Role of local flow conditions in river biofilm colonization and early growth
19
0
0
Texte intégral
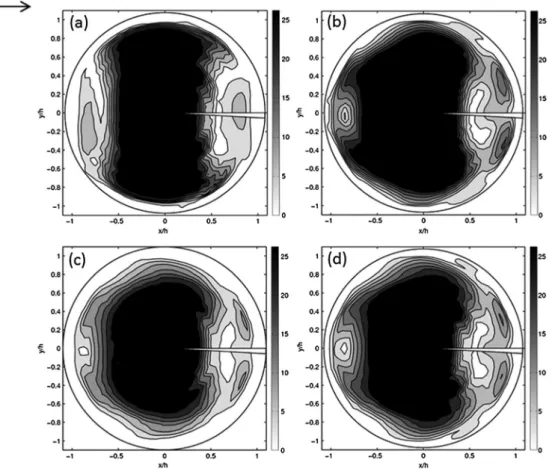
Figure

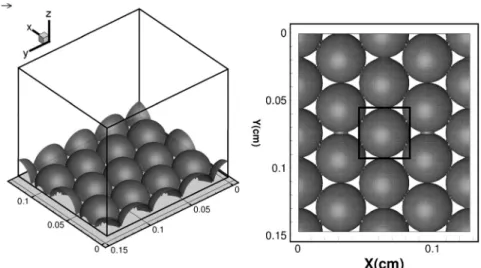
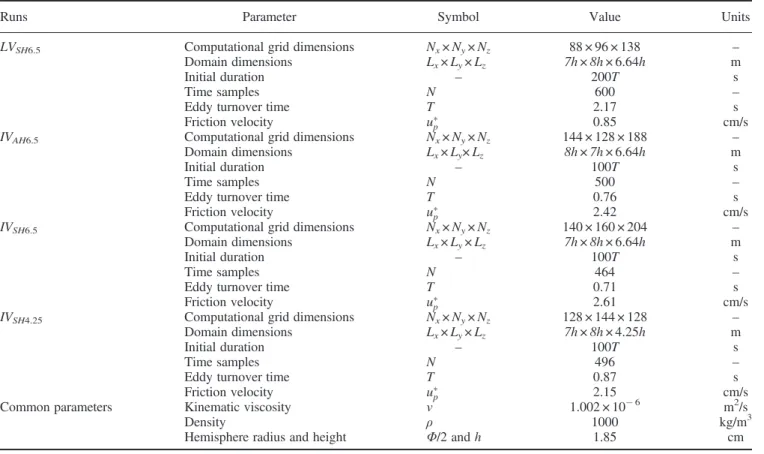
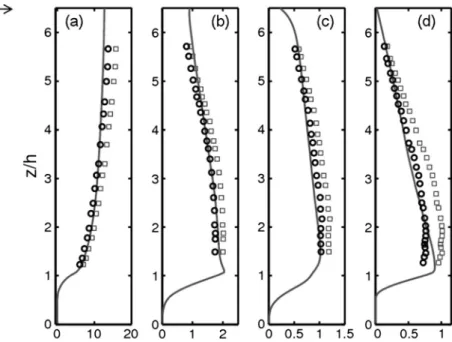
+7


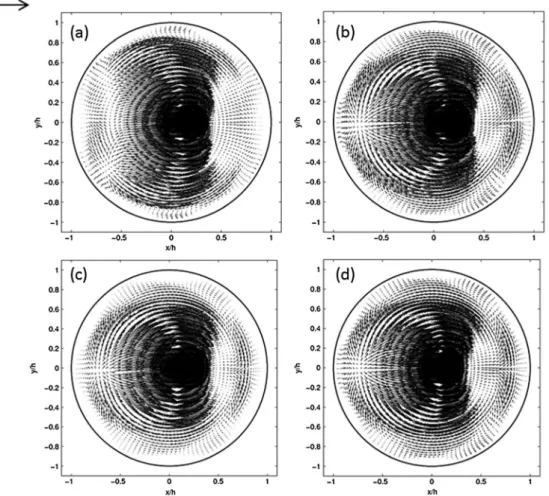


Documents relatifs
