Isolement et caractérisation des métapyrocatchases induites par le biphényle chez comamonas testosteroni B-356 et essai de clonage de la catéchol 2,3-dioxygénase.
Texte intégral
Figure

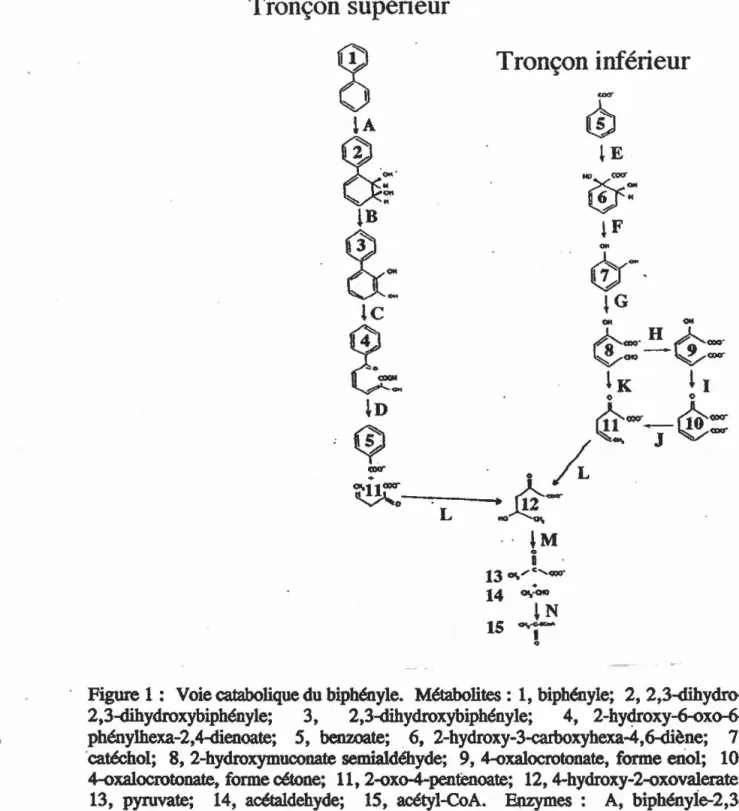
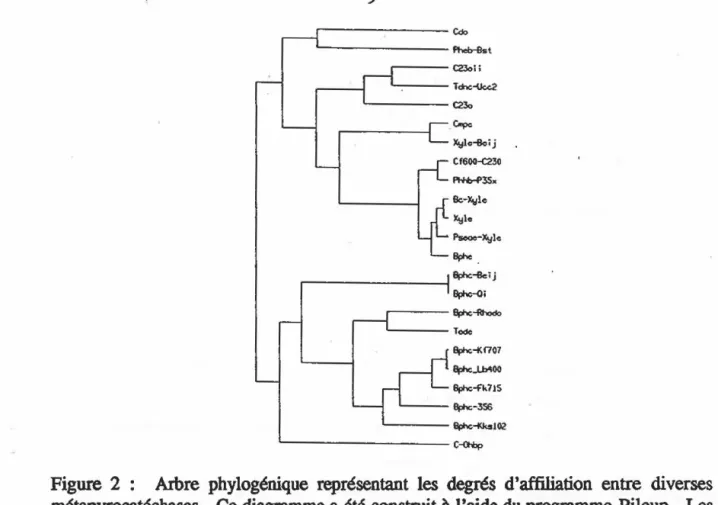

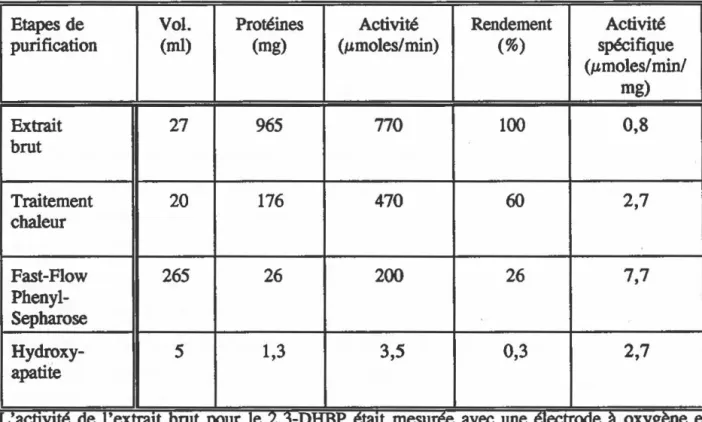

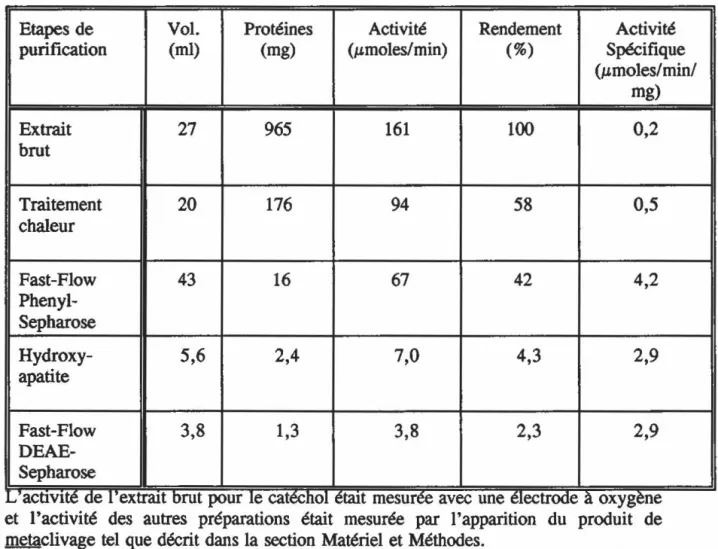



Documents relatifs
Side events/meetings, Work package meetings, or free-time. ALTER-Net projects get a chance to meet the LTER site and network representatives to explore how they can help
رﻻود نوﯾﻠﻣ (. - تﺎﯾﺎﻔﻧﻟا ﻰﻠﻋ ءﺎﺿﻘﻟا ندﻣﻟا نﻣ برﻘﻟﺎﺑ ﺔﻋوﻧﻣﻣﻟا ىرﺑﻛﻟا. - ﺔﺋﯾﺑﻟا نﮭﻣﻟ دﮭﻌﻣ ﻊﺿو.. - ﻊﻣ ﻲﺋﯾﺑ ﻲﺳﯾﺳﺣﺗ ﺞﻣﺎﻧرﺑ قﯾﻘﺣﺗ سرادﻣﻟا ﻲﻓ ﺔﯾﺋﯾﺑﻟا ﺔﯾﺑرﺗﻟا جﺎﻣدإ
179 Tableau 30 – Evaluation statistique de la relation liant l’âge et la parité des patientes avec les résultats histologiques sur pièces de conisations………..…………..……136
Pour mesurer la réponse des vitroplants suite aux différents traitements (LAM, MeJa, SA et T), les quantités de trois hormones impliquées dans les réponses de défenses
Monitoring Land Use/Land Cover Changes in Urban Areas Using Medium Spatial Resolution Satellite Imagery: the Case of the Montreal Metropolitan Area.. (Suivi des changements
The model is based on a model of concurrent aspects, CEAOP [11], and is provided in the form of a language for modeling the adaptation of applications to context changes.. The
0,0 1,0 2,0 3,0 4,0 5,0 6,0 7,0 8,0 9,0 Libre Collé Te m ps e n jo ur s POSE INITIALE Dépose du sol Reconnaissance / Préparation de support Séchage Implantation Mise en œuvre
Mais ça me rend peut-être moins patiente avec les gens autour de moi, et puis même après dans des situations banales de la vie quotidienne si je pense au boulot et que je suis
