Mise au point d’une technique de pulvérisation des
pupes de trichogramme pour la lutte biologique contre
la pyrale dans la culture du maïs sucré
Mémoire
Pascal Gauthier
Maîtrise en génie agroalimentaire
Maître ès sciences (M.Sc.)
Québec, Canada
Mise au point d’une technique de pulvérisation des
pupes de trichogramme pour la lutte biologique contre
la pyrale dans la culture du maïs sucré
Mémoire
Pascal Gauthier
Sous la direction de :
Mohamed Khelifi, directeur de recherche
iii
Résumé
La pyrale du maïs, Ostrinia nubilalis (Hübner), est le principal ravageur du maïs sucré au monde. Sa présence cause d’importantes baisses de rendement. Actuellement, ce sont les pesticides qui sont principalement utilisés pour lutter contre ce ravageur. Les effets néfastes des pesticides sur la santé et sur l’environnement sont bien connus et toujours d’actualité. Malgré tout, des alternatives viables existent. Depuis quelques années, la lutte biologique utilisant les trichogrammes a permis une répression efficace de la pyrale sans les effets néfastes des pesticides. Toutefois, bien qu’efficace, l’utilisation des trichogrammes est plus coûteuse et fastidieuse que l’utilisation des pesticides. C’est pourquoi la pulvérisation des pupes de trichogrammes est une alternative permettant de rendre la lutte biologique plus facile et moins coûteuse que les méthodes habituelles utilisées pour introduire les trichogrammes au champ. La faisabilité de l’introduction inondative de l’espèce Trichogramma ostriniae par pulvérisation a été évaluée en laboratoire et a débuté par la recherche d’une solution efficace de dispersion et d’adhésion. Une solution aqueuse de 3 g/L de gomme xanthane et 3 g/L de gomme de guar a permis d’obtenir une solution visqueuse dispersant bien les pupes, les collant au feuillage et offrant une bonne résistance à une pluie simulée en laboratoire. Un système de pulvérisation a par la suite été conçu et testé pour vérifier la viabilité des pupes une fois immergées et pulvérisées. Le taux d’émergence des pupes pulvérisées en laboratoire a dépassé les 80% et a été comparable à celui du témoin. Le pulvérisateur a aussi été testé sur des plants de maïs. Une excellente adhésion des gouttelettes au feuillage du maïs a été observée. Par contre, la résistance à la pluie de la solution doit être évaluée en situation réelle. La survie des pupes devrait aussi être vérifiée pour confirmer l’efficacité de cette méthode de lutte biologique.
iv
Abstract
The European corn borer, Ostrinia nubilalis (Hübner), is the major insect pest of sweet corn in the world. Its presence results in important yield decreases. Currently, chemical insecticides are mainly used to control this insect pest. Chemical hazards for health as well as the environment are well documented and still relevant. Nevertheless, some potentially viable alternatives exist. For many years, the biological control using trichogramma has been successful in controlling the corn borer without the adverse effects of chemical pesticides. This biological control method is however more expensive and time and energy consuming than chemical insecticides. For this reason, spraying trichogramma pupae is an easier and inexpensive alternative than the common methods used to introduce trichogramma in the fields. Technical feasibility of inundative release by spraying
Trichogramma ostriniae has been investigated in the laboratory. The first trials aimed at
finding an adequate aqueous solution to both scatter the pupae in the sprayer tank and stick them to corn leaves once sprayed. An aqueous solution of 3 g/L of xanthan gum and 3 g/L of guar gum allowed obtaining a viscous solution which disperses well the pupae in the sprayer tank, sticks them on corn leaves, and offers a good resistance to simulated rain in the laboratory. Thereafter, a spraying system was designed and built to check the viability of trichogramma pupae after being immersed in the spraying solution and sprayed. Results showed that the sprayed pupae emergence rate exceeded 80% and was comparable to that of unsprayed pupae. The spraying system was also tested on corn crops. An excellent adhesion of droplets to corn foliage was observed. However, rain resistance of the spraying solution must be evaluated under real conditions. Pupae survival should also be checked to confirm the effectiveness of this biological control method.
vi
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... vi
Liste des tableaux ... ix
Liste des figures ... x
Remerciements ... xiii
Avant-propos ... xiv
Chapitre 1 Introduction ... 1
Chapitre 2 Revue de littérature ... 2
2.1 Maïs ... 2
2.1.1 Historique du maïs ... 2
2.1.2 Hybridation ... 3
2.1.3 Pratiques culturales ... 5
2.1.4 Culture du maïs au Canada ... 5
2.2 Pyrale du maïs (Ostrinia nubilalis) ... 6
2.2.1 Stades de développement, dommages causés et échantillonnages de la pyrale du maïs ... 7
2.3 Pesticides ... 10
2.3.1 Évaluation du danger des pesticides ... 10
2.3.2 Indice de risque pour la santé ... 10
2.3.3 Indice de risque pour l’environnement ... 12
2.3.4 Résistance aux pesticides ... 12
2.4 Génie génétique ... 15
2.4.1 Maïs Bt ... 16
2.4.2 Refuge de maïs non-Bt ... 16
2.5 Agriculture biologique ... 17
2.5.1 Marché biologique ... 17
2.6 Lutte biologique aux phytophages ... 19
2.6.1 Biopesticides ... 19
2.6.2 Parasitoïdes ... 20
2.6.3 Trichogrammes ... 21
vii 2.7.1 Degré-jour ... 24 2.7.2 Trichocartes ... 25 2.8 Distribution sèche ... 27 2.9 Distribution humide ... 29 2.10 Principe de pulvérisation ... 37 2.10.1 Débit et pression ... 38
2.10.2 Diamètre moyen des gouttelettes ... 38
2.10.3 Caractérisation des gouttes ... 39
2.10.4 Volume median diameter ... 39
2.10.5 Number median diameter ... 40
2.11 Pompes ... 41 2.11.1 Équation de Bernoulli ... 41 2.11.2 Puissance de pompe ... 42 2.11.3 Efficacité ... 42 2.11.4 Cavitation ... 42 2.11.5 NPSH ... 43 2.11.6 Pompes volumétriques ... 43 2.11.7 Pompe centrifuge ... 45
Chapitre 3 Hypothèse et objectifs ... 47
3.1 Hypothèse de recherche ... 47
3.2 Objectifs ... 47
Chapitre 4 Matériel et méthodes ... 48
4.1 Expérience en laboratoire ... 48
4.1.1 Gomme xanthane et gomme de guar ... 48
4.1.2 Immersion des pupes de trichogrammes dans la gomme xanthane et la gomme de guar ... 48
4.1.3 Analyse statistique des données des essais d’immersion ... 50
4.1.4 Viscosité des solutions ... 50
4.1.5 Pulvérisation des solutions en laboratoire ... 50
4.1.6 Pulvérisation des pupes de trichogrammes en laboratoire ... 51
4.1.7 Essai du premier pulvérisateur ... 54
4.1.8 Essai du deuxième pulvérisateur ... 55
viii
4.2.1 Première pulvérisation au champ ... 58
4.2.2 Deuxième pulvérisation au champ ... 60
4.2.3 Troisième pulvérisation au champ ... 60
4.3 Adhésion des pupes de trichogrammes au feuillage ... 60
4.3.1 Colle blanche diluée ... 60
4.3.2 Gomme xanthane concentrée et mélange de gomme xanthane et de guar . 61 Chapitre 5 Résultats et discussions ... 63
5.1 Immersion des pupes de trichogrammes dans la gomme xanthane et la gomme de guar ... 63
5.2 Analyse statistique des données des essais d’immersion ... 63
5.3 Viscosité des solutions ... 65
5.4 Essai du premier pulvérisateur ... 66
5.5 Essai du deuxième pulvérisateur ... 68
5.5.1 Première pulvérisation au champ ... 69
5.5.2 Deuxième pulvérisation au champ ... 70
5.5.3 Troisième pulvérisation au champ ... 71
5.6 Adhésion des pupes de trichogramme au feuillage ... 74
5.6.1 Colle blanche diluée ... 74
5.6.2 Gomme xanthane concentrée et mélange de gomme xanthane et de guar . 75 Chapitre 6 Conclusions et recommandations ... 81
6.1 Conclusions ... 81
6.2 Recommandations ... 82
Bibliographie ... 84
Annexe 1 Données recueillies lors des essais ... 89
ix
Liste des tableaux
Tableau 2.1: Production mondiale et canadienne de différentes céréales et graines
oléagineuses pour l’année 2011 (Hamel et Dorff, 2014). ... 2
Tableau 2.2: Nombre de fermes et superficies ensemencées au Canada et dans différentes provinces en 1971 et 2011 (Statistique Canada, 2011) ... 6
Tableau 2.3: Répartition du maïs –grain et du maïs sucré selon la culture traditionnelle et la certification biologique au Canada en 2011 (Statistique Canada, 2011) ... 16
Tableau 2.4 : Coût saisonnier par hectare pour l’utilisation de différentes méthodes de contrôle de la pyrale du maïs au Québec (Boisclair et al., 2011) ... 26
Tableau 2.5: Émergence des adultes Trichogramma pretiosum pour les traitements et le témoin avec un intervalle de confiance de 95% (Morrison et al., 1998) ... 30
Tableau 2.6: Pourcentage d’émergence et temps requis pour différentes méthodes de dispersion (Jalali et al., 2005) ... 33
Tableau 2.7 : Nombre théorique de gouttes/cm2 en fonction de leur diamètre et du volume/ha (Barthelemy et al., 1990) ... 41
Tableau 5.1: Résultats de l’ANOVA des données d’immersion dans les solutions de gomme de guar (GG) et de gomme xanthane (GX) ... 64
Tableau 5.2: Résultats des contrastes pour l’essai d’immersion dans la gomme de guar et la gomme xanthane ... 64
Tableau 5.3 : Résultats du test de Kruskal-Wallis pour l’essai du deuxième pulvérisateur ... 69
Tableau 5.4: Résultats de l’ANOVA pour l’essai d’adhésion ... 77
Tableau 5.5: Comparaison des variables feuille et traitement ... 77
Tableau 5.6: Résultats du test de Kruskal-Wallis pour l’essai d’adhésion ... 78
Tableau 5.7: Test exacte de Wilcoxon pour comparaison entre le témoin et la GX à 4 g/L pour l’essai d’adhésion ... 78
Tableau 5.8: Test exacte de Wilcoxon pour comparer de l’adhésion du témoin avec celle du mélange de gomme xanthane 2 g/L et de gomme de guar 2 g/L ... 79
Tableau 5.9 : Test exacte de Wilcoxon pour comparer l’adhésion du traitement de gomme xanthane 4 g/L avec celle du mélange de gomme xanthane 2 g/L et de gomme de guar 2 g/L pour l’essai d’adhésion ... 80
x
Liste des figures
Figure 2.1: Données sur la production mondiale de maïs en 2012-2013 (Société
plantureux et associés, 2014). ... 3 Figure 2.2: Évolution du rendement des hybrides de maïs aux États-Unis de 1865 jusqu’à 2005 (Bennetzen et Hake, 2009). ... 4 Figure 2.3 : Pyrale du maïs mâle à gauche et femelle à droite au stade adulte (Ferland et al., 2000). ... 7 Figure 2.4 : Stades de développement en fonction du temps pour les races univoltine et bivoltine de la pyrale du maïs (Boisclair et Jean, 2009). ... 8 Figure 2.5: Œufs de pyrale du maïs et jeunes larves (Brodeur, 2008). ... 9 Figure 2.6 : Larves de pyrale ayant creusées dans une tige et dans des épis de maïs (Hagerman, 1997). ... 9 Figure 2.7: Représentation schématique des différentes résistances aux pesticides chez un insecte (Haubruge et Amichot, 1998). ... 13 Figure 2.8 : Processus biochimique de métabolisation des composés toxiques absorbés par l’insecte (Haubruge et Amichot, 1998). ... 15 Figure 2.9: Estimations de l’évolution du marché total des produits biologiques au Canada de 2000 à 2010 (ÉcoRessources Consultants, 2012). ... 18 Figure 2.10 : Femelle Trichogramma ostriniae parasitant des œufs de la pyrale du maïs (Marchal, 1936). ... 21 Figure 2.11 : Cycle de production et d’utilisation des trichogrammes dans la lutte
biologique contre la pyrale du maïs (Hawlitzky, 1992). ... 24 Figure 2.12 : Une trichocarte contenant des œufs parasités par les trichogrammes sur un plant de maïs (Revue UFA, 2014). ... 25 Figure 2.13: Système d’ouverture des contenants (Khelifi et al., 2011). ... 28 Figure 2.14 : Taux d’émergence des œufs de pyrale de la farine parasités par
Trichogramma pretiosum après immersion et agitation dans l’eau jusqu’à 180 minutes
(cercles) et immersion et agitation jusqu’à 60 minutes puis pulvérisation (carrés). Les barres indiquent l’erreur type et les lignes une régression par la méthode des moindres carrés (Gardner et Giles, 1997). ... 31 Figure 2.15 : Concentration d’œufs de pyrale de la farine parasitée par Trichogramma
pretiosum pulvérisés sur une période de 60 minutes. Les barres indiquent l’erreur type et
la ligne une régression par la méthode des moindres carrés. La pente est significativement différente de zéro (Gardner et Giles, 1997). ... 32 Figure 2.16 : Taux d’émergence des trichogrammes sur et sous les feuilles après
distribution dans le nuage de pulvérisation à l’aide d’un pulvérisateur rotatif à verger
(Zimmermann et Wührer, 2010). ... 35 Figure 2.17 : Taux d’émergence des trichogrammes pour différentes solutions et
pressions de pulvérisation (Kienzle et al., 2012). ... 36 Figure 2.18: Schéma de fonctionnement d’une pompe à membrane pneumatique
(Delange, 2008). ... 44 Figure 2.19 : Principe de fonctionnement d’une pompe à vis hélicoïdale (Bornemann Pumps, 2005). ... 44 Figure 2.20 : Pompe à lobes avec rotors à trois lobes (INOXPA, 1999). ... 45
xi
Figure 2.21 : Principe de fonctionnement d’une pompe centrifuge (Jouve, 2008). ... 46 Figure 4.1 : Équipements utilisés pour l’expérience d’immersion dans la gomme xanthane. ... 49 Figure 4.2 : À gauche, tamis utilisé pour rincer de la solution visqueuse et à droite,
l’entonnoir avec le papier filtre pour recueillir les pupes. ... 49 Figure 4.3: Réservoir rectangulaire de 23 L utilisé pour les essais de pulvérisation en laboratoire. ... 52 Figure 4.4: À gauche, la pompe à diaphragme SHURflo de 5,7 L/min et 12 VDC. À droite, la source de courant continu de 13,8 V. ... 53 Figure 4.5: Régulateur de pression à piston de marque Spraying System Co. et de
modèle 23120 (Spraying System Co.). ... 54 Figure 4.6: Buses TeeJet XR 110 VP modèles 02 à 06 et la buse originale de la lance de pulvérisation. ... 54 Figure 4.7 : Parcelles de maïs sucré utilisées pour les essais de pulvérisation des
trichogrammes. Chacune des parcelles comporte trois rangs de 10 plants; les parcelles sont espacées par 75 m de maïs de la même variété et traités aux pesticides. ... 56 Figure 4.8: À gauche, un pulvérisateur à dos modifié pour accueillir : a) la pompe, b) le manomètre, c) l’interrupteur et d) la batterie. À droite, la rampe à cinq buses adaptée sur la poignée d’origine du pulvérisateur. ... 58 Figure 4.9: Plat de pétri contenant approximativement 4 000 pupes de trichogramme. L’amas de pupes a été divisé en quatre pour en utiliser 1 000 à chacune des
pulvérisations. ... 58 Figure 4.10: Pulvérisation des trichogrammes au champ le 29 juillet 2015. ... 59 Figure 4.11: Parcelles protégées de la pulvérisation de pesticides. ... 59 Figure 4.12 : Montage expérimental des trois traitements (témoin, xanthane et xanthane + guar) en trois réplicas pour le dessus (U) et le dessous (D) des feuilles de maïs. ... 62 Figure 5.1 : Taux d’émergence en fonction du temps pour l’immersion des pupes dans des solutions de gomme de guar et de xanthane. Les barres d’erreurs représentent la valeur de l’écart type autour de la moyenne. ... 63 Figure 5.2: Viscosité apparente en fonction de la vitesse de rotation de la tige pour les solutions de GX 3 g/L, GX 4 g/L, GG 6 g/L et du mélange de GX 2 g/L + GG 2 g/L à 20oC. ... 66 Figure 5.3: Taux d’écrasement des pupes de trichogrammes en fonction du temps de fonctionnement de la pompe. ... 67 Figure 5.4: Taux d’émergence pour les traitements effectués avec le deuxième
pulvérisateur avec une rampe à cinq buses et une pression de 15 psi (buse 1 = buse 04 et buse 2 = buse 06). ... 68 Figure 5.5: Gouttes obtenues par la pulvérisation de la solution de gomme xanthane à 3 g/L. À droite, la viscosité des gouttes leur permet de bien adhérer sur le feuillage même à la verticale. ... 69 Figure 5.6: Première observation de la pulvérisation 2 (Pu2) le 7 août pour les deux groupes de pupes (N1 et N2) de la parcelle 1 (P1) et les deux groupes (N3 et N4) de la parcelle 2 (P2). ... 70
xii
Figure 5.7: Deuxième observation de la pulvérisation 2 (Pu2) le 10 août pour les deux groupes de pupes (N1 et N2) de la parcelle 1 (P1) et les deux groupes (N3 et N4) de la parcelle 2 (P2). ... 71 Figure 5.8: Première observation de la pulvérisation 3 (Pu3) le 13 août pour les deux groupes de pupes (N1 et N2) de la parcelle 1 (P1) et les deux groupes (N3 et N4) de la parcelle 2 (P2). ... 72 Figure 5.9: Deuxième observation de la pulvérisation 3 (Pu3) le 17 août pour les deux groupes de pupes (N1 et N2) de la parcelle 1 (P1) et les deux groupes (N3 et N4) de la parcelle 2 (P2). ... 73 Figure 5.10: Taux d’adhérence au feuillage des pupes suite aux trois arrosages pour la gomme xanthane seule (GX 3 g/L), le mélange no.1 avec de la colle blanche (GX 3 g/L + CB 1 goutte/10 mL) et le mélange no.2 avec de la colle blanche (GX 3 g/L + CB 1
goutte/20 mL). ... 74 Figure 5.11: Taux d’adhésion pour le témoin (eau), la gomme xanthane seule (GX 4 g/L) et le mélange de gomme xanthane et gomme de guar (GX 2 g/L + GG 2 g/L). ... 75 Figure 5.12: Échantillons pour le dessus (U) et le dessous (D) des feuilles pour la
répétition 3 (R3) du traitement à la gomme xanthane 4 g/L (X). ... 76 Figure 5.13: Échantillons pour le dessus (U) et le dessous (D) des feuilles pour la
xiii
Remerciements
Je tiens d’abord à remercier mon directeur de recherche, le Dr Mohamed Khelifi, pour ses judicieux conseils et la confiance accordée à mon égard dès le début du projet. Sa disponibilité et son désir de partager ses connaissances ont permis de solutionner plusieurs problèmes lors de l’avancement du projet.
Je tiens aussi à remercier le principal collaborateur au projet, Anatis Bioprotection inc., pour avoir fourni les pupes de trichogrammes et l’expertise en entomologie. De plus, les conseils de l’entomologiste Mylène St-Onge ont été très utiles pour la préparation des essais au champ.
Un grand merci au Dr Conrad Cloutier pour l’enseignement des théories sur le contrôle naturel des populations d’insectes.
Je tiens aussi à remercier Francis Demers, étudiant au deuxième cycle en statistique et consultant au service de statistique de l’Université Laval.
xiv
Avant-propos
Ce projet de recherche a été financé par le Programme Innov’action agroalimentaire du MAPAQ qui est issu d’un partenariat fédéral-provincial. Ce programme vise, entre autres, à améliorer les méthodes de lutte aux ravageurs en agriculture. Ce mémoire de maîtrise est constitué de six chapitres. Le premier chapitre consiste en une introduction sur le sujet du mémoire. Le deuxième chapitre comporte une revue de littérature exhaustive sur la culture du maïs, ses ravageurs, les méthodes de lutte conventionnelles, la lutte biologique et les méthodes de distribution d’insectes bénéfiques. Le troisième chapitre présente l’hypothèse de recherche et les objectifs du projet. Le quatrième chapitre traite du matériel et de la méthodologie utilisés lors des essais. Le cinquième chapitre présente les résultats et discussion des différents essais en laboratoire et au champ. Le sixième chapitre consiste en des conclusions sur les travaux effectués et des recommandations pour la suite du projet.
1
Chapitre 1
Introduction
La popularité actuelle de lutte biologique aux phytophages est principalement due à une sensibilisation de la société envers les dangers des pesticides. Leurs effets néfastes pour la santé humaine et pour l’environnement sont maintenant bien connus. Bien que cette prise de conscience soit réelle, l’utilisation des pesticides croit sans cesse au Canada. La lutte biologique est une alternative efficace, mais est généralement plus coûteuse et laborieuse que l’utilisation des pesticides chimiques. Les méthodes d’utilisation des agents biologiques doivent être améliorées pour faciliter leur utilisation et leur acceptation par les agriculteurs. L’utilisation des trichogrammes pour lutter contre la pyrale du maïs en est un bon exemple. Bien qu’efficace, les pupes de trichogrammes doivent être apposées sur des cartons cirés avant d’être déposées manuellement dans les champs de maïs sucrés. Ces étapes sont coûteuses et exigent beaucoup de temps à l’agriculteur.
L’objectif principal de ce projet de recherche visait à développer une technique de pulvérisation des pupes de trichogramme de l’espèce Trichogramma ostriniae pour la lutte biologique contre la pyrale du maïs dans la culture du maïs sucré. Les méthodes de pulvérisation étant bien connues des agriculteurs, la pulvérisation des trichogrammes permettra de rendre la lutte biologique plus facilement acceptable.
La faisabilité technique de la pulvérisation a été évaluée en laboratoire. Différentes solutions de pulvérisation ont été étudiées afin de permettre une bonne dispersion des pupes dans le liquide et une bonne adhésion aux plants de maïs. La résistance à la noyade des pupes de trichogrammes a aussi été évaluée ainsi que l’effet de la pulvérisation sur le taux d’émergence.
2
Chapitre 2
Revue de littérature
2.1 Maïs
2.1.1 Historique du maïs
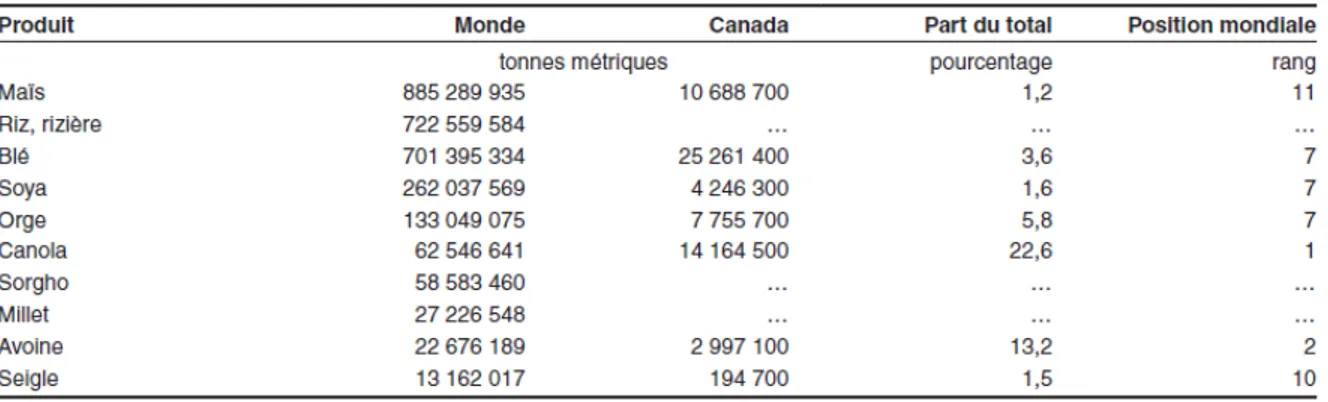
Le maïs (Zea mays) appartient à la famille des graminées. Il est cultivé en Amérique depuis des milliers d’années et serait originaire du Mexique. Son ancêtre, le Téosinte, était une sorte d’épi avec un seul rang de grains. La domestication a permis de sélectionner les meilleurs plants et d’obtenir avec le temps le maïs que l’on connait aujourd’hui. Il est la céréale la plus cultivée au monde devant le riz, le blé et le soya (Hamel et al., 2014). Il est apprécié pour son rendement plus élevé que le blé et ses qualités nutritionnelles et gustatives. Le tableau 2.1 montre les quantités annuelles des principales céréales cultivées au monde.
Tableau 2.1: Production mondiale et canadienne de différentes céréales et graines oléagineuses pour l’année 2011 (Hamel et Dorff, 2014).
Au tableau 2.1, on constate que la quantité de maïs produite en 2011 s’élève à plus de 885 000 000 tonnes métriques. Le maïs est donc d’une importance capitale dans l’économie mondiale, car il offre de bons rendements en plus d’être utilisé pour l’alimentation humaine et animale, dans la fabrication d’objet et la production de bioénergie. La figure 2.1 présente la répartition de la production de maïs dans le monde en 2012 et 2013.
3
Figure 2.1:Données sur la production mondiale de maïs en 2012-2013 (Société plantureux et associés, 2014).
La figure 2.1 permet de constater qu’en 2012-2013, les États-Unis étaient le plus gros producteur avec 35% de la production mondiale. Ils ont aussi consommé 42% de leur production pour produire de l’éthanol. Pour la même période, la Chine arrive au deuxième rang avec 22% de la production mondiale.
2.1.2 Hybridation
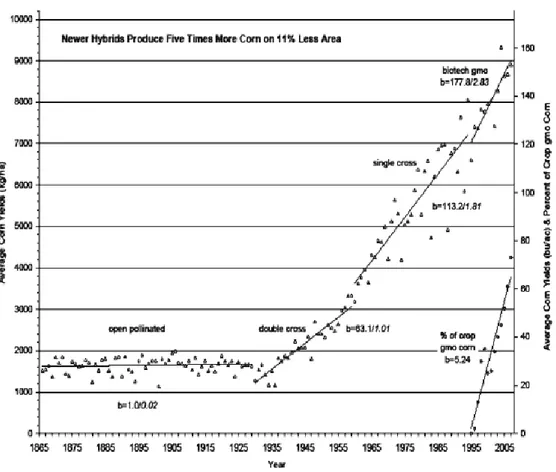
Le maïs a été cultivé avant les années 1930 par pollinisation ouverte, permettant au pollen de plusieurs variétés de se combiner. La sélection de meilleures semences se faisait donc par essai et erreur sur une longue période de temps. Par la suite, une meilleure compréhension de la génétique a permis de sélectionner les variétés pour obtenir des performances plus élevées. La figure 2.2 permet de constater cette augmentation du rendement en fonction des différentes technologies utilisées.
4
Figure 2.2: Évolution du rendement des hybrides de maïs aux États-Unis de 1865 jusqu’à 2005 (Bennetzen et Hake, 2009).
La figure 2.2 montre que les nouveaux hybrides produisent environ cinq fois plus de maïs sur une surface 11% moins grande par comparaison aux variétés d’avant 1925. De plus, le pourcentage d’utilisation du maïs génétiquement modifié a rapidement augmenté depuis son introduction vers 1995 pour atteindre une utilisation dans environ 60% des champs de maïs américains en 2005. Cette figure montre aussi qu’avant 1925, les rendements ont toujours été faibles par comparaison à ceux obtenus au demi-siècle suivant. Cela s’explique par le fait que les variétés utilisées étaient à pollinisation ouverte, ce qui faisait constamment varier le bagage génétique des grains de maïs qui pouvaient provenir de pollen différents. Comme un épi de maïs contient des centaines de grains, son utilisation comme semence engendre des plants aux gènes différents. À partir des années 1930, le travail d’hybridation de lignées pures a permis d’obtenir des plants de maïs avec de meilleures propriétés.
5 2.1.3 Pratiques culturales
Au Québec, le maïs est généralement semé à la fin du mois d’avril où lorsque la température du sol atteint au moins 10oC. Une température du sol adéquate lors du semis permet une germination rapide et évite des problèmes de pourriture des semences. Le sol doit être bien drainé et maintenu à un pH entre 6,2 et 6,5. Il doit aussi être riche en nutriment, particulièrement en azote; une fertilisation ou une rotation des terres avec des légumineuses est donc nécessaire pour maintenir un rendement maximal. Des études ont démontré que la culture du maïs après une récolte de soya permettait un rendement supérieur de 10% environ par comparaison à du maïs en monoculture (Centre de lutte antiparasitaire, 2006).
Les mauvaises herbes doivent aussi être contrôlées, car elles entrent en compétition avec les jeunes plants de maïs en captant l’eau du sol et les nutriments en plus d’être l’hôte de maladies et d’insectes ravageurs.
2.1.4 Culture du maïs au Canada
Au Canada, trois types de maïs sont cultivés. Le premier est le maïs-grain dont les grains sont moulus et utilisés pour la consommation humaine et animale et aussi comme matière première industrielle. Le second type de maïs est celui à ensilage. Dans ce cas, l’épi, la tige et les racines sont utilisées pour l’alimentation animale. Le troisième type est le maïs sucré qui est utilisé pour la consommation humaine et vendu en épis, en conserve ou surgelé. C’est ce type qui possède les meilleures qualités gustatives, car il est généralement assez sucré. Selon les recensements d’Agriculture Canada de 2011, 32 300 fermes ont ensemencé 1,63 millions d’hectares de maïs pour une valeur en maïs-grain de 2,08 milliards de dollars. Cela fait du maïs la troisième culture en importance en termes de valeur après le canola et le blé (Hamel et al., 2014). Au Québec, le maïs est la céréale la plus cultivée en 2015 avec une production de 364 000 ha de maïs-grain pour un rendement de 10,33 t/ha. Le maïs fourrager représente 59 500 ha de culture la même année (Institut de la statistique du Québec, 2015).
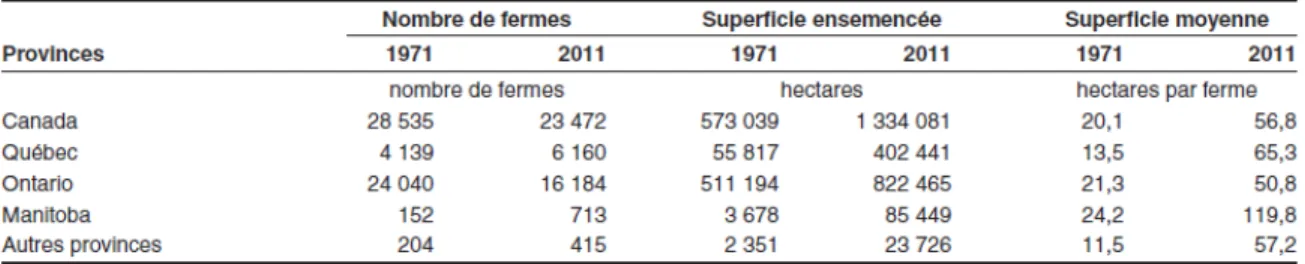
Comme partout au monde, les méthodes de cultures canadiennes ont évolué avec le temps. Par le passé, un plus grand nombre d’agriculteurs produisaient individuellement de plus petites quantités. Actuellement, la tendance est à l’inverse comme le montre le tableau 2.2.
6
Tableau 2.2: Nombre de fermes et superficies ensemencées au Canada et dans différentes provinces en 1971 et 2011 (Statistique Canada, 2011)
Le tableau 2.2 met en évidence le phénomène d’intensification agricole par l’augmentation marquée du nombre d’hectares ensemencés par ferme. Par exemple, le nombre d’hectares ensemencés au Québec a passé de 55 817 ha en 1971 à 402 441 ha en 2011. Cela représente en termes de superficie moyenne par ferme 13,5 ha en 1971 et 65,3 ha en 2011. Les superficies ensemencées par ferme sont donc environ cinq fois plus grandes en 2011 qu’en 1971. Les méthodes culturales ont donc évolué pour permettre un meilleur rendement par hectare. Parmi ces méthodes, il y a la rotation des cultures qui permet au sol de se régénérer. En 2011, 95% des producteurs de maïs-grain ont pratiqué la rotation des cultures suivantes : maïs-maïs-grain-soya-blé (le plus populaire), maïs-grain-soya (le deuxième) et maïs-grain-foin-soya-blé (le troisième) (Hamel et al., 2014). Cette rotation, en particulier avec les légumineuses, permet la fixation de l’azote dans le sol, évite l’érosion et rompt le cycle de propagation des maladies et des ravageurs.
2.2 Pyrale du maïs (Ostrinia nubilalis)
La pyrale du maïs, Ostrinia nubilalis (Hübner), est originaire d’Europe et a été introduite dans la région des Grands Lacs vers 1900 (Hagerman, 1997). Elle s’est ensuite répandue dans l’est et le centre de l’Amérique du Nord. La pyrale du maïs fait partie de l’ordre des Lépidoptères et de la famille des Crambidés. L’adulte a une longueur d’environ 12 mm. La figure 2.3 montre la pyrale adulte.
7
Figure 2.3 : Pyrale du maïs mâle à gauche et femelle à droite au stade adulte (Ferland et al., 2000).
2.2.1 Stades de développement, dommages causés et échantillonnages de la pyrale du maïs
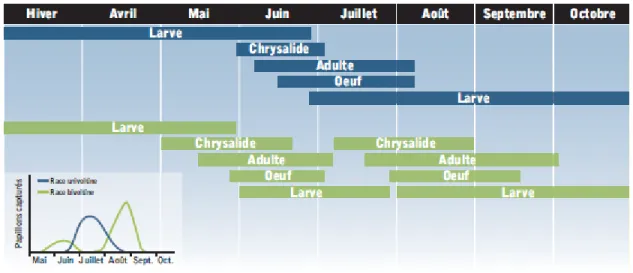
La pyrale possède quatre stades de développement en passant d’œuf en larve puis chrysalide et finalement en papillon adulte. La pyrale du maïs se présente sous deux races, soit la race univoltine et la race bivoltine. L’univoltine a une seule génération par année vers le mois de juillet, soit la mi-saison. La bivoltine a deux générations par année, une au mois de mai et une autre vers le mois d’août, soit en début et fin de saison. Pour l’univoltine, c’est la larve ayant passé l’hiver qui se développe durant l’été. Pour la race bivoltine, la larve ayant survécu à l’hiver débute sa croissance plus tôt en saison que l’univoltine. Tôt au printemps, la larve se transforme en chrysalide et ensuite en papillon entre la fin du mois de mai et la fin-juin, selon la disposition géographique. Le papillon adulte mesure de 1,5 à 2 cm de longueur et 1 cm de largeur les ailes repliées (Hagermann, 1997). La femelle adulte nouvellement formée pond des œufs qui seront à la base de la deuxième génération de la saison. La figure 2.4 présente les différents stades de développement des deux générations de pyrale du maïs pour une saison.
8
Figure 2.4 : Stades de développement en fonction du temps pour les races univoltine et bivoltine de la pyrale du maïs (Boisclair et Jean, 2009).
La pyrale du maïs est le principal ravageur du maïs au Québec. Les larves de la pyrale du maïs s’attaquent d’abords aux feuilles des plants de maïs, puis creusent des galeries dans les tiges et les épis. La verse des plants peut ainsi survenir et rendre la récolte difficile menant à un rendement moindre. Les attaques des épis rendent le produit peu attrayant pour les clients en plus de causer des maladies. Pour le producteur, les pertes financières peuvent être importantes.
La pyrale du maïs s’attaque non seulement au maïs mais aussi aux poivrons et aux pommes de terre. Ces trois cultures sont importantes pour la province du Québec. Pour contrôler les ravages causés par cet insecte nuisible, il convient de surveiller sa croissance. Plusieurs méthodes sont utilisées. Lors de la croissance du maïs, les feuilles sont examinées pour vérifier la présence d’ooplaques, soit une masse d’œufs. La femelle pond des masses de 10 à 40 œufs. Les masses d’œufs sont blanches et ont seulement quelques millimètres de longueur (Hagermann, 1997). Les ooplaques sont souvent disposées sous les feuilles de maïs près de la nervure. La femelle peut pondre deux ooplaques par jour sur une période de dix jours. L’éclosion des œufs prend de trois à neufs jours selon la température. Lorsque les œufs présentent une tête noire, l’éclosion se fera dans les prochains jours comme le montre la figure 2.5.
9
Figure 2.5: Œufs de pyrale du maïs et jeunes larves (Brodeur, 2008).
La présence de larves dans les tiges ou dans les épis peut aussi être vérifiée en les coupants comme le montre la figure 2.6.
Figure 2.6 : Larves de pyrale ayant creusées dans une tige et dans des épis de maïs (Hagerman, 1997).
Finalement, les papillons peuvent être capturés par des pièges à phéromones ou lumineux. Les pièges à phéromone possèdent des capsules contenant des phéromones sexuelles spécifiques à la pyrale du maïs et une surface engluée pour piéger les papillons. Les pièges sont vérifiés deux à trois fois par semaine et les papillons capturés sont comptés. Il a été constaté qu’au Québec, peu de pyrales adultes (papillon) de race bivoltine sont capturées au printemps. Cela s’explique par le cycle biologique de la pyrale, car ces larves sont issues de la seconde génération de bivoltine née tardivement à la fin de l’été précédent. Elles sont donc plus fragiles et survivent moins bien aux premières gelées à l’automne. Cela se traduit par une faible population de bivoltine au printemps suivant (Caron et al., 1995). Ces différentes méthodes de dépistage peuvent être faites soit par un agronome, une firme spécialisée en dépistage de phytophages ou par l’agriculteur lui-même. Le réseau d’avertissement phytosanitaire (RAP) permet aussi de connaître les régions et la date où la pyrale sera en éclosion au Québec. Le RAP
10
documente les captures dans plusieurs régions et permet de constater en temps réel l’émergence des phytophage. Il émet des rapports sous forme d’avertissement pour informer les utilisateurs des dates approximatives d’utilisation de pesticides ou de trichogrammes.
2.3 Pesticides
Les pesticides chimiques sont les plus utilisés pour contrôler la pyrale du maïs. Au Québec, plusieurs classes de composés chimiques ont été homologuées en 2015 pour la lutte contre la pyrale du maïs dans la culture du maïs sucré. Plusieurs composés appartenant aux carbamates, aux spinosynes, aux pyréthroïdes synthétiques, aux organophosphatés, aux diacylhydrazine ou aux diamines ont pu être utilisés aux champs (RAP, 2015).
2.3.1 Évaluation du danger des pesticides
L’utilisation des pesticides chimiques n’est pas sans danger. En effet, près de 10% des intoxications rapportées annuellement au Centre antipoison du Québec concernent des utilisateurs professionnels de pesticides (Samuel et St-Laurent, 2014). La contamination par les pesticides se fait surtout par contact cutané mais parfois aussi par voies respiratoires par aspiration d’aérosol, de brouillard ou de gaz. Pour se protéger des pesticides, l’utilisateur doit donc porter un équipement adéquat de protection individuelle et respecter scrupuleusement les consignes d’utilisation du produit. Chaque pesticide comporte des risques différents. C’est pourquoi le Québec s’est doté d’un outil de caractérisation des dangers des pesticides nommé indicateur de risque des pesticides
du Québec (IRPeQ). La stratégie phytosanitaire québécoise en agriculture (SPQA)
2011-2021 s’est donnée comme objectif de réduire de 25% les indicateurs de risque pour la santé et l’environnement d’ici 2021 par rapport à la moyenne des années 2006, 2007 et 2008. Deux unités de mesure sont utilisées par l’IRPeQ pour évaluer les risques reliés à l’utilisation des pesticides, soit l’indice de risque pour la santé (IRS) et l’indice de
risque pour l’environnement (IRE).
2.3.2 Indice de risque pour la santé
L’indice de risque pour la santé prend en considération la toxicité aiguë (effets immédiats), la toxicité chronique (effets à long terme), l’effet du type de formulation ainsi que la méthode d’application. Il est la combinaison de différents points accordés aux
11
facteurs de danger reliés à l’utilisation d’une matière active (Samuel et al., 2012). L’équation 2.1 montre l’indice de risque toxicologique.
∗ 2.1
où FPer est le facteur de persistance et le potentiel de bioaccumulation chez l’humain. L’équation 2.1 est mise au carré pour augmenter la pondération des matières présentant le plus de risques. Cette équation est utilisée dans le calcul de l’IRS pondéré qui est l’indice de risque pour la santé pondéré d’une matière active. L’équation 2.2 détaille ce calcul.
è ∗ ∗
10 2.2
où FPf est le facteur de pondération tenant compte de la formulation et FCP est le facteur de pondération pour la concentration de la matière active dans la préparation commerciale et dans la dose appliquée. L’IRS de la matière active pondéréeest divisée par 10 pour obtenir un ordre de grandeur acceptable. Comme une préparation commerciale peut contenir plusieurs matières actives, l’IRS de la préparation commerciale sera la somme des IRS des matières activescomme le montre l’équation 2.3.
é è 2.3
Cette approche conservatrice permet de ne pas sous-estimer un effet propre à un composé. Dans le but de relier l’IRS à l’utilisation d’un pesticide dans un champ, l’IRS ajustépermet de tenir compte de la technique et du lieu de pulvérisation. L’équation 2.4 montre ce calcul.
é é ∗ 2.4
où FPa est le facteur de pondération relié à l’application. Finalement, l’IRS traitementqui est le risque pour la santé relié à un traitement est la somme des IRS de toutes les préparations commerciales utilisées pendant le traitement comme le montre l’équation 2.5.
12 2.3.3 Indice de risque pour l’environnement
L’indice de risque pour l’environnement tient compte de plusieurs facteurs comme la toxicité pour les espèces non visées et le comportement à long terme dans la nature (bioaccumulation, persistance et mobilité). Il est calculé en tenant compte de l’impact écotoxicologique et physicochimique des pesticides sur l’environnement. L’équation 2.6 permet de constater le poids de chacun de ces paramètres.
è 1,75 ∗ 1 2.6
où les paramètres écotoxicologiques sont représentés par les impacts sur les invertébrés terrestres (T), les oiseaux (O) et les organismes aquatiques (A). Les paramètres physicochimiques quant à eux sont représentés par la mobilité (M), la persistance dans le sol (P) et la bioaccumulation (B). Tous ces paramètres sont déterminés par des équations empiriques détaillées dans l’ouvrage Indicateur de risques
des pesticides du Québec (Samuel et al., 2012). Ils sont une adaptation des indicateurs
de risques utilisés en Norvège. Tout comme pour l’IRS, l’IRE pour une préparation commerciale est la somme des IRE des matières actives utilisées. Finalement, l’IRE de traitement est la somme de tous les IRE des préparations commerciales utilisées lors de ce traitement.
2.3.4 Résistance aux pesticides
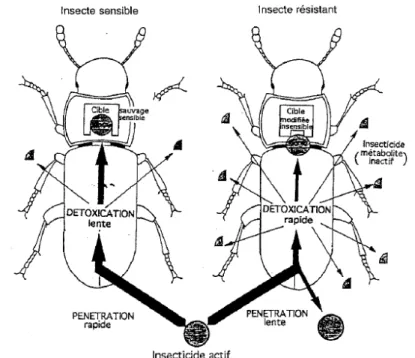
L’utilisation de pesticides peut engendrer à long terme un phénomène de résistance du ravageur. Par le passé, cette résistance était contrée par l’augmentation des doses de pesticides ou par l’utilisation de nouveaux composés. Heureusement, ces méthodes ne sont plus utilisées aujourd’hui à la lumière du danger et des coûts reliés à leur utilisation. La résistance aux pesticides peut se présenter sous trois formes, soit la résistance comportementale, physiologique et biochimique (Haubruge et Amichot, 1998). La figure 2.7 montre ces trois types de résistance.
13
Figure 2.7: Représentation schématique des différentes résistances aux pesticides chez un insecte (Haubruge et Amichot, 1998).
Deux types de résistance comportementale peuvent avoir lieu. Le premier est la résistance liée à la mobilité de l’insecte qui peut détecter les molécules toxiques et éviter les zones où elles sont présentes ou diminuer grandement le temps de contact avec celles-ci. Le second type de résistance comportementale est associé à l’immobilité de l’insecte qui peut limiter ses déplacements sur une surface où il y a des pesticides. Par exemple, le comportement des larves résistantes aux pyréthrinoïdes de l’espèce
Héliothis virescens fut étudié en laboratoire en comparaison à des individus sensibles à
ces pesticides (Sparks et al., 1989). Le résultat fut que les larves résistantes étaient en contact avec une quantité de pesticides beaucoup moins grande que les larves sensibles. En limitant leur mouvement, les larves résistantes viennent en contact avec moins de composés toxiques et leurs chances de survie en sont ainsi augmentées. La résistance physiologique tient compte de la cinétique d’absorption des composés à travers la cuticule ou la paroi digestive, leur séquestration ou leur excrétion. La vitesse d’absorption est différente pour chaque espèce et pour chaque produit. Si elle est suffisamment lente, la molécule pourra être dégradée par le système de détoxification. La concentration en produits toxiques restera donc faible et peu d’effets seront ressentis. La population d’une espèce possédant cette cinétique d’absorption sera favorisée au détriment des autres absorbants plus spécifiquement les pesticides. La
14
population sera de plus en plus constituée d’individus résistants et le pesticide ne sera plus aussi efficace. Certaines espèces peuvent aussi excréter les composés toxiques sans biotransformation (Sparks et al., 1989).
La résistance biochimique se passe au niveau cellulaire. Les composés pénètrent plus ou moins rapidement les protéines et les enzymes des cellules et modifient leur comportement. Deux types de modification biochimiques peuvent avoir lieu, soit une augmentation des activités de dégradation des composés étrangers et une modification de la cible de l’insecticide devenant capable de fonctionner malgré la présence de l’insecticide.
De nombreux insecticides étant liposolubles, certaines modifications du métabolisme des insectes résistants permettent de modifier la molécule toxique en ajoutant des fonctions la rendant plus polaire et facilitant par le fait même son excrétion. Le métabolisme peut aussi conjuguer les toxines avec des sucres ou des acides aminés afin de diminuer leurs réactivités.
L’augmentation des activités de dégradation des xénobiotiques (pesticides) peut se faire en une ou deux phases. La phase I consiste en l’oxydation ou l’hydrolyse du composé qui devient plus polaire et peut être excrété ou stocké loin des organes vitaux. Par la suite, les composés de la phase I peuvent être modifiés par la phase II qui consiste en une conjugaison du métabolite primaire avec des sucres ou des acides aminés qui sont fortement hydrophiles. Ils sont donc plus facilement excrétés par l’organisme. La figure 2.8 présente ces différents processus de métabolisation des composés toxiques.
15
Figure 2.8 : Processus biochimique de métabolisation des composés toxiques absorbés par l’insecte (Haubruge et Amichot, 1998).
La modification de la cible de l’insecticide consiste en la présence d’un nouveau gène qui rend la cible insensible à la molécule toxique. Ce gène nommé knockdown
resistance (kdr) a permis à certains insectes de rapidement devenir insensibles aux DDT
et pyréthrinoïdes.
2.4 Génie génétique
La découverte de l’ADN a permis aux scientifiques de modifier en laboratoire le patrimoine génétique des organismes vivants. Le maïs a été génétiquement modifié dans les années 1990 afin d’inclure dans son génome un ou plusieurs gènes de résistance à certains insectes ravageurs ou à des herbicides (Razze et Mason, 2012). Ces organismes génétiquement modifiés permettent d’augmenter le rendement par hectare en diminuant les ravages des insectes.
En Ontario, principal producteur canadien de maïs, 71,3% de la surface totale cultivée en maïs était génétiquement modifié en 2011 (Hamel et al., 2014). À la même année, 73,8% du maïs-grain était génétiquement modifié.
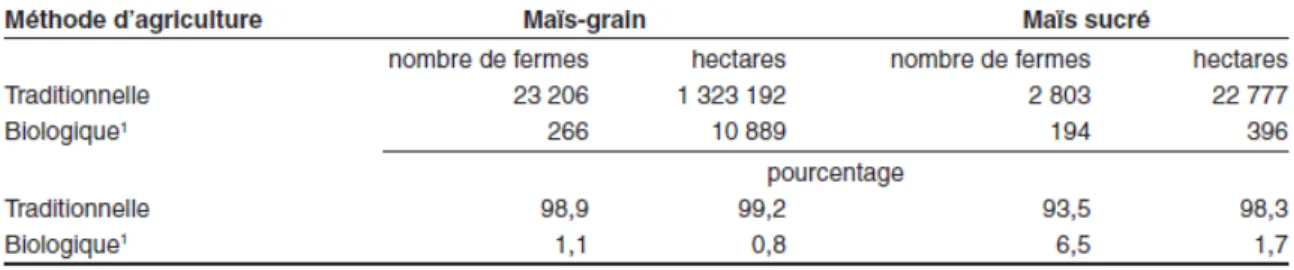
Au Canada, la grande majorité des cultures était conventionnelles, c’est-à-dire en utilisant des semences génétiquement modifiées, des pesticides ou les deux. Seulement 0,8% et 1,7% des superficies cultivées en maïs-grain et en maïs sucré, respectivement, étaient certifiées biologiques ou en voie de le devenir. Probablement que le désir des
16
consommateurs de manger des aliments plus sains forcera l’industrie à offrir plus de produits biologiques à sa clientèle. Le tableau 2.3 présente la part de chaque type de culture du maïs au Canada.
Tableau 2.3: Répartition du maïs –grain et du maïs sucré selon la culture traditionnelle et la certification biologique au Canada en 2011 (Statistique Canada, 2011)
2.4.1 Maïs Bt
Le maïs Bt possède des gènes provenant de la bactérie Bacillus thuringiensis (Bt) codant la production de plusieurs protéines insecticides crystalines (Cry) (Razze et Mason, 2012). Par exemple, le maïs Bt 176 possède un gène codant une protéine toxique à la pyrale du maïs provenant de la bactérie du sol Bacillus thuringiensis. Santé Canada a jugé que ce type de maïs n’est pas toxique et qu’il est aussi nutritif que les variétés conventionnelles (Santé Canada, 1999).
2.4.2 Refuge de maïs non-Bt
Les insectes ravageurs, en contact avec la protéine Bt, peuvent développer une résistance à celle-ci à cause de la variabilité génétique de la population. La présence de cette protéine sélectionne les individus tolérants et avec le temps, met en péril l’efficacité de cette technologie (Razze et Mason, 2012 ; Siegfried et Hellmich, 2012). C’est pour cette raison qu’il est très important de créer des refuges avec des hybrides non-Bt pour permettre le croisement entre individus tolérants et non-tolérants et ainsi maintenir la vulnérabilité des insectes au Bt. Pour s’assurer que les refuges soient créés, l’Agence canadienne d’inspection des aliments (ACIA) oblige les producteurs utilisant du maïs Bt à semer des refuges avec des hybrides non-Bt. Les refuges doivent représenter entre 5 et 20% de la surface totale ensemencée dépendamment du type de Bt utilisé. Un Bt ayant plusieurs modes d’action doit représenter 5% de la surface totale et être à moins de 400 m du champ pour la lutte contre la pyrale du maïs. Dans le cas d’un Bt spécifique à la pyrale, 20% minimum de la superficie du champ doit être non-Bt et se trouver à moins de 400 m. Pour un Bt spécifique à la chrysomèle, un minimum de 20% de la
17
surface totale doit aussi être non-Bt mais ce refuge doit être adjacent ou dans la parcelle Bt pour être efficace. Cette différence de distance du refuge s’explique par la plus grande mobilité de la pyrale en comparaison à la chrysomèle pendant l’accouplement (RAP, 2014).
2.5 Agriculture biologique
L’agriculture biologique est une méthode de production agricole respectueuse de l’environnement et des écosystèmes. Aucun intrant de synthèse n’est utilisé en agriculture biologique comme les pesticides de synthèse, les fertilisants chimiques, les hormones de croissance et les antibiotiques (Office des normes générales du Canada, 2011).
2.5.1 Marché biologique
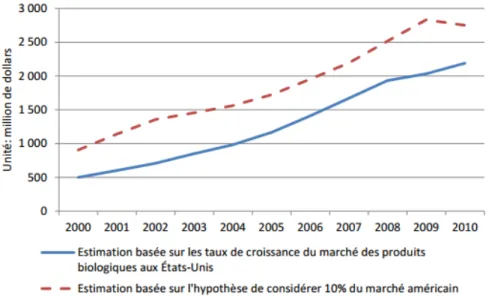
La demande pour les produits issus de l’agriculture biologique ne cesse d’augmenter, car la population est plus sensibilisée aux enjeux environnementaux et sociaux. Les problèmes de pollution de l’eau par les pesticides sont bien connus et les effets de ceux-ci sur la santé des humains et des animaux sont de plus en plus préoccupants. Entre 2000 et 2009, les ventes dans le monde de produits biologiques au monde ont plus que triplé (Willer et Kilcher, 2011). En 2010, le marché américain représentait environ la moitié du marché mondial, soit 26,7 milliards de dollars. Pour le Canada, il n’existe pas d’estimation précise des ventes de produits biologiques. Les données américaines ont donc été utilisées pour estimer l’évolution du marché canadien. Deux méthodes ont été utilisées, soit l’estimation basée sur les taux de croissance du marché des produits biologiques et l’estimation basée sur l’hypothèse de considérer 10% du marché américain (la population du Canada est environ le dixième de celle des États-Unis). La figure 2.9 présente l’évolution du marché des produits biologiques au Canada.
18
Figure 2.9: Estimations de l’évolution du marché total des produits biologiques au Canada de 2000 à 2010 (ÉcoRessources Consultants, 2012).
La figure 2.9 montre une croissance marquée du marché biologique dans la dernière décennie. Cependant, la crise économique de 2008 a ralenti quelque peu l’évolution du marché. Comme le marché des aliments biologiques est restrictif, il nécessite certains incitatifs car les coûts de production sont plus élevés et le marché est assez limité. Les gouvernements canadien et québécois ont donc créé des programmes pour aider financièrement les producteurs. Une certification biologique a aussi été créée pour permettre au consommateur de connaître l’authenticité des produits et rassurer les producteurs face à la conservation de leurs parts de marché. Une part grandissante de l’utilisation canadienne des fruits et légumes biologiques sert à produire des jus et des soupes ou pour la congélation. En effet, les ventes de légumes biologiques congelés ont passé de 900 000 dollars en 2006 à 7,2 millions de dollars en 2008 (AC Nielsen Canada, 2009). De plus en plus de fermes québécoises se convertissent en agriculture biologique.
Entre 2006 et 2013, le nombre de fermes au Canada ayant obtenu la certification biologique est passé de 855 à 1003 soit une augmentation de 15%. Cependant, le marché biologique reste assez marginal avec seulement 2 à 3% de la production alimentaire canadienne. De plus, plus de 70% des produits biologiques consommés au Québec sont importés (Fédération d’agriculture biologique du Québec, 2014). Le marché biologique québécois doit donc augmenter son volume de production et parfaire sa publicité pour mieux répondre à la demande des consommateurs.
19
2.6 Lutte biologique aux phytophages
La lutte biologique aux phytophages consiste à utiliser des organismes prédateurs, parasitoïdes, microorganismes entomopathogènes ou des toxines naturelles pour contrôler une population de ravageurs. En présence d’insectes ravageurs dans un champ de maïs par exemple, l’utilisation de parasitoïde comme le trichogramme permet de diminuer le nombre de ces ravageurs à un niveau acceptable. Comme une très grande quantité de nourriture est disponible au champ, les phytophages ont tendance à se multiplier rapidement. L’introduction d’insectes prédateurs permet ainsi de rétablir l’équilibre biologique des parcelles cultivées.
La lutte biologique aux insectes ravageurs ne date pas d’hier. En effet, les chinois utilisaient la fourmi Pharaon Monomorium pharaonis pour combattre les insectes s’attaquant aux réserves de grains. Cette pratique a été utilisée aussi tôt que 324 av. J.-C. (Coulson et al., 1982). Plus récemment en 1887-88, la coccinelle Rodolia cardinalis fut utilisée pour la première fois dans les vergers d’agrumes contre la cochenille australienne Icerya purchasi. Ce coléoptère fut trouvé dans l’habitat naturel de la cochenille en Océanie et il est encore en action dans les vergers aux États-Unis. Cette utilisation d’un prédateur contre un insecte ravageur est un bon exemple d’une méthode durable, simple et peu coûteuse de gestion des risques phytosanitaires.
2.6.1 Biopesticides
Les biopesticides sont des composés insecticides d’origine naturelle. Avant la seconde guerre mondiale, les chercheurs en agriculture se sont beaucoup intéressés aux produits naturels biologiquement actifs provenant des plantes. Cependant, la croissance industrielle dans la seconde moitié du XXe siècle provoqua le développement de composés chimiques de synthèses utilisés massivement au champ (Regnault-Roger et al., 2008). Les effets négatifs de cette utilisation intensive menèrent à la pollution des cours d’eau et à la mise en péril de certaines espèces animales. Depuis quelques décennies, un regain d’intérêt pour les biopesticides d’origine végétale a eu lieu partout dans le monde. Cet intérêt est appuyé par le fait que la plupart des biopesticides possèdent une sélectivité remarquable envers leur cible, une activité spécifique, et sont généralement biodégradables. Ces caractéristiques permettent une utilisation très ciblée contre un insecte en particulier en évitant de déranger la faune avoisinante.
20
De plus et à l’aide des outils analytiques modernes, les chercheurs sont en mesure de mieux comprendre les relations structure-activité des composés pour pouvoir identifier de nouveaux composés avec une forte activité biologique. Au Québec, les ventes de biopesticides ont passées de 9 141 kg en 1992 à 100 883 kg en 2010. Ces ventes en 2010 représentent 2,6% des ventes totales de pesticides (Gorse et Balg, 2013).
2.6.2 Parasitoïdes
Les parasitoïdes sont caractérisés par le mode d’alimentation de leurs larves qui se nourrissent exclusivement du corps de leur hôte et qui éventuellement le tuent, à la différence d’un parasite (Godfray, 1994). Contrairement à un prédateur qui peut consommer plusieurs proies pour compléter son développement, un parasitoïde se nourrit d’un seul hôte pour achever sa croissance. De plus, un parasitoïde parasite son hôte durant son développement avant de le tuer puis vit librement au stade adulte. Le stade de développement des parasitoïdes comporte quatre phases, soit l’œuf, la larve, la pupe et l’adulte. Les parasitoïdes peuvent êtres solitaires ou grégaires selon le nombre d’individus se nourrissant d’un même hôte. Les parasitoïdes koinobionte vivent avec leur hôte vivant pendant un certain temps après l’oviposition avant de le tuer, ce qui permet à l’hôte de continuer à se nourrir et à alimenter les parasitoïdes en développement. À l’inverse, les idiobiontes sont des parasitoïdes qui tuent leur hôte rapidement après l’oviposition, ce qui limite les nutriments disponibles à ceux présents au départ. Ils peuvent être endoparasitoïdes ou ectoparasitoïdes selon le lieu de développement, soit dans l’hôte ou à la surface de l’hôte, respectivement. Généralement, les parasitoïdes koibiontes sont des endoparasitoïdes et sont spécifiques à leur hôte à cause de l’adaptation nécessaire pour pouvoir échapper aux mécanismes de défense de celui-ci (Hajek, 2004). À l’inverse, les idiobiontes sont souvent des ectoparasitoïdes et sont plus spécifiques à un habitat qu’à un hôte en particulier. Les parasitoïdes peuvent aussi être classés selon le stade de croissance où l’hôte sera attaqué. Des parasitoïdes d’œufs, de larves, de pupes et d’adultes existent ainsi que des parasitoïdes d’œufs-larves et larves-pupes. Ces derniers sont des koinobionte laissant leur hôte vivant suffisamment longtemps pour le laisser changer de phase de développement. Parfois, plusieurs ovipositions de femelles de la même espèce ont lieu dans un seul hôte ce qui mène au superparasitisme. Le superparasitisme est néfaste au développement des larves en augmentant la compétition pour les nutriments présents dans l’hôte. Le multiparasitisme est semblable, mais implique un seul hôte attaqué par plusieurs femelles de différentes espèces. L’hyperparasitisme implique la présence
21
d’une seconde espèce de parasitoïde se nourrissant de la larve du premier parasitoïde. Les hyperparasitoïdes facultatifs peuvent se développer seuls dans un hôte ou dans un parasitoïde présent dans l’hôte s’il est déjà parasité lors de l’oviposition. Les hyperparasitoïdes obligatoires ne peuvent pas se développer seuls et doivent parasiter une larve de parasitoïde déjà présente dans un hôte (Godfray, 1994).
2.6.3 Trichogrammes
Les trichogrammes sont des Hyménoptères parasitoïdes oophages appartenant à la famille des Chalcidides. Il existe plus de 600 espèces réparties sous 80 genres (Boisclair et Jean, 2009). Un parasitoïde oophage pond ses œufs dans ceux de son hôte qui sert de milieu de croissance et comme source de nourriture pour les larves. Contrairement à un parasite, un parasitoïde tue sa proie à un moment ou à un autre pour s’en nourrir. Un trichogramme ressemble à une petite guêpe de moins d’un millimètre de long. La figure 2.10 montre son action sur des œufs de pyrale du maïs.
Figure 2.10 : Femelle Trichogramma ostriniae parasitant des œufs de la pyrale du maïs (Marchal, 1936).
Le cycle de vie des trichogrammes débute par l’éclosion de l’un des œufs parasités l’été précédent sur l’ooplaque. Les œufs en diapause pondus par les trichogrammes sont généralement ceux du centre du groupe d’œufs comme ceux en noir à la figure 2.10. Suite à son éclosion, les femelles cherchent des œufs à parasiter. Souvent, les œufs en périphérie de l’ooplaque sont libres et utilisés par les nouvelles femelles comme le montre la figure 2.10. Une femelle T. brassicae par exemple peut parasiter entre 80 et 110 œufs hôtes au cours de sa vie. Si la femelle n’a pas été fécondée, elle pondra des œufs d’où émergeront des mâles alors que fécondée, ses progénitures seront des deux sexes (Hawlitzky, 1992).
22
Suite à la ponte d’un œuf dans l’hôte, celui-ci est tué peu de temps après et les tissus désintégrés servent de nutriments à la larve du trichogramme. Douze heures après l’injection de l’œuf, l’embryon hôte est en voie de dégradation probablement à cause de substances introduites par la femelle trichogramme. Durant sa vie larvaire, le trichogramme consommera la totalité de l’hôte.
Les œufs parasités deviennent complètement noirs après quatre à cinq jours. Ils se transforment ensuite en nymphes puis en adultes avant de percer l’enveloppe de l’insecte hôte après une dizaine de jours (Jean, 2008). L’adulte est prêt pour l’accouplement et la ponte vers d’autres hôtes. L’adulte se nourrit de la substance sucrée produite par les pucerons (miellat) ou de substances protéiques (pollen).
Les trichogrammes sont des insectes haplodiploïdes, ce qui signifie que les œufs fécondés donneront des femelles alors que les non-fécondés donneront des mâles. La femelle trichogramme pondant les œufs choisit lesquels seront fécondés, phénomène visible par la présence d’une pause lors des contractions. Fait intéressant, la température influence beaucoup la détermination du sexe du trichogramme (Moiroux et al., 2014). Lorsqu’il fait chaud (34oC), la femelle choisit d’engendrer plus de mâles. À l’inverse, une température plus fraiche (14oC) fera en sorte que la femelle choisira de créer autant de femelles qu’à température moyenne, mais un problème de fécondité occasionnera la présence de plus de mâles. Ainsi, un facteur physiologique empêche la femelle de féconder convenablement ses œufs à basse température.
2.7
Utilisation
des trichogrammes pour la lutte contre la pyrale du
maïs
Les trichogrammes sont utilisés depuis les années 1970 dans la lutte contre les insectes ravageurs des cultures. Les trichogrammes étant déjà présents dans l’environnement, le but du traitement consiste à augmenter considérablement leur présence au champ. Ils ne présentent aucun danger pour la flore, les animaux et les humains.
Cet insecte parasitoïde est très étudié par les scientifiques pour mieux comprendre ses caractéristiques et son mode de reproduction. Depuis 1982, un groupe de travail dédié à ce parasitoïde a été créé au sein de l’Organisme international de lutte biologique (OILB) ainsi qu’un journal de liaison annuel nommé Trichogramma News. Des recherches pour trouver les souches les mieux adaptées aux différents milieux de culture sont
23
couramment effectuées. Par exemple, une étude réalisée par Wajnberg et Colazza (1998) se questionna sur l’effet de la vitesse de marche de différentes souches de
Trichogramma brassicae sur le taux de parasitisme. En filmant les femelles marchant et
en analysant les paramètres de déplacement, les chercheurs ont conclu que certaines populations de T. brassicae, en provenance de pays plus chauds, se déplaçaient plus vite et plus efficacement. Ce comportement serait à l’origine d’une plus grande capacité à parasiter les œufs hôtes.
Dans le but de contrôler la production des trichogrammes, les potentialités des individus doivent être étudiées. Des paramètres comme la fécondité, la longévité, le taux de parasitisme, le taux d’émergence et les proportions de mâles et de femelles permettent d’obtenir des rendements de production intéressants. L’état de l’hôte est aussi important pour obtenir un taux d’émergence optimal.
Les trichogrammes sont utilisés au Québec depuis plus de 15 ans pour lutter contre la pyrale du maïs. Au moins 15% des producteurs de maïs sucré de la province utilisent cette méthode de lutte biologique dans leurs champs (Anatis Bioprotection, 2013). Cela correspond à environ 1500 hectares protégés par les trichogrammes.
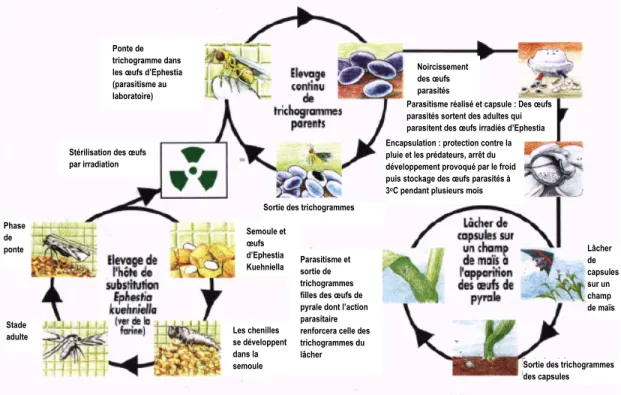
En Europe, les études ont été principalement faites sur Trichogramma brassicae. Pour une utilisation au Québec, Trichogramma ostriniae a été l’espèce privilégiée suite à des essais sur terrain et en laboratoire (Boisclair et al., 2011). Les auteurs ont étudiés les deux espèces sous les mêmes conditions et ils ont constaté que Trichogramma ostriniae présentait de meilleurs résultats que Trichogramma brassicae en termes de suivi et de capacité de parasitisme. Les trichogrammes sont présentement élevés dans des installations contrôlées avant que leurs œufs soient expédiés aux agriculteurs. Les œufs hôtes où les trichogrammes pondent leurs œufs sont préalablement stérilisés avant d’être parasités et proviennent de la pyrale de la farine Ephestia kuehniella. La figure 2.11 montre le processus de production des trichogrammes.
24
Figure 2.11 : Cycle de production et d’utilisation des trichogrammes dans la lutte biologique contre la pyrale du maïs (Hawlitzky, 1992).
Dans cette figure, on distingue trois cycles distincts lors de la production des trichogrammes. Le premier cycle consiste à produire des œufs hôtes pouvant être parasités. L’hôte de substitution utilisé est élevé dans un milieu de développement riche en nutriments pour favoriser sa croissance. Une fois les œufs hôtes prêts, ils sont stérilisés par radiation pour tuer l’œuf sans modifier sa composition. Le second cycle peut débuter en mettant en contact les œufs stérilisés avec les femelles trichogrammes. Celles-ci peuvent parasiter les œufs qui noircissent. Une partie est conservée pour compléter le cycle alors que la majorité est encapsulée ou disposée sur des trichocartes puis mise au froid en attendant le moment propice à leur utilisation. Cette mise au froid engendre la diapause chez le trichogramme qui est un animal poïkotherme ou à sang-froid. En refroidissant le milieu où ils sont, leur métabolisme est suffisamment ralenti pour conserver les œufs parasités pendant deux mois maximum.
2.7.1 Degré-jour
Le degré-jour ou DJ est une unité empirique utilisée pour déterminer la chronologie de croissance des poïkothermes. La vitesse de croissance des poïkothermes est très dépendante de la température tout comme la cinétique des réactions chimiques.
Ponte de trichogramme dans les œufs d’Ephestia (parasitisme au laboratoire)
Stérilisation des œufs par irradiation Phase de ponte Semoule et œufs d’Ephestia Kuehniella Les chenilles se développent dans la semoule Parasitisme et sortie de trichogrammes filles des œufs de pyrale dont l’action parasitaire renforcera celle des trichogrammes du lâcher Sortie des trichogrammes
Noircissement des œufs parasités
Parasitisme réalisé et capsule : Des œufs parasités sortent des adultes qui parasitent des œufs irradiés d’Ephestia Encapsulation : protection contre la pluie et les prédateurs, arrêt du développement provoqué par le froid puis stockage des œufs parasités à 3oC pendant plusieurs mois
Lâcher de capsules sur un champ de maïs Stade adulte
Sortie des trichogrammes des capsules
25
Le degré-jour se base sur une température seuil (ou de base) où la croissance est nulle. L’équation 2.7 présente les détails du calcul standard du degré-jour.
2 2.7
Les DJ peuvent ensuite être calculés et rapportés au nombre de DJ total nécessaire à la croissance d’un organisme (Plouffe et al., 2011). Pour la pyrale du maïs, le seuil retenu est de 10oC (Hawlitzky, 1992). Ainsi, il est plus facile de surveiller l’évolution de la croissance du ravageur en fonction des conditions climatiques présentes pendant la saison et d’effectuer les lâchers de trichogrammes au moment où les œufs du ravageur seront disponibles.
2.7.2 Trichocartes
L’utilisation des trichogrammes dans la lutte biologique contre la pyrale au Québec se fait à l’aide de trichocartes. Les trichocartes sont des cartons cirés pliés en deux ou trois, où les œufs parasités sont placés à l’intérieur pour les protéger des éléments extérieurs. La figure 2.12 montre une trichocarte.
Figure 2.12 : Une trichocarte contenant des œufs parasités par les trichogrammes sur un plant de maïs (Revue UFA, 2014).
Une trichocarte de l’entreprise Anatis Bioprotection contient 8 000 œufs parasités par des trichogrammes et permet de couvrir une surface de 200 m2. Il faudra alors 50 cartes/hectare pour protéger un champ de maïs, ce qui représente une dose de 400 000 œufs/ha. Le tableau 2.4 permet de comparer les coûts d’utilisation des différentes méthodes de lutte contre la pyrale du maïs durant une saison.
26
Tableau 2.4: Coût saisonnier par hectare pour l’utilisation de différentes méthodes de contrôle de la pyrale du maïs au Québec (Boisclair et al., 2011)
IRE = Indice de risque pour l’environnement. IRS = Indice de risque pour la santé.
1 (x5) = Nombre d’introductions ou d’applications.
2 Les coûts incluent le prix moyen de vente du produit (RAP, 2011) + 4$ (main- d’œuvre)
par introduction de trichogrammes ou + 11,05$/ha (main-d’œuvre et machinerie) par application d’insecticides (CRAAQ, 2010).
3 Le coût saisonnier est basé sur le nombre d’applications minimales de produits insecticides de
synthèse estimés (4) qui seraient requises pour couvrir la période de ponte de la pyrale du maïs. Ce nombre peut varier selon la durée et le niveau d’infestation de chaque génération de la pyrale du maïs. Le prix indiqué est le prix pour la dose moyenne recommandée.
27
Dans le tableau 2.4, on constate que le coût saisonnier d’utilisation de T. brassicae est supérieure à celui des autres stratégies de lutte contre la pyrale (430 $/ha). Le tableau 2.4 présente les coûts d’utilisation de T. ostriniae pour une dose de 150 000 pupes/ha Par contre, l’étude montre qu’il faut plutôt utiliser T. ostriniae à une dose de 400 000 pupes/ha pour obtenir une efficacité optimale. Ainsi, le coût saisonnier d’utilisation de T.
ostriniae est supérieur au coût des pesticides conventionnels mais avec un risque pour
l’environnement et pour la santé beaucoup moindre, voir même nul. Somme toute, l’utilisation des trichogrammes pour la lutte biologique contre la pyrale est assez coûteuse à cause des efforts requis pour élever, nourrir, manipuler, conditionner et transporter les trichogrammes qui sont des organismes vivants fragiles. L’utilisation des trichocartes par les agriculteurs est aussi plus difficile que celle d’un pesticide. En effet, les trichocartes doivent être disposées selon un patron précis et installées au bon moment. Cela demande plus d’efforts et de temps qu’une pulvérisation d’un produit chimique liquide.
Une expérience a été menée sur une ferme par le MAPAQ pour déterminer les coûts d’utilisation des trichocartes lors d’une forte présence de la pyrale du maïs (Vinet et al., 2013). L’étude portait sur un champ de 4,07 ha divisé en sept parcelles avec une utilisation de 75 cartes/ha alors que la dose recommandée est de 49 cartes/ha. Le coût d’utilisation obtenu était de 766,14 $/ha, soit 0,25 $/dz pour un rendement de 3 000 dz/ha. Ce coût a été comparé à celui de l’utilisation de l’insecticide Coragen avec quatre applications qui s’élevaient à 336,92 $/ha, soit 0,11 $/dz. Une différence de 0,14 $/dz est donc présente entre les deux méthodes. Cependant, comme les épis de maïs cultivés avec l’utilisation des trichogrammes se démarquaient par leur production sans pesticides, il pouvait être vendu plus cher.
2.8 Distribution sèche
Plusieurs essais ont eu lieu pour mécaniser la distribution d’insectes bénéfiques. Gardner et Giles (1997) réussirent à disperser efficacement des œufs de chrysopes vertes pour lutter contre des insectes ravageurs. Ils ont mélangé les œufs avec de la vermiculite et les ont placés dans un distributeur rotatif équipé d’un jet d’air. Les vibrations et la distribution n’ont pas eu d’effet négatif sur le taux d’éclosion en comparaison à un témoin. Plus récemment, Khelifi et al. (2011) ont réussi à distribuer des nymphes de Podisus maculiventris qui sont des prédateurs du doryphore de la