EBA et l’FBA seraient sélectifs à la perception du corps, alors que l’OFA et l’FFA seraient sélectifs à la perception du visage (voir Figure 4 dans Peelen et Dawning, 2007). D’autres régions sous-corticales seraient impliquées dans la charge émotionnelle associée à la perception d’indices sociaux relatifs à l’état émotionnel d’autrui.
Le processus de mimétisme est probablement l’un des mécanismes les plus rudimentaires de l’empathie (de Waal et Preston, 2017). Il se définit comme la prédisposition innée à imiter spontanément, automatiquement et inconsciemment les comportements d’autrui (Hatfield, Bensman, Thornton et Rapson, 2014). Ce mécanisme serait contrôlé par le système nerveux autonome (SNA) et se déclinerait en deux sous-processus, soit le mimétisme moteur et le mimétisme autonomique (Prochazkova et Kret, 2017). Le mimétisme moteur est la capacité à imiter spontanément les comportements moteurs d’autrui. Il englobe les expressions faciales, les vocalisations, les postures corporelles et les mouvements oculaires (Prochazkova et Kret, 2017). De nombreuses études en électromyographie démontrent qu’en effet, la perception d’expressions faciales
élicite une activité congruente dans les groupes musculaires associés à ces expressions faciales (p. ex., Dimberg et Petterson, 2000 ; Dimberg et Thunberg, 2012 ; Maillot et coll., 2012). Par exemple, le corrugateur s’active généralement en réponse à l’observation de visages exprimant de la colère et le zygomatique s’active généralement en réponse à l’observation de visages exprimant de la joie (Dimberg et Petterson, 2000). Le mécanisme de mimétisme moteur serait relié à l’empathie (Dimberg et Thunberg, 2012 ; Maillot et coll., 2012). Une étude en électromyographie démontre que des participants ayant un score élevé en comparaison à ceux ayant un score faible à un questionnaire mesurant l’empathie dispositionnelle présentent une activité plus grande et rapide du muscule zygomatique lors de l’observation d’images illustrant des expressions faciales de joie (Dimberg et Thunberg, 2012). On définit l’empathie dispositionnelle comme la réponse empathique par défaut en tant que trait ou caractère permanent d’une personne (Brunel & Cosnier, 2012). Les participants ayant un score plus élevé au questionnaire d’empathie dispositionnelle présentaient également une activité plus grande et rapide du corrugateur lors de l’observation d’images illustrant des expressions faciales de colère (Dimberg et Thunberg, 2012). Lors d’une autre étude en électromyographie, des participants visionnaient de courtes vidéos illustrant différents niveaux d’intensité d’expressions faciales de douleur et recevaient subséquemment des stimulations nociceptives (Maillot et coll., 2012). L’observation d’expressions faciales de douleur d’intensité élevée en comparaison à l’observation d’expressions faciales de douleur de faible intensité ou neutre était associée à une augmentation de la douleur autorapportée, de l’amplitude du réflexe de flexion nociceptif et de l’activité du corrugateur (Maillot et coll., 2012). Les réponses autorapportées d’intensité et de désagrément de la douleur étaient par ailleurs modulées à la baisse chez les individus qui présentaient des niveaux plus élevés d’empathie dispositionnelle (Maillot et coll., 2012). Ces résultats suggèrent que les processus afférants d’imitation et de réflexe nociceptif sont modulés par des processus efférants de plus hauts niveaux d’empathie dispositionnelle (Maillot et coll., 2012). En somme, ces études psychophysiologiques nous enseignent que nous avons une disposition innée à imiter spontanément les comportements moteurs d’autrui en contexte émotionnel, ce qui semblerait favoriser une représentation interne des états moteurs et affectifs d’autrui.
L’empathie dispositionnelle module, à la hausse ou à la baisse, le mécanisme de mimétisme moteur selon le contexte expérimental.
La théorie du couplage neuronal entre la perception et l’action (perception-action
model; Preston et de Waal, 2002) est pour plusieurs au cœur du mécanisme de mimétisme
moteur (p. ex., Jackson et Decety, 2004 ; de Waal et Preston, 2017). Découverts par Vittorio Gallese (1996), les neurones dits « miroirs » s’activeraient tant lors de la perception que de l’exécution d’une action (Keysers et Fadiga, 2008). En effet, des études démontrent qu’effectuer des mouvements, ainsi que voir ou entendre ces mêmes actions effectuées par autrui, recruteraient de mêmes neurones chez le primate et de mêmes régions cérébrales chez l’humain (Keysers, 2006). Chez le primate, tous les neurones de la partie ventrale du cortex prémoteur (F5) s’activeraient lorsque le primate observe des actions effectuées par un individu et lorsque le primate effectue ces mêmes actions, et ce, peu importe s’il est en mesure de voir ou non ses propres actions (Gallese, Fadiga, Fogassi et Rizzolatti, 1996). On retrouverait ces neurones miroirs dans un circuit neuro-fonctionnel précis qui incluerait la partie antérieure du sulcus intrapariétal, le lobule pariétal inférieur (Keysers, 2006) et le sulcus temporal supérieur (Keysers et Gazzola, 2006). Notons toutefois qu’au sein du sulcus temporal supérieur, certains neurones s’activeraient lors de l’observation et de l’exécution d’actions. D’autres cesseraient de s’activer lors de l’exécution d’actions lorsque ces actions sont observées par l’individu exécutant ces actions (Perrett et coll., 1990). Ces neurones spécifiques auraient ainsi une fonction inhibitrice comparativement à la fonction excitatrice des neurones dans la partie ventrale du cortex prémoteur et dans le lobule pariétal inférieur (Keysers et Gazzola, 2006). En d’autres mots, une partie des neurones semblerait répondre aux actions de soi et d’autrui, alors qu’une autre partie des neurones semblerait assurer la capacité de distinguer l’origine des actions entre soi et autrui (Keysers et Gazzola, 2006). Chez l’humain, bien que le sujet soit toujours débattu, des données suggèrent que certaines régions cérébrales auraient des propriétés « miroirs », soit dans le cortex prémoteur ventral et dorsal, le lobule pariétal inférieur, l’aire motrice supplémentaire, les cortex somatosensoriels primaire (S1) et secondaire (S2), le gyrus temporal postérieur moyen, le sulcus temporal supérieur et le cervelet (Keysers, 2006 ; Keysers et Gazzola, 2006). Comme des projections neuronales sont observées de ces régions vers l’insula antérieure (IA) et que l’IA projette vers
l’amygdale, il a été proposé que l’IA assure la connexion entre le circuit associé au mimétisme moteur et les régions assurant le traitement des émotions (Prochazkova et Kret, 2017). En effet, l’IA et l’amygdale sont des régions classiquement associées aux émotions (Adolphs, 2001 ; Atkinson et Adolphs, 2005 ; Carr et coll., 2003).
Le mimétisme autonomique, le « linkage physiologique », ou encore la synchronie physiologique, se définit par la synchronisation des états autonomiques d’une personne à ceux d’une autre personne (Prochazkiva et Kret, 2017). Ce phénomène a été documenté principalement dans la dyade mère-enfant (pour une revue systématique, voir Palumbo et coll., 2016). Il englobe la synchronisation des hormones, du rythme cardiaque, de la dilatation pupillaire et du rougeoiement (Prochazkova et Kret, 2017). Par exemple, une étude a documenté le rythme cardiaque de mères et leur bambin de trois mois alors que ceux-ci étaient en interaction face-à-face (Feldman et coll., 2011). Lors de moments d’affection où les contacts visuels et les vocalisations entre les mères et leur bambin étaient synchronisés comparativement à des périodes de non-synchronisation, on observait que le rythme cardiaque des deux parties se synchronisait en un laps de temps de moins d’une seconde (Feldman et coll., 2011). Une autre étude menée auprès de triades entre des adolescents et leurs parents a démontré que les niveaux de cortisol corrélaient positivement entre les pères, les mères et les adolescents lors de discussions chargées émotionnellement (Saxbe et coll., 2014). Plus surprenant, les niveaux de cortisol étaient modérés par le niveau d’affiliation et le genre entre le parent et l’adolescent. Les niveaux de cortisol des adolescents étaient plus fortement associés au parent du même genre et au niveau d’affiliation avec la mère (Saxbe et coll., 2014).
Un circuit neuro-fonctionnel associé au mécanisme de mimétisme autonomique a été proposé dans le modèle neurocognitif de la contagion émotionnelle (Prochazkova et Kret, 2017). Celui-ci suggère que le mimétisme autonomique repose principalement sur les mêmes substrats neuronaux et circuits que ceux impliqués dans les émotions et l’activation du SNA. En situation de stress, par exemple, un individu présentera une activation plus importante de l’axe hypothalamo -hypophyso-surrénalien, qui se traduira par une augmentation du niveau de cortisol dans le sang. Celle-ci s’accompagne d’une augmentation de l’activité du SNA en parallèle à des changements cardiovasculaires, de la
tension musculaire, de la conductance électrodermale, de la dilatation pupillaire, du rougeoiement dans le visage et de la transpiration (Prochazkova et Lret, 2017). Ces fins signaux socioémotionnels seraient traités par un observateur par le colliculus supérieur et le pulvinar, et ensuite l’amygdale. L’amygdale est une région associée à la détection de mouvements biologiques, incluant les émotions exprimées par le visage ou le corps (Adolphs, 2001 ; Atkinson et Adolphs, 2005). L’activation de ce circuit activerait subséquemment l’axe hypothalamo-hypophyso-surrénalien chez l’observateur. Le circuit colliculus supérieur-pulvinar et l’amygdale, ayant des projections dans les régions corticales incluant le cortex orbito-frontal, le cortex cingulaire antérieur (CCA) et le cortex préfrontal (CPF) ventro-médial, activerait ces régions. Ces dernières assureraient subséquemment la prise de décision sociale. Par ailleurs, lorsqu’un observateur perçoit les signaux fins (dilatation pupillaire, rougeoiement du visage, transpiration) d’activation du SNA chez autrui, l’amygdale, ayant des projections vers le locus coeruleus, activerait le locus coeruleus. Le locus coeruleus, ayant des connexions avec la partie ventrale du thalamus, activerait cette région. Celle-ci activerait par la suite des régions motrices, lesquelles assureraient le comportement le plus adaptatif à effectuer (Prochazkova et Kret, 2017).
La contagion émotionnelle est un autre mécanisme décrit comme étant associé à l’empathie. La contagion émotionnelle réfère à la tendance à s’imprégner des états sensoriels, moteurs, physiologiques et affectifs d’autrui (Preston et de Waal, 2002). Elle émanerait de l’interaction entre plusieurs types de processus de mimétisme (Hatfield et coll., 1994). La contagion émotionnelle demeurerait un mécanisme automatique, involontaire, incontrôlable et en grande partie, inconscient (Hatfield et coll., 2014). De nombreuses études démontrent que, comparativement à l’observation de visages, reproduire, exagérer ou imiter des expressions faciales occasionne une plus grande réponse émotionnelle, telle que mesurée au plan subjectif, comportemental et physiologique (Hatfield et coll., 2014). Par exemple, une étude en électromyographie démontre que l’amplitude du mouvement de mimétisme faciale est reliée positivement à une mesure autorapportée de partage émotionnel entre soi et autrui (Hess et Blairy, 2001). Une autre étude démontre qu’une augmentation de l’activité du zygomatique et du corrugateur est associée à une augmentation de comportements prosociaux d’aide de même qu’associée à
l’empathie dispositionnelle (Balconi et Canavesio, 2013). La contagion émotionnelle est, pour certains théoriciens, l’expression la plus simple de la composante affective de l’empathie (Decety et Jackson, 2004), et pour d’autres, l’une des formes les plus rudimentaires d’empathie (de Waal et Preston, 2017).
La théorie du couplage neuronal entre la perception et l’action permettrait d’expliquer le mécanisme de contagion émotionnelle (Jackson et Decety, 2004 ; de Waal et Preston, 2017). En effet, il a été suggéré que l’observation d’un état émotionnel chez une personne activerait automatiquement les représentations neuronales chez l’observateur de cette émotion en simultané aux réponses motrices et autonomiques (Bernhardt et Singer, 2012 ; Keysers et Gazzola, 2010). Ce phénomène est communément appelé résonance neuronale et a été documenté dans plusieurs études sur l’observation de la douleur (p. ex., Jackson et coll., 2005 ; 2006). La résonance neuronale serait en partie sous-tendue par les régions cérébrales ayant des propriétés « miroirs ». Par exemple, une lésion des cortex somato-sensoriels droits a été associée à une diminution de la capacité à reconnaître des émotions de base par l’observation d’expressions faciales, telles que la joie, la surprise, la colère, la tristesse et la peur (Adolphs, Damasio, Tranel, Cooper et Damasio, 2000). Il est intéressant de remarquer que les régions associées aux neurones miroirs, telles que le lobule pariétal inférieur, l’aire motrice supplémentaire, le cortex prémoteur dorsal et les cortex somatosensoriels SI et SII, se retrouvent presque systématiquement activées dans différents paradigmes en IRMf portant sur l’empathie pour la douleur (pour des méta-analyses, voir Lamm et coll., 2011 ; Timmers et coll., 2018) et l’empathie pour les émotions positives et négatives (Fan et coll., 2011). Les différences d’activation dans les régions associées aux neurones miroirs seraient également reliées aux différences individuelles d’empathie dispositionnelle (Pfeifer et Dapretto, 2011). Lors d’une étude en IRMf, l’activité BOLD d’adolescents a été mesurée alors que ceux-ci observaient ou imitaient diverses expressions faciales. Les résultats démontraient que l’activité du pars opercularis dans le lobule pariétal inférieur corrélait fortement et positivement avec l’empathie dispositionnelle, telle que mesurée par le Interpersonal Reactivity Index (Davis, 1983;Pfeifer, Iacoboni, Mazziotta et Depretto, 2008).
La théorie des neurones miroirs fait toutefois l’objet de critiques à ce jour (Krishnan et coll., 2016). En effet, il importe de mentionner que les études sur lesquelles se basent cette théorie utilisent des techniques d’imagerie cérébrale dont la résolution spatiale ne permet pas de conclure directement sur l’aspect indissociable des réseaux cérébraux sollicités lors de l’expérience et l’observation d’un état affectif (Oosterhof, Tipper et Downing, 2013). Par des analyses de patrons mutivoxels et somato-topiques, Krishnan et coll. (2016) ont en effet identifié deux réseaux fonctionnels dissociables associés spécifiquement à l’expérience et à l’observation de la douleur. L’expérience de la douleur était associée à un circuit somato-sensoriel, lequel incluait l’insula antérieure (IA), l’insula dorso-postérieure, le CCA dorsal et l’air somato-sensoriel. L’observation de la douleur était associée à l’activation du CPF dorso-médial, de l’amygdale, du cortex cingulaire postérieur et de la jonction temporo-pariétale. Les nouvelles techniques d’analyse des données en IRMf suscitent un intérêt croissant à l’heure actuelle. Elles permettent de détecter de manière plus fine des différences d’activité cérébrale entre des conditions expérimentales comparativement aux analyses univariées conventionnelles. Bien qu’à ce jour la notion de neurones miroirs soit critiquée, elle a grandement influencé notre compréhension de l’empathie et des processus sous-jacents.
En somme, le modèle neurocognitif de la contagion émotionnelle de Prochazkova et Kret (2017) permet de lier les mécanismes de mimétisme moteur et autonomique à la contagion émotionnelle, au couplage neuronal et à l’empathie. Celui-ci stipule que lors d’interactions socioémotionnelles, l’état émotionnel d’une personne est reflété par de subtils indices tels que des mouvements non-verbaux (expressions faciales, posture, contact visuel) et des changements de l’activité du SNA (augmentation du rythme cardiaque, changement de la couleur du visage, dilatation pupillaire). Ces signaux sont captés par l’observateur et influenceront sa propre réponse physiologique de même que les réponses affectives et cognitives associées. Ces mécanismes seraient sous-tendus par une activation neuronale partagée entre une personne exprimant une émotion et un observateur, permettant à celui-ci de simuler les aspects sensorimoteurs correspondant aux émotions d’autrui. L’activation de l’une des composantes d’une émotion, qu’elle soit motrice ou autonomique, activera ses autres composantes motrices ou autonomiques. La conjonction des différentes informations serait essentielle à l’activation des représentations internes de