Mémoire
Mays Merjaneh
Maîtrise en biologie cellulaire et moléculaire Maître ès sciences (M. Sc.)
Québec, Canada
Évaluation du rôle des microvésicules de myofibroblastes de plaie dans l’angiogénèse
Mémoire
Mays Merjaneh
Sous la direction de :
Le processus de cicatrisation est complexe et hautement régulé par de nombreux facteurs. La différenciation des fibroblastes (Fb) du derme en myofibroblastes (Wmyo) est très importante pour la guérison des plaies. Les Wmyo accélérent la fermeture des plaies par leur contraction, et jouent un rôle central dans la reconstitution de la MEC. Notre équipe a déjà montré que les Wmyo stimulent l’angiogénèse de façon plus importante que les Fb. De plus, les Wmyo produisent des microvésicules (MV) : des vésicules produites par bourgeonnement de la membrane plasmique des cellules qui jouent un rôle dans la communication intercellulaire. En analysant le protéome de ces MV, l’équipe a démontré que les MV contiennent des protéines impliquées dans l’angiogénèse.
Mon hypothèse est que les MV produites par les Wmyo pouvaient influencer l’angiogenèse. Mes objectifs étaient d’évaluer le rôle des MV produites par les Wmyo sur l’angiogenèse en évaluant la réponse des cellules endothéliales (CE) selon les trois mécanismes fondamentaux de la formation des capillaires sanguins : la prolifération et la migration des CE, ainsi que la formation de structures tubulaires. Pour finir, les mécanismes d’interaction et d’action des MV sur les CE cibles ont été étudiés.
Mes résultats ont prouvé que la présence de MV isolées à partir de Wmyo induit une augmentation significative et dose-dépendante de la prolifération et de la migration des cellules endothéliales microvasculaires de peau (CEMV) tout en favorisant la formation de structures tubulaires sur Matrigel®. Finalement, l’internalisation des MV dans les CEMV par la voie de l’endocytose est probable, et l’action des MV sur la migration des CE semble être dépendante d’une traduction protéique.
En conclusion, nous avons démontré que les MV produites par les Wmyo sont favorables au développement angiogénique, élargissant le rôle des Wmyo pendant la guérison de plaie. Nous avons également prouvé que les mécanismes d’action des MV sont liés à l’endocytose des MV et de la traduction protéique.
The healing process is complex and highly regulated by many factors. The differentiation of dermal fibroblasts (Fb) into myofibroblasts (Wmyo) is very important for wound healing. The Wmyo accelerates wound closure by their contraction. Also, these cells play a central role in the reconstruction of the extracellular matrix. Our team has already shown that Wmyo strongly stimulate angiogenesis, more than Fb and that Wmyo produce microvesicles (MVs). In literature, MVs produced by budding of the plasma membrane of the cells play a role in intercellular communication. By analyzing the proteome of these MVs, I have demonstrated that MVs contain proteins capable of stimulating angiogenesis.
Our hypothesis was that MVs produced by Wmyo could influence angiogenesis.
Our objectives were to evaluate the role of MV produced by Wmyo on angiogenesis by evaluating the response of endothelial cells (EC) according to the three fundamental mechanisms of blood capillary formation: proliferation and migration of endothelial cells, as well as the formation of tubular structures. Finally, we have studied their mechanism of interaction and action on the target EC.
Our results showed that the presence of MVs isolated from Wmyo induces a significant and dose-dependent increase of the proliferation and migration of microvascular endothelial cells of the skin (MVEC) while promoting the formation of tubular structures on Matrigel ®. Also, the internalization of MVs is through endocytosis, and MVs action on EC migration seems to be dependent on protein translation.
In conclusion, we have shown that MVs produced by Wmyo are favorable to an angiogenic development, widening the role of Wmyo during wound healing. We have also shown that the interaction of MVs with EC seems to include the pathway of endocytosis and their role seems to pass via protein translation.
Table des matières
Resumé ... III Summary ... IV Liste des abréviations ... VIII Liste des tableaux ... XI Liste des figures ... XII Remerciement ... XIV
Hypothèse et objectifs ... 1
Chapitre 1-Introduction ... 2
1-1 La peau ... 2
1-1-1 les trois couches de la peau : ... 2
1-1-1-1 L’épiderme ... 2
1-1-1-2 Le derme ... 2
1-1-1-2-1 La matrice extracellulaire ... 3
1-1-1-2-2 Les fibroblastes ... 3
1-1-1-3 L’hypoderme ... 3
1-1-2 Les fonctions de la peau ... 4
1-2 La cicatrisation cutanée... 5
1-2-1 Les phases de la cicatrisation ... 5
1-2-1-1 La phase vasculaire ... 6
1-2-1-2 L’inflammation... 6
1-2-1-3 La prolifération et la migration cellulaire. ... 7
1-2-1-4 le remodelage et maturation ... 8
1-3 L’angiogenèse ... 9
1-3-1 Description de l’angiogenèse ... 9
1-3-2 L’angiogenèse pendant la cicatrisation ... 10
1-3-2-1 La prolifération des cellules endothéliales ... 10
1-3-2-2-la migration des cellules endothéliales ... 11
1-3-2-3-la formation de vaisseaux sanguins et la maturation ... 12
1-4-1 Description des myofibroblastes ... 13
1-4-2 Les myofibroblastes et la cicatrisation ... 14
1-5 Les vésicules extracellulaires (VE) ... 16
1-5-1 Microvésicules et exosomes ... 18
1-5-1-1 Définition des MV ... 18
1-5-1-2 Formation des MV ... 19
1-5-1-4 Implication des MV dans la physiologie et pathologie ... 21
1-5-1-6 Formation des exosomes ... 22
1-5-1-7 La composition des exosomes ... 23
1-5-2 Angiogénèse et microvésicules ... 24
1-5-3 Mode d’action des vésicules extracellulaire ... 25
1-5-3-1 Liaison au récepteur ... 26 1-5-3-2 Internalisation : ... 27 1-5-3-3 Fusion membranaire : ... 30 Chapitre 2- Article ... 33 2-0 Article ... 33 Cell Culture ... 37
Chapitre 3- Résultats complémentaires ... 64
3-0 Résumé ... 64
3-1 Matériel et méthodes ... 65
3-1-1 Transduction des Wmyo ... 65
3-1-2 Isolation des MV fluorescentes ... 65
3-1-3 Quantification des MV isolées ... 65
3-1-4 Test de pénétration des microvésicules dans les cellules endothéliales ... 65
3-1-5 Test de migration cellulaire avec les inhibiteurs d’endocytose, de transcription et de traduction... 66
3-1-6 Analyses statistiques ... 66
3-2 Résultats ... 67
3-2-1 Photographies au microscope fluorescence ... 67
3-2-2 Test de migration cellulaire en présence de l'inhibiteur de l'endocytose ... 68
3-2-4 Test de migration cellulaire en présence d’un inhibiteur de la traduction ... 71
Chapitre 4- Discussion et Conclusion ... 72
4-0 Discussion et Conclusion ... 72
Liste des abréviations
Français Anglais
Act-D Actinomycine D Actinomycin D
ADN Acide Désoxy Ribonucléique Desoxyribonucleic Acid Ang-2 Angiopoïétine Angiopoietin
AP2 Protéine adaptatrice 2 Adaptive Protein 2 ARN Acide ribonucléique RiboNucleic Acid ATP Adénosine triphosphate Adenosine triphosphate
bFGF Facteur de croissance des basic Fibroblasts Growth Factor
fibroblastes basique
CA Corps Apoptotiques Apoptotic bodies CE/EC Cellules Endothéliales Endothelial Cells
CEMV/HMEC Cellules Endothéliales Human Microvascular Endothelial Cells
Microvasculaire de peau
CFDA CarboxyFluorescéine CarboxyFluorescein DiAcetate
DiAcetate
CHX Cycloheximide Cycloheximide
CMH Complexe Majeur Major Histocompatibility Complex
d'Histocompatibilité
CMV/MVBs Corps MultiVésiculaire Multi Vesicular Bodies
DAVID Base de données pour Database for Annotation,Visualization and
l’annotation, la visualisation Integrated Discovery
et découverte intégrée
DMEM Milieu d’Eagle modifié Dulbecco's Modified Eagle’s Medium
par Dulbecco
Dyn Dynasore Dynasore
EBM-2 Milieu basal pour cellules Endothelial Basic Medium-2
endothéliales
ED-A Variant de la fibronectione Fibronectin variant
EDC Endocytose Dépendant Caveolin-Dependant Endocytosis
de Cavéoline
EGF Facteur de croissance Epidermal Growth Factor
épidermique
EGFR Recepteur du facteur de Epidermal Growth Factor Receptor
croissance des fibroblastes
ELISA Dosage d'immunoadsorption Enzyme-Linked Immunosorbent Assay
par enzyme lié
EGM-2MV Milieu pour la croissance Endothelial Cell Growth Medium-2
des cellules endothéliales MicroVascular
EMC Endocytose Médié par Clathrine Mediated Endocytosis
Clathrine
ES MS/MS Spectrométrie de masse ElectroSpray Mass Spectrometry
Exo Exosomes Exosomes
FACS Trieur de cellules marquées par Fluorescence-Activated Cell Sorting
fluorescence
Fb Fibroblastes Fibroblasts
FGF Facteurs de croissance Fibroblats Growth Factor
des fibroblastes
GAG Glycosaminoglycane GlycosAminoGlycan
GMCSF Facteur stimulant les colonies Granulocyte-Macrophage Colony-Stimulating de granulocytes-macrophages Factor
GTP Guanosine-5'-TriPhosphate Guanosine-5'-TriPhosphate HIF Hypoxy Inducible Factor Hypoxia Inducible Factor HSP Protéine de choc thermique Heat Shock Protein
HSPG protéoglycane de sulfate Heparan Sulfate Proteoglycan
d'héparine
ICAM-1 Molécule d'Adhérence Intercellular Adhesion Molecule-1
Intercellulaire-1
IEF Point Isoélectrique IsoElectric Focusing
IGF Facteur de croissance Insuline-like Growth Factor
semblable à l'insuline
IL-1 Interleukine-1 Interleukin-1
IPG Gradient de pH immobilisé Immobilized pH gradient
KEGG Encyclopédie de Kyoto des Kyoto Encyclopedia of Genes and Genomes
gènes et des génomes
KGF Facteur de croissance des Keratinocyte Growth Factor Kératinocytes
LC-MS/MS Chromatographie en phase Liquid Chromatography–Mass Spectrometry liquide-spectrométrie de masse
LOEX Laboratoire d'Organogenese Laboratory of Organogenesis
Expérimentale Experimental
MCP-1 Proteine chemoattratante Monocyte Chemoattractant Protein
pour les monocytes -1
MEC/ECM Matrice ExtraCellulaire ExtraCellular Matrix
MET transition Mesenchymal-Epithelial transition
épithéliale-mésenchymateuses
MMC Mitomycine C Mitomycin C
MOI Multiplicité de l'infection Multiplicity of Infection MV Microvésicules Microvesicles
MβCD Méthyl-β-CycloDextrine Methyl-β-CycloDextrin
NKG2D cellules tueuses naturelles Natural Killer Group 2D
Groupe 2D
NTA analyse de suivi de Nanoparticle Tracking Analysis
nanoparticule
PAF Facteur d'Activation des Platelet-Activating Factor
Plaquettes
PBS Tampon phosphate salin Phosphate Buffered Saline
PDEGF Facteur de croissance Platelet-Derived Epidermal Growth Factor
épidermique dérivé des plaquettes
PDGF Facteur de croissance dérivé Platelet-Derived Growth Factor
des plaquettes
PAGE Electrophorèse sur gel de PolyacrylAmide Gel Electrophoresis
polyacrylamide
RAC-1 Substrat 1 de la toxine Ras-related C3 botulinum toxin substrate1
botulique C3 liée à Ras
SDS Dodécylsulfate de sodium Sodium Dodecyl Sulfate SNARE Récepteur soluble de la protéine Soluble NSF Attachment Protein REceptor
attachée à NSF
SVF/ FBS Sérum de Veau Fetal Fetal Bovine Serum TGF β Facteur de croissance Transforming Growth Factor β
transformant β
TIMP Tissu Inhibitor of Metalloproteinase TNF-α Facteur de nécrose tumorale Tumor Necrosis Factor alpha
alpha
TSG101 Gène de susceptibilité Tumor susceptibility gene 101
Tumorale 101
VE/EV Vésicules Extracellulaires Extracellular Vesicles
VEGF Facteur de croissance de Vascular Endothelial Growth Factor
l'endothélium vasculaire
Wmyo myofibroblaste cicatriciel Wound myofibroblast α-SMA Actine de muscle lisse alpha alpha smooth muscle actin
2D-DIGE Electrophorèse en gel de Second Dimension-DIfference Gel Electrophoresis seconde dimension
Liste des tableaux
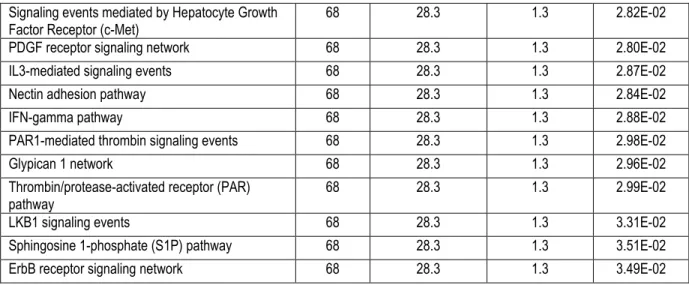
Tableau 1:Comparaison entre les trois types de vésicules extracellulaires ... 24 Tableau 2 :Results of functional annotations clustering analysis performed with the
DAVID database for the six clusters with the highest enrichment score and that are
noteworthy for interpreting the estimated role of MVs. ... 54 Tableau 3 : Enrichment analysis in the GO term “Cellular component” using the FunRich database as background. ... 55 Tableau 4 :Enrichment analysis in pathways using the FunRich database as background. 57 Tableau 5:Full dataset of all the proteins detected in MV samples including accession number, description and molecular weight... 99 Tableau 6: Results of functional annotations clustering analysis performed with DAVID database. ... 133
Liste des figures
Figure 1: Représentation schématique de la peau humaine et de ses principaux constituants ... 4 Figure 2 : schéma présentant les différentes phases de la cicatrisation ... 9 Figure 3 : Représentation schématique des phases de l’angiogénèse par bourgeonnement pendant la cicatrisation. ... 13 Figure 4: schéma présentant la différenciation des fibroblastes en Myofibroblastes pendant la cicatrisation cutanée... 16 Figure 5 : Formation et libération des exosomes, des microvésicules et des corps
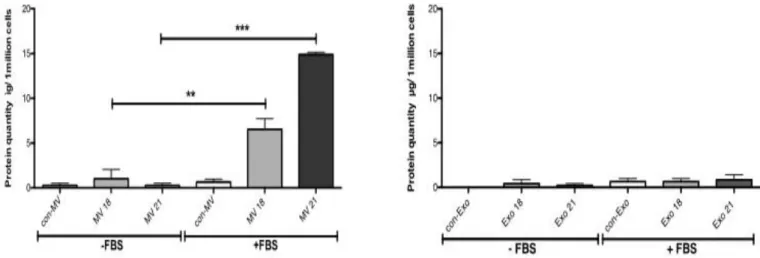
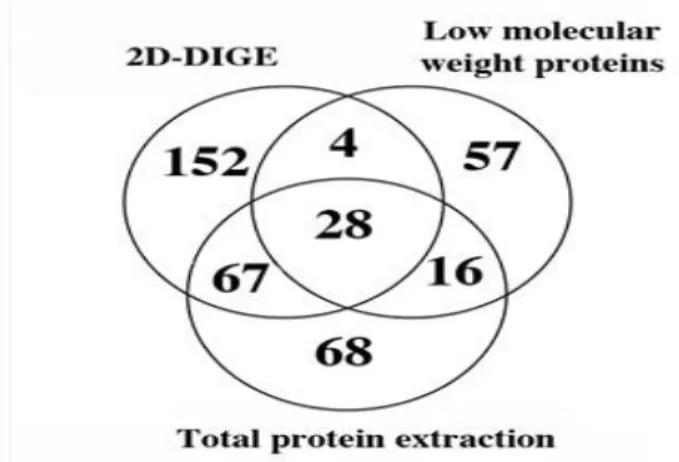
apoptotiques ... 18 Figure 6 : Mode de communication des vésicules extracellulaires avec les cellules cibles.31 Figure 7: Serum stimulated the release of MVs by WMyos but not the release of exosomes ... 58 Figure 8: Three-way Venn diagram showing the overlap between MV proteins isolated using the 2D-DIGE method. ... 59 Figure 9 :MVs produced from DsRed transduced Wmyo contained DsRed and transferred fluorescent staining to HMEC when added on their culture medium. ... 60 Figure 10: The MV addition in culture medium stimulated endothelial cell growth. ... 61 Figure 11: HMEC migration rate was increased in presence of MVs. ... 62 Figure 12 : MVs stimulated capillary-like formation by HMECs in Matrigel®-reduced growth factors. ... 63 Figure 13 : Photographies au microscope à fluorescence des CEMV 14 non fixées ... 67 Figure 14: Photographies au microscope à fluorescence des CEMV 14 fixées ... 68 Figure 15 : Test de scratch réalisé avec des CEMV14 en présence du Dynasore® et des MV de Wmyo ... 69 Figure 16 : Test de scratch réalisé sur les CEMV14 en présence de l'actinomycine D® et des MV de Wmyo. ... 70 Figure 17 : Test de scratch de CEMV 14 en présence de cycloheximide® et des MV
deWmyo ... 71 Figure 18 : Schéma représentant les étapes d’endocytose dépendant de la clathrine et son inhibition par le dynasore. ... 76 Figure 19: Schéma représentant le site d’intervention de l’actinomycine D dans le
mécanisme de la transcription. ... 77 Figure 20 : Schéma représentant l’implication de la cycloheximide dans l’inhibition de traduction. ... 78
À une ville que j’ai la fierté d’appeler chez moi, une ville qui m’a grandi solidement, une ville qui souffre, dans l’espoir qu’elle retrouve la paix, ma chère Alep…
À un père vaillant, pour son amour gratuit, son sacrifice énorme, son soutien irremplaçable, Anis... À une mère héroïque, qui croit énormément en moi, m’accompagne comme jamais, et m’apprend à
aller jusqu’à bout de mes limites, Rania…
J’espère que vous serez toujours fiers de moi, je vous offre ce travail
Un scientifique dans son laboratoire n’est pas un simple technicien : il est aussi un enfant confronté à des phénomènes naturels qui l’impressionnent comme dans des contes de fées.
Marie Curie
Remerciement
En premier lieu, je tiens à remercier sincèrement ma merveilleuse directrice de recherche Dre. Véronique Moulin. Véronique, merci mille fois pour m’avoir acceptée et encadrée dans ton laboratoire de recherche, tout au long de ma maitrise. Tu m’as accompagnée comme jamais. Le temps que tu as passé à m’expliquer et me guider malgré ton agenda très chargé est énormément apprécié. Tu étais toujours disponible avec ton sourire encourageant, ton esprit optimiste et ton énergie stimulante. Grâce à toi, ces deux ans de maîtrise ont été une expérience incroyable. Tu m’as offert un projet très intéressant qui m’a permis de progresser et m’a appris à affronter mes difficultés et repousser mes limites. Il n’y a rien de plus satisfaisant que d’obtenir de beaux résultats. J’espère également que j’étais à la hauteur de tes attentes.
Dre Moulin, je te souhaite les meilleures chances dans ton laboratoire. Merci encore pour la confiance que tu m’as accordée et pour être une directrice exceptionnelle, j’ai été chanceuse!
Un gros merci à Sébastien Larochelle. Je tiens à te témoigner de ta compétence incroyable et de tes idées enrichissantes. Le projet n’aurait pas pu marcher aussi bien sans ton aide précieuse.
Je remercie également Dr. Francois Auger pour m’avoir donner la chance de m’intégrer dans un laboratoire aussi développé, varié, et organisé. Si quelqu’un veut apprendre la culture cellulaire, c’est au LOEX. Toute ma gratitude pour votre généreux accueil.
Un grand merci à mes deux stagiaires d’été Manon Salvetti et Marion Pierre. C’est toujours plaisant de travailler en équipe. Merci pour votre aide très apprécié dans les expériences, vos discussions intéressantes et aussi pour la correction de mes fautes de français ! C’était un plaisir de travailler avec vous.
Merci à Alexandra Laberge pour avoir partagé avec moi des moments stimulants dans le R-231b, pour sa collaboration et sa présence pour m’aider et m’encourager.
Je remercie toute la gang du LOEX, un merci spécial à tous les assistants de recherche qui travaillent fort pour préparer et surveiller le matériel commun. Merci aux collègues, surtout les amis de bureau pour le temps agréable que j’ai passé avec vous, Angela Dakiw Piaceski, Josy Siebra Naud, Kiefer Thouin, Thierry de Serres-Bérard, Vincent Roy, Sergio Cortez Ghio, Meryem Safoine, Cassandra Goulet, Fabien Kawecki, et Priscilla Hoarau. Vous m’avez offert une famille de plus et deux ans de nombreux rires.
Je remercie tous les amis de ma chère ville d’Alep, spécialement les amis scouts des frères maristes, qui malgré leurs difficiles conditions et malgré la distance, étaient toujours là pour moi. Vos encouragements et votre partage m’ont stimulée pour travailler mieux.
Finalement je remercie ma chère famille, qui a été toujours supportant, autant dans la distance que dans la proximité; Line pour être une sœur exceptionnelle, merci pour ta disponibilité si précieuse, et tes grands oreilles pour m’écouter; Chahem pour être un frère génial, merci pour ton soutien et ton partage de ma fatigue ainsi que de mes réussites pendant ces deux ans; Becher pour ton esprit si spécial, merci pour tes encouragements et tes bons mots stimulants; Perla et Georges pour être si proches même dans la distance, merci pour votre forte influence et votre présence motivante. Vous êtes tous merveilleux la famille, et comme on dit toujours : On PEUT ! n’est-ce pas ?!
Hypothèse et objectifs
Les microvésicules (MV), sont des vésicules produites par bourgeonnement de la membrane plasmique des cellules. Notre équipe a démontrée précédemment que les myofibroblastes (Wmyo), des cellules de la peau qui apparaissent pendant la cicatrisation, produisent des MV. Ces MV pourraient être impliquées dans la communication intracellulaire pendant la guérison de plaie. Nous avons précédemment détecté par spectrométrie de masse, 381protéines présentes de façon constante dans les MV produites par 6
populations de Wmyo. Par le logiciel DAVID, 19protéines impliquées dans le développement de vaisseaux sanguins/angiogenèse ont été identifiées. De plus, la présence de VEGF et de FGF2 dans les MV a été détectée par ELISA.
Mon hypothèse est que les MV produites par les Wmyo cicatriciels pourraient jouer un rôle dans l’angiogenèse puisque ce mécanisme est régi par des interactions cellulaires et que les MV agissent comme transporteurs pour les médiateurs intercellulaires
Mes Objectifs sont divisés en deux sous-objectifs principaux ; premièrement, analyser le rôle des MV isolées de Wmyo sur le processus de l’angiogenèse en évaluant la réponse des cellules endothéliales microvasculaires de peau selon les trois mécanismes fondamentaux de la formation des capillaires sanguins : la croissance de CEMV qui a été évaluée par comptage cellulaire après traitement pendant 6 jours. La migration des CEMV qui a été évaluée par un test de Scratch pendant 24 heure. Enfin, la formation des structures tubulaires qui a été quantifiée en ensemençant les CEMV sur du Matrigel ® pendant 24heures.
La présence de MV a induit une augmentation significative et dose-dépendante de la croissance, de leur vitesse de migration, ainsi que de la formation des structures en tubules typiques des capillaires sanguins. Ces résultats positifs m’ont amené à mon deuxième objectif : étudier les mécanismes probables de l’interaction et de l’action de MV avec et sur les cellules endothéliales cibles en ajoutant des MV fluorescentes sur les CEMV. Les résultats du microscope à fluorescence ainsi que la revue de la littérature m’a conduit à évaluer le mécanisme le plus probable de l’interaction des MV avec la cellule cible ; l’endocytose, l’implication de cette voie a été étudiée par le test de scratch en ajoutant un inhibiteur spécifique. Ensuite l’action de MV sur les CEMV par la transcription et la traduction protéique a été évaluée de la même manière selon les inhibiteurs spécifiques.
Chapitre 1-Introduction 1-1 La peau
La peau est un organe étendu recouvrant le corps complet. Étant le plus grand organe chez les mammifères, cet organe occupe 1-2 m² de surface et à un poids de 4 à 5 kg. La peau se compose de trois couches successives, de l'extérieur vers l’intérieur du corps : l’épiderme, le derme et l’hypoderme [1, 2].
1-1-1 les trois couches de la peau : 1-1-1-1 L’épiderme
L'épiderme est la partie externe de la peau. C’est un tissu épithélial sans vaisseau sanguin. Il est constitué principalement de kératinocytes (95%) dont le rôle est de protéger la peau des éléments extérieurs grâce à la présence des kératines dans leur cytosquelette [3]. La division des kératinocytes au niveau des cellules souches de la couche basale, la couche la plus profonde de l’épiderme, et leur différenciation dans les couches superficielles forment les différentes couches de l'épiderme et permettent son renouvellement tous les 26 à 42 jours [4]. Les autres composants de l’épiderme, en pourcentage beaucoup moins élevé, sont les mélanocytes, cellules responsables de la pigmentation et de la protection de la peau contre les rayons solaires en produisant la mélanine ; les cellules de Langerhans qui prennent en charge la réponse immunitaire, et les cellules de Merkel servent de mécanorécepteurs. En plus de ces cellules, la présence de pores dans l’épiderme lui confère la capacité de réguler la perte des fluides et la température du corps [3].
1-1-1-2 Le derme
Le derme est le tissu conjonctif sous-jacent à l'épiderme. Les deux tissus sont séparés par un dépôt matriciel spécialisé appelé membrane basale produite conjointement par les cellules des deux tissus. Les deux composants majoritaires du derme sont la matrice extracellulaire et les cellules fibroblastiques. Il contient également des vaisseaux sanguins et lymphatiques, des glandes sébacées et sudoripares, des follicules pileux, des terminaisons nerveuses, des mastocytes, et des dendrocytes. Des lymphocytes et macrophages s’y trouvent en petit nombre [5].
1-1-1-2-1 La matrice extracellulaire
La matrice extracellulaire (MEC) de la peau lui confère d’importantes caractéristiques mécaniques telles la souplesse et l’élasticité[2]. Elle est composée de fibres de collagène (collagène I et III) et élastiques (élastine et fibrilline), de protéines de liaison (fibronectine, tenascine) et des protéoglycans (décorine, lumican, fibromoduline) [6]. Ces différents composants entourent tous les annexes du derme et aident les cellules dermiques à adhérer, à proliférer, à se différencier, et à migrer [7]. Les protéines de la MEC permettent également la liaison temporaire de nombreux facteurs de croissance cellulaire [8]. L'équilibre quantitatif entre le dépôt de la MEC et sa dégradation est primordial afin d’éviter des pathologies dermiques telles que les fibroses (cicatrices hypertrophiques).
1-1-1-2-2 Les fibroblastes
Les fibroblastes (Fb) sont les cellules majoritaires du derme. Ils produisent, déposent et remodèlent la MEC, afin de maintenir l'intégrité de la structure des tissus conjonctifs [9]. Le mésenchyme primitif est à la source des Fb[10].
1-1-1-3 L’hypoderme
L’hypoderme est la couche la plus profonde de la peau. Il permet la fixation de la peau aux muscles et à l'os sous-jacent. Il est constitué principalement de tissus adipeux formés d’adipocytes remplis d’acides gras et de triglycérides, d’un réseau important de capillaires sanguins, de Fb et de macrophages. Ce tissu joue un rôle dans la thermorégulation, dans la protection contre les stresses mécaniques et dans le stockage de l’énergie. Environ près 50% de tout le gras présent dans le corps se retrouve dans cette couche [13].
Figure 1: Représentation schématique de la peau humaine et de ses principaux constituants
Figure tirée du site web biologie de la peau (http://biologiedelapeau.fr/). Libre distribution
1-1-2 Les fonctions de la peau
La peau représente un facteur important pour la santé et la survie d’un organisme. De nombreuses fonctions sont conférées par la peau :
La protection contre les menaces environnementales externes : Puisqu'elle est une barrière anatomique entre l'intérieur et l'extérieur du corps humain, la peau protège le corps contre des agressions chimiques, physiques, biologiques et de colonisation infectieuse. Certaines des cellules présentes dans la peau font partie du système immunitaire [14].
Les glandes sudoripares et les vaisseaux sanguins de la peau participent à la thermorégulation [5, 14, 15].
La peau est une barrière relativement sèche et semi-imperméable qui contrôle l’évaporation. La peau est impliquée dans la synthèse de la vitamine D [14, 15].
Grâce aux terminaisons nerveuses présentes dans la peau, celle-ci est responsable de diverses sensation [5, 15].
La peau renferme un réservoir sanguin important de par les nombreux vaisseaux sanguins qui traversent le derme et l’hypoderme. Environ 10 % du sang en circulation dans l'ensemble du corps est présent dans cet organe [5, 14]
Enfin, la peau contribue à l'absorption et au stockage des nutriments essentiels (eau, lipides).[5, 14, 15]
1-2 La cicatrisation cutanée
Lorsque la rupture traumatique de la barrière cutanée est assez profonde pour détériorer l'intégrité de la peau, alors une plaie survient [16]. Le degré de gravité de cette blessure varie selon plusieurs paramètres comme la profondeur, la surface et la nature de la plaie [17]. Le mécanisme de réparation des couches abimées est un processus physiologique qui implique des évènements dynamiques coordonnés tels la croissance cellulaire, la migration cellulaire et la production de la MEC [17]. De nombreux facteurs modulent la vitesse de guérison, y compris certaines hormones/cytokines ainsi que l’ampleur de la circulation sanguine dans la plaie. [18].
1-2-1 Les phases de la cicatrisation
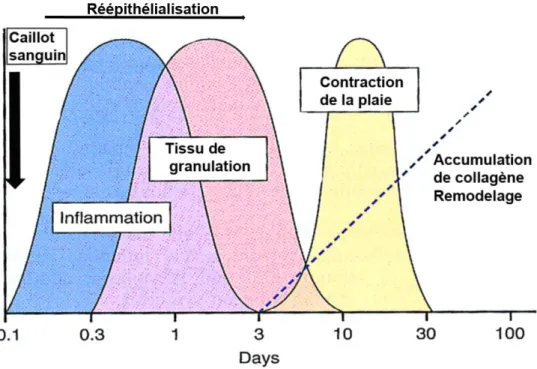
Le modèle classique du processus cicatriciel se divise en 3 phases qui se chevauchent tout au long de son évolution graduelle : 1) l'inflammation. 2) la prolifération et la migration de cellules permettant la formation du tissu de granulation en profondeur, et la ré-épithélialisation superficiellement. 3) la phase de remodelage ou maturation de tissu de granulation dont le but est d'obtenir un nouveau derme presque aussi solide ??? que le derme initial. [19, 20].
1-2-1-1 La phase vasculaire
La lésion vasculaire conduit à une vasoconstriction prolongée et accrue grâce aux substances libérées par les plaquettes telles que le thromboxane A2. Cette vasoconstriction ralentit le débit sanguin. Elle s'établit par l’activation de la thrombine lorsque des vaisseaux sanguins sont lésés. La thrombine transforme une protéine soluble, le fibrinogène, en fibrine, suite à l’activation de cascade de coagulation [21]. Les plaquettes s’activent également et forment un agrégat [22]. Afin de consolider l'agrégat plaquettaire, la fibrine s’agglutine avec les plaquettes, et forme le caillot sanguin [23]. Ce dernier joint les bordures de la plaie et sert de nouvelle MEC [24, 25]. Cette matrice temporaire a comme fonctions d’arrêter l'écoulement sanguin, et de fournir une matrice provisoire servant de support aux cellules endothéliales (CE), fibroblastiques, et inflammatoires (macrophages et polynucléaires) migrant dans la plaie [26, 27]. A ce stade, certaines cytokines, ou facteurs de croissance, comme le PDGF, le TGFß, l'IGF ou l'EGF, sont secrétés par les plaquettes puis par les macrophages et participent à la différenciation des Fb [22].
1-2-1-2 L’inflammation
Le rôle principal de la phase inflammatoire est la détersion de la plaie. L’inflammation est une réaction du tissu vascularisé de courte durée contre une agression locale, elle dure environ 3 jours dans les plaies de petites surfaces mais peut durer plus longtemps selon la taille de la plaie [24]. La lésion vasculaire et le phénomène de coagulation stimulent la libération de cytokines, ce qui provoque une vasodilatation locale et une augmentation de la perméabilité capillaire. Les vaisseaux deviennent perméables favorisant la migration des monocytes vers la lésion et la création d’un œdème [24].
Certains médiateurs biochimiques provenant des débris de l'agrégation plaquettaire et des bactéries favorisent par chimiotactisme l’invasion des leucocytes inflammatoires dans la plaie[24]. Dans un premier temps les neutrophiles s'accumulent au niveau de la lésion et agissent à la surface de la plaie. Une fois dans la plaie, ils la nettoient en phagocytant les molécules externes [28]. Enfin, les neutrophiles disparaissent par apoptose et les monocytes prennent le relais du nettoyage. Les monocytes envahissent la région de la plaie en réponse aux certains facteurs comme le TGFβ et le MCP-1. Les monocytes actifs se différencient en macrophages grâce aux facteurs locaux. Le rôle des macrophages est multiple, ils participent au nettoyage de la plaie, ils s’impliquent dans la
réponse immunitaire adaptative en présentant les antigènes aux lymphocytes [29], et ils stimulent également la phase de réparation en sécrétant des facteurs tels que le PDGF et stimulent la prolifération des lymphocytes, des Fb, des kératinocytes et des CE [22].
1-2-1-3 La prolifération et la migration cellulaire.
Cette phase comporte plusieurs étapes et dure environ 10 à 15 jours. Elle aboutit à la formation d’un tissu provisoire appelé tissu de granulation[30].
La formation du tissu de granulation commence quelques heures après la blessure et débute avant même que la phase inflammatoire ne soit complétée. Plusieurs types de cellules vont se déplacer par chimiotactisme vers/dans la plaie [25]. Ce tissu provisoire se compose de cellules inflammatoires, de Fb, de Wmyo, de nouveaux vaisseaux sanguins, le tout supporté par une nouvelle MEC [27]. La MEC provisoire a une composition différente de la MEC présente dans les tissus normaux et ses composants sont produits par les Fb et les Wmyo. Elle se compose de fibronectine, collagènes, élastine, glycosaminoglycanes (GAG), glycoprotéines et protéoglycanes. Avec le temps, cette matrice provisoire est remplacée par une MEC permanente qui forme un tissu plus solide.
Les macrophages sécrètent des facteurs de croissance stimulant la migration des Fb et des CE [27, 31].La migration de Fb vers la plaie est à l’origine de la transformation de la MEC temporaire en tissu de granulation [26].
Les cellules fibroblastiques se différencient en Wmyo suite à une exposition d’un stress mécanique et leur stimulation par certains facteurs de croissance principalement le TGFβ [32]. Les Wmyo, à leur tour, facilitent la fermeture de la plaie en contractant ses bords.Les CE vont être responsables de la réparation et de la croissance des vaisseaux sanguins endommagés de façon à nourrir les nouveaux tissus cicatriciels [33].
La réépithélialisation se met en place lorsque les kératinocytes qui forment l’épiderme migrent vers le centre de la plaie [27]. Après activation de ces cellules, une dissolution des desmosomes et des hémidesmosomes survient. Ceux-ci permettent l’attachement des cellules entre elles et l’attachement des cellules à la membrane basale [27]. Les kératinocytes en migration recouvrent la
la blessure [23, 34]. Une fois que la première couche de kératinocytes est formée, la migration de ceux-ci s’arrête et commencent leurs multiplication et différenciation [20, 24]. Graduellement, la membrane basale est reconstruite, et l’épiderme est colonisé par des mélanocytes. Les kératinocytes, comme les macrophages, produisent des facteurs de croissance qui stimuleront l’angiogenèse et la fibrogénèse dans le derme [20, 24].
1-2-1-4 le remodelage et maturation
Cette phase peut durer plus d’un an. Les facteurs influençant la durée de cette phase sont : l’âge, la génétique, la localisation, la surface et la gravité de la plaie, la durée et la puissance de la phase inflammatoire. [23, 34].
La phase de remodelage est considérée comme la phase de transition entre le tissu de granulation et la cicatrice finale. Elle se définit par des mécanismes de réparation plus lents, entraînant un retour à l’état normal; les fibres de collagène commencent à être mieux structurées : initialement, fines et désorganisées, leur diamètre devient plus important et elles se rattachent aux autres fibres des bords de la plaie [23, 34]. En plus des fibres de collagène, d’autres constituants comme les protéoglycans et les GAG s’ajoutent à la MEC qui se réorganise en contrôlant l’équilibre entre la production et la dégradation continue de ses constituants [27]. Ce remodelage implique une coordination précise des MMP et de leurs inhibiteurs (TIMP) qui sont sécrétés par plusieurs types cellulaires [27]. Graduellement, l’épiderme récupère son épaisseur physiologique, les cellules en excès qui demeurent après la formation du tissu de granulation entrent en apoptose et les vaisseaux sanguins retrouvent leur taille initiale [35].
Les trois phases cicatricielles se développent d’une manière organisée et selon un schéma précis impliquant une coordination très importante entre ces différentes étapes afin d’éviter les cicatrices pathologiques telles que les cicatrices hypertrophiques et la fibrose ou la formation d’ulcères.[24, 36].
Figure 2 :schéma présentant les différentes phases de la cicatrisation
Figure adaptée de site web : https://360woundcare.wordpress.com. Libre distribution.
1-3 L’angiogenèse
1-3-1 Description de l’angiogenèse
Le mot angiogenèse se dérive du mot grec Angeion qui veut dire : formation de vaisseaux sanguins à partir de vaisseaux préexistants. Dans le corps, les cellules métaboliquement actives sont à moins de quelques centaines de micromètres d'un capillaire de sang. Ce capillaire fournit aux cellules les nutriments et l'oxygène nécessaire et leur permet d'éliminer les déchets métaboliques générés par leur activité.
précurseurs vasculaires [37]. L'angiogenèse va plutôt permettre l'accroissement du réseau primaire par la formation de plus petits vaisseaux, vascularisant ainsi les organes et les tissus en développement [37]. Il faut par contre noter que parfois des cellules souches progénitrices endothéliales interviennent également dans le processus angiogénique [38, 39]. Dans l'organisme, on distingue de nombreux facteurs de croissance ayant une puissance proangiogénique ou une action inhibitrice. Au repos, il existe un équilibre entre les inducteurs et les inhibiteurs. Dans un contexte de réparation tissulaire, l'équilibre est bouleversé et un déplacement en faveur des inducteurs survient. Deux protéines particulièrement importantes interviennent: le bFGF (basic fibroblast growth factor) [27, 40] et le VEGF (vascular endothelial growth factor) [27, 31, 40]. L’expression de ces deux facteurs est stimulée suite à la sécrétion de plusieurs molécules angiogéniques notamment le TGFß, mais également lorsque les niveaux d’acide lactique sont élevés ou la quantité d’oxygène est basse [27, 41].
Le contrôle de l'angiogenèse comme outil thérapeutique a entraîné un grand intérêt pour ce phénomène au cours des 40 dernières années. La stimulation de l'angiogenèse peut être bénéfique, comme dans le processus cicatriciel et la maladie cardiaque ischémique. En revanche, réduire ou inhiber l'angiogenèse peut également être thérapeutique dans le cas des cancers, certaines conditions ophtalmiques, l'arthrite rhumatoïde ou d’autres maladies. [27, 42].
1-3-2 L’angiogenèse pendant la cicatrisation
Dans un tissu en réparation, le rétablissement d’un réseau vasculaire est nécessaire afin de permettre la survie des cellules. Dans une situation de cicatrisation, l’angiogénèse intervient particulièrement par bourgeonnement [41, 43]. Le TGFβ est une cytokine clé de la cicatrisation, elle s'implique dans différents aspects tels que la migration, l’angiogenèse, la fibrogénèse, et la production des constituants de la MEC [41].
Les étapes de l’angiogénèse par bourgeonnement peuvent se diviser en une série de mécanismes décrits ci-dessous:
1-3-2-1 La prolifération des cellules endothéliales
Dans un état physiologique, les CE des vaisseaux sanguins se trouvent dans un état de quiescence. [44]. En revanche, pendant le mécanisme cicatriciel, elles s'activent et se mettre à proliférer et à
migrer de façon à construire un nouveau réseau microcirculatoire pour assurer l'irrigation sanguine adéquate du tissu en réparation [33].
Pour que l’angiogénèse progresse, les récepteurs de la fibronectine présente pendant la formation du bouchon hémostatique doivent s’activer à la surface des CE [27]. De plus, du VEGF et du bFGF doivent être libérés par les plaquettes, favorisant l'activation du processus angiogénique dès le début [27, 30, 45].
Les cellules autour du tissu blessé participent également à l'activation des CE et à la réaction inflammatoire en relâchant des cytokines et des facteurs de croissance [38, 46]. Par exemple, les kératinocytes sécrètent du TGFß, du VEGF, du bFGF, du PDGF et du IL-1, alors que les Fb expriment le TGFß, le bFGF, et le PDGF [46-48]. Les cellules engagées dans l'inflammation prennent part aussi à l'activation des CE par la libération de VEGF, de bFGF, de TGFß et d'autres cytokines intervenant plus ou moins directement.
L'autre déclencheur important de l'angigogènése est l'hypoxie. Le manque d'oxygène engendré par les bris des capillaires sanguins sensibilise les cellules qui, à leur tour, expriment la protéine HIF (Hypoxia Inducible Factor). Cette protéine stimule l'expression des facteur pro-angiogéniques tels que l'Ang-2 et le VEGF [49].
L'hémostase, l'inflammation et l'hypoxie contribuent donc à la forte stimulation angiogénique au site lésionnel ce qui aboutit à une déstabilisation rapide des vaisseaux avoisinants. Une dilatation des vaisseaux et la dégradation de leur membrane basale se met alors en place, les péricytes se libèrent, et les CE perdent leur état de quiescence et se détachent les unes des autres. Bien que l'ensemble du processus impliqué dans le mécanisme d'activation n’est pas encore clarifié, le rôle du VEGF et de l’Ang-2 semble primordiale, afin d’activer les CE [50].
1-3-2-2-la migration des cellules endothéliales
Comme écrit précédemment, les CE reposent sur leur membrane basale dans un état de quiescence physiologique. Suite à une blessure du vaisseau, les CE en périphérie de la plaie s'activent et migrent pour reconstituer le réseau vasculaire endommagé. La barrière empêchant le déplacement des cellules est la membrane basale. La protéolyse matricielle résout ce problème physique et
matricielles entraine l'exposition de nouveaux sites biologiquement actifs qui stimulent de nombreux mécanismes biologiques dont la migration des cellules et la régulation du processus angiogénique [51].
En parallèle, cette dégradation permet de relâcher les facteurs de croissance qui s’y attachent. Ces derniers encouragent le propagation angiogénique en créant un gradient qui guide le déplacement des cellules par chimiotactisme [52].
La dégradation de la matrice libère également des inhibiteurs de l'angiogenèse que l'on appelle les matrikines. La digestion enzymatique de ces protéines les transferts d'un état inactif à un état actif [53].
La dégradation de la matrice est donc importante dans l'ensemble de l'angiogénèse. Il existe une gamme abondante de protéases engagées dans la migration et dans la formation des capillaires. L'hétérogénéité des molécules impliquées dans ce processus confère aux cellules une adaptation aux différents composants de la matrice sur laquelle les cellules doivent migrer [52, 54, 55].
1-3-2-3-la formation de vaisseaux sanguins et la maturation
Au cours de la néovascularisation, les CE migrent selon un mécanisme chimiotactique contrôlé et dans une direction fixe. Une suite d'évènements basés sur l'expression de protéines mène à l'adoption d'un phénotype spécifique pour les cellules en tête de file, et un autre phénotype pour les cellules qui les suivent [56]. Les cellules en tête de file (tip cells) sont les cellules apicales. Les cellules qui suivent (stalk cells) sont les cellules de queue. Ces cellules ne répondent pas de façon similaire à une stimulation avec le VEGF [57]. Le VEGF active les premières cellules qui auront alors un phénotype de cellules apicales et se diviseront afin d'augmenter le diamètre du vaisseau. La cellule apicale ne forme pas de lumière [58], elle renonce à sa polarité, mais elle reste toujours attachée aux cellules suivantes [58]. Les cellules adjacentes se multiplient, se polarisent et conduisent à la formation de la lumière du vaisseau récemment établit [58, 59]. Le processus d'anastomose coopère au mécanisme d’angiogénèse; les capillaires provenant des artérioles et des veinules se rejoignent, amenant un flux sanguin continu dans la plaie.
Finalement, le réseau vasculaire atteint sa fonctionnalité par l'amélioration de ses structures et par sa stabilisation. Le mécanisme de maturation exige un remodelage de la structure vasculaire,
impliquant une augmentation du nombre d'embranchements et une modification du diamètre des vaisseaux. La stabilité des vaisseaux assure une bonne perméabilité et une intégrité solide pour assurer une réponse efficace aux besoins spécifiques des tissus [60].
Figure 3 : Représentation schématique des phases de l’angiogénèse par bourgeonnement pendant la cicatrisation.
Figure tirée de l’article: Notch signaling regulates tumor angiogenesis by diverse mechanisms. Dufraine, J., Y. Funahashi, and J. Kitajewski,Oncogene, 0000. 27(38): p. 5132-5137. Avec la permission de Nature Publishing Group et Copyright Clearance Center.
1-4 Les myofibroblastes
1-4-1 Description des myofibroblastes
Les myofibroblastes (Wmyo) sont des cellules de grandes tailles avec un réticulum endoplasmique très actif [61]. Ils montrent une forme étoilée et un ratio cytoplasme/noyau élevé [36, 62].
Ils ont la capacité de se différencier à partir de nombreux types cellulaires [30]. Dans la peau, les Fb sont à l'origine des Wmyo bien que d’autres études montrent que les Wmyo pourraient provenir des cellules épithéliales ou encore de la différenciation des péricytes [63] . Les Wmyo présentent un cytoplasme contenant un marqueur mésenchymateux, la vimentine, ainsi que l’isoforme de
l'alpha actine du muscle lisse (a-SMA) [64].Ce sont donc des cellules qui ont les caractéristiques des Fb et des cellules de muscles lisses [65].
La présence élevée d’a-SMA organisée en fibres de stress est le marqueur distinctif des Wmyo [66]. En effet, les Wmyo possèdent des faisceaux de microfilaments qui se terminent à la surface de la cellule dans un complexe d'adhérence spécialisée, appelée fibronexus ou adhésion locale mature. Ce complexe de microfilaments internes agit comme un pont entre les Wmyo et la fibronectine extracellulaire, servant ainsi de lien et permettant à ces cellules de transférer une force à la MEC avoisinante. Cette force contractile est maintenue au cours du temps et renforcée par le dépôt de collagène [61].
1-4-2 Les myofibroblastes et la cicatrisation
Sept à 21 jours après la blessure, on observe une modification des caractéristiques des Fb présents dans le tissu de granulation. Des fibres de stress organisées ainsi qu’un ensemble de jonctions cellulaires plus raffiné apparaissent dans le cytoplasme [67, 68] . Enfin, des replis et des indentations apparaissent au niveau du noyau et modulent sa morphologie en myofibroblastes [68] Les trois éléments connus déclenchant la différenciation des Fb en Wmyo sont le TGFβ1 [67], la fibronectine [69], et le stress mécanique [70-72].
Le TGFβ1 sécrété par les macrophages activés ayant migré dans la plaie [73] stimule la différenciation des Fb en Wmyo in vivo et in vitro [67]. Cette cytokine est reconnue comme étant l'élément déclencheur de l’apparition des structures nécessaires à la contraction.
Le variant ED-A de la fibronectine (ED-A FN) joue aussi un rôle précieux dans la différenciation. Il est nécessaire pour l'activation du TGFβ1 pendant le mécanisme de transformation des Fb en Wmyo [69]. Historiquement, ED-A FN apparaît tôt dans la cicatrisation [71], avant même l’apparition de l’a-SMA suite à la stimulation de la différenciation par le TGFβ1 [69]. Hinz et son équipe ont montré l’importance des forces de tension sur la différenciation des Fb en Wmyo ainsi que dans le maintien du phénotype [71]. La disparition de ce stress entraîne une disparition du phénotype de Wmyo [70, 71] et des points focaux matures qui apparaissent en même temps que l’augmentation de la tension sur la cellule [74]. Il existe une autorégulation positive de la différenciation des Wmyo. Les forces de tension exercées sur la cellule stimulent la production de
TGFβ1 qui, via la production de l’a-SMA, favorise la présence des forces exercées sur les cellules [72].
Au cours de la cicatrisation, Les Wmyo synthétisent la nouvelle MEC et stimulent l’angiogenèse de façon nettement plus marquée que les Fb [40]. De plus, ils possèdent une capacité contractile importanteliée à leur cytosquelette contenant le complexe actine-myosine riche en a-SMA [36], la quantité présente de cette isoforme d’actine étant proportionnelle à la force contractile des cellules [75]. Ceci permet l’accélération de la fermeture des plaies en diminuant la surface à réparer [36]. En effet, si on diminue les forces de tension dans la plaie, on peut observer une forte diminution du taux d’α-SMA ainsi que du variant ED-A de la fibronectine dans les plaies, entrainant une diminution de la capacité contractiles des Wmyo [71].
Une étroite relation existe entre les Wmyo et l'angiogenèse. Il a été établi que les CE des capillaires aident à la différenciation phénotypique des Wmyo [76], alorsque les Wmyo ont un potentiel pro-angiogénique marqué [40].
Lorsque la guérison est atteinte, Les Wmyo disparaîtront par l’apoptose [76]après avoir atteint leur maximum d’expression de l’α-SMA [35]. Quand le programme de mort cellulaire ne fonctionne pas comme il faut, les Wmyo persistent, entrainant une augmentation importante de la MEC (fibrose) et une contraction pathologique [72]. Ceci entraine plusieurs maladies fibrotiques (fibrose, cirrhose du foie,..) qui se présentent au niveau de la peau sous forme de cicatrices pathologiques appelées cicatrices hypertrophiques. [69].
Figure 4: schéma présentant la différenciation des fibroblastes en Myofibroblastes pendant la cicatrisation cutanée.
Figure tirée de l’article :Fibroblasts and myofibroblasts in wound healing.,Darby, I.A., et al., Fibroblasts and myofibroblasts in wound healing. Clin Cosmet Investig Dermatol, 2014. 7: p. 301-11. Avec la permission de Dove Medical Press LTD and Copyright Clearance Center.
1-5 Les vésicules extracellulaires (VE)
Depuis quelques années, les cellules eucaryotes sont connues pour produire une population hétérogène de structures extracellulaires nanométriques comprenant une membrane bicouche lipidique [77]. Toutes les cellules du corps peuvent produire ces VE qui sont produites via des mécanismes sécrétoires non conventionnel qui n'engagent pas la voie classique de transport sécrétoire de signal peptidique [77]. Les VE enferment de nombreux messages envoyés par la cellule à son environnement et aux autres cellules. Les VE offrent donc un moyen de communication intercellulaire unique et très important, différent des méthodes traditionnelles que sont la communication médiée par le contact directe cellule-cellule et la communication par le transfert de molécules sécrétées [78].
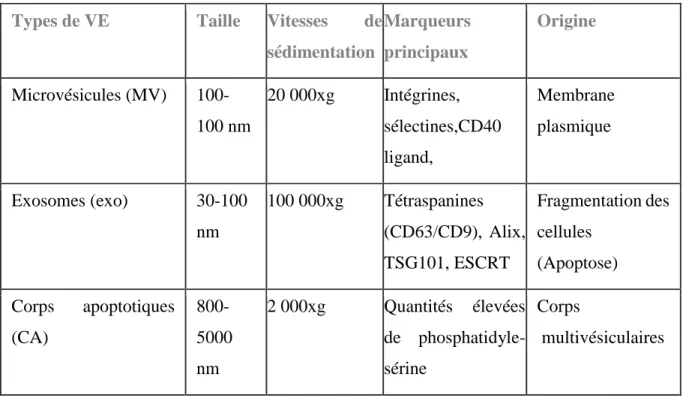
Il n’existe pas encore une méthode discriminative pour distinguer entre les types de VE, mais elles peuvent être classifiées principalement selon trois catégories, basées notamment sur leur taille et leur origine :
(1) les corps apoptotiques (CA) ont un diamètre qui varie entre 800-5000 nm, et sont libérés par les cellules subissant une mort cellulaire programmée [79];
(2) les microvésicules (MV), aussi appelées microparticules (MP) ou ectosomes sont des vésicules de diamètre 100-1000 nm qui sont produites par bourgeonnement de la membrane plasmique [79]; (3) les exosomes (exo) ont un diamètre de 30-100 nm et sont considérés comme ayant pour origine les corps multivesiculaires (CMV) qui fusionnent avec la membrane plasmique et relarguent les Exo [78, 80].
Les CA sont considérés comme un sous-produit de la mort cellulaire [79]. Ils expriment des phosphatidyl sérine (PS) sur la surface membranaire[79, 81], et ils contiennent de nombreux débris cellulaires, y compris de l'ADN fragmenté et des organites cellulaires [82]. Ils peuvent transférer des oncogènes et de l'ADN entre les cellules et se sont révélés être importantes dans la présentation des antigènes et dans l'immunosuppression [82, 83].
Bien que la libération des CA par la fragmentation cellulaire pendant l'apoptose soit connue depuis longtemps [79], le fait que la plupart des cellules vivantes produisent également des vésicules est une découverte récente [79, 84]. Concernant les MV et les exo il n’existe pas encore un principe stricte pour distinguer les uns des autres, mais les deux termes décrivent des vésicules nanométriques limités par une membrane bicouche lipidique, généralement de forme sphérique, et possédant la capacité de transférer des protéines fonctionnelles à la fois sur leur surface et à l’intérieur, des ARN non codants, des ADN, des micro ARN, et des lipides. [85].
Figure 5 : Formation et libération des exosomes, des microvésicules et des corps apoptotiques
(A) Les Exo dérivent des corps multi-vésiculaires. La fusion entre les vésicules et la membrane cellulaire conduit à leur libération. (B) Les MV proviennent de la membrane cellulaire externe lors de l'activation cellulaire. Le bourgeonnement, l'excrétion et le détachement sont les étapes principales de la formation des MV. (C) Le rétrécissement cellulaire, caractéristique de l'apoptose, conduit à la génération de CA qui contiennent les restes de la dégradation, y compris les composants cytoplasmiques et nucléaires
Figure tirée de l’article: Signaling by Extracellular Vesicles Advances Cancer Hallmarks, Kanada, M., M.H. Bachmann, and C.H. Contag,. Trends in Cancer. 2(2): p. 84-94. Avec la permission d’ Elsevier et Copyright Clearance Center.
1-5-1 Microvésicules et exosomes 1-5-1-1 Définition des MV
Les MV sont des vésicules sécrétées par les cellules, principalement par bourgeonnement de la membrane plasmique vers l'extérieur. Elles forment une population hétérogène de diamètre variant entre 100-1000 nm [79]. La libération des MV est un processus hautement régulé induit par un stimulus cellulaire comme une infection, une activation, la différenciation ou le stress. Puisque les stimulus à l’origine de la libération des MV sont nombreux, leur composants diffèrent conséquemment [81]. Les MV contiennent une variété de biomolécules comme des récepteurs, des molécules d'adhésions, des cytokines et facteurs de croissance ainsi que de l'ARNm et des microARN [79, 86, 87]. Le patron de molécules est différent selon l'agent initiateur et l'entourage extracellulaire [87]. Cette combinaison permet une augmentation de la concentration de certaines molécules dans les MV comparativement avec la concentration trouvée dans la cellule mère [87]. L’identification de l’origine de MV est possible grâce à l’expression des mêmes antigènes que la cellule parente [88].
Le protocole d’isolation des MV est différent d’une équipe à l’autre : avec ou sans pré-centrifugation à 2000 x g pour enlever les CA et les débris cellulaires [89, 90], suivi d’une centrifugation d’isolement de 13 000xg [89] ou de 20 000xg [90, 91] afin de séparer les MV. Les temps de centrifugation varient : entre 20 à 90 minutes selon les équipes [89, 92] .
1-5-1-2 Formation des MV
Même si le mécanisme exact de la formation des MV reste indéterminé , la modification de l'arrangement des phospholipides de la membrane cellulaire est le mécanisme reconnu comme étant responsable de la production des MV [93]. Dans les cellules au repos, les phospholipides sont distribués asymétriquement à travers la bicouche membranaire. Les phosphatidylcholines et les sphingomyélines sont principalement situés dans la couche extérieure, alors que les phosphatidylsérines chargés négativement et les phosphatidyléthanolamines sont situées dans la couche interne [94]. Les flippases et les floppases sont les deux transporteurs des lipides entre les couches interne et externe et sont dépendantes de l'ATP. Elles sont responsables de l'entretien de cette distribution asymétrique. Les flippases déplacent les PS de l'extérieur vers l'intérieur du feuillet, tandis que les floppases transportent les phospholipides dans le sens inverse [95]. En réponse à l'activation cellulaire ou l'apoptose, le 𝐶𝑎+2 cytosolique augmente, ce qui inhibe l'activité
des flippases et déclenche l'activation des scramblases, un transporteur bidirectionnel dépendant du 𝐶𝑎+2
[96]. L’activation des scramblases entraine une redistribution rapide et aléatoire des phospholipides dans la bicouche lipidique, perturbant l'asymétrie de la membrane. Ce réarrangement entraine une relocalisation des PS vers le feuillet extérieur, relocalisation nécessaire à la libération des MV dans la plupart des cellules [96, 97].
Un autre aspect concernant la formation des MV est sa localisation au niveau de la membrane cellulaire. Les MV sont formées spécifiquement au niveau des microdomaines membranaires hautement conservés appelés radeaux lipidiques, enrichis en cholestérol et sphingolipides. En effet, il a été rapporté qu’à la surface des MV des monocytes [98], des plaquettes [99], et des CE [100] s'exprime une grande quantité de protéines associées aux radeaux lipidiques. De plus, il a été démontré que l'intégrité structurelle et fonctionnelle des radeaux lipidiques dans les cellules est en corrélation avec la libération des MV [98, 100, 101].
Le dernier aspect important dans la formation des MV est la réorganisation du cytosquelette. Suite à la perte de l'asymétrie des phospholipides, les interactions entre les protéines du cytosquelette et
le stimulus qui déclenche la production des MV via l‘augmentation du calcium cytosolique. Par exemple, lors de l'activation des cellules, la calpaïne, une thiol protéase dépendante du calcium se déplace du cytosol vers la membrane plasmique et s'active en réponse à un agoniste médié par l’influx du 𝐶𝑎+2 extracellulaire [102-104]. La calpaïne est une enzyme pouvant induire le
désassemblage du cytosquelette en clivant l'α-actine et les microfilaments de talin, détachant ainsi la membrane plasmique du cytosquelette. Ceci permet alors le bourgeonnement des MV [97]. Ce mécanisme calpaïne-dépendant n’a pas lieu dans tous les types cellulaires. Il a été démontré que la calpaïne n'est pas impliquée dans la formation de MV induite par le complément dans les globules rouges pathologiques, ni dans la formation des MV générées dans les CE suite à un stress mécanique ou par l'angiotensine II. [100, 105]. Cepandant, d'autres mécanismes indépendants de l'apoptose ont été avancés pour initier la production de MV en réponse à certains agonistes distinctifs. [106].
1-5-1-3 La composition des MV
Les MV sont relativement hétérogènes. Les changements spécifiques qui sont induits au moment de la nucléation et du bourgeonnement des MV font que les membranes de celles-ci ne sont pas complétement identiques à la membrane plasmique de la cellule d'où elles viennent. La composition moléculaire des MV est moins bien définie par rapport aux Exo. Néanmoins, plusieurs études ont mis en évidence le fait que les MV contiennent une population diverse de protéines, y compris les métalloprotéinases matricielles (MMP) [107], les glycoprotéines, par exemple GPIb, GPIIb-IIIa et la P-sélectine [108, 109], les intégrines, par exemple Mac-1 [110], les récepteurs, par exemple, EGFRvIII [110], les composants cytosquelettiques tels que la β-actine et α-actinine-4 [111] et des ligands tels que CD40 [112]. Keerthikumar et al ont confirmé l'enrichissement des MV par des protéines mitochondriales, centrosomales, ribosomales, les protéines de ESCRT, des tétraspanines et des protéines impliquées dans la fusion et le trafic (par exemple annexines, intégrines et flotillines) [113]. Weerheim et coll. ont montré que la membrane de MV, étant formées par le bourgeonnement vésiculaire, contenait principalement de la phosphatidylcholine, de la sphingomyéline, et de la phosphatidyléthanolamine [114].
1-5-1-4 Implication des MV dans la physiologie et pathologie
Les MV ont été détéctées principalement dans le sang [115,117], le liquide synovial [90, 118], le liquide cérébrospinal [119, 120], le lait maternel [121], la salive et l’urine [122] en contiennent également.
Bien que certaines propriétés générales de MV semblent être analogues à celles des agents solubles, il semblerait que beaucoup d'autres voies signalétiques soient distinctes et plus complexes. Par exemple, lors de l'interaction entre les microglies et les astrocytes, les MV semblent délivrer des signaux transmembranaires dus à l'activation des récepteurs de surface [123]; dans d'autres cas, tels que la coagulation ou l'inflammation, les MV fonctionnent non seulement en tant que messagers, mais aussi comme des platesformes nécessaires au développement de fonctions cordonnées [98, 124]. De plus, les MV étant des structures signalétiques liées à la membrane, elles peuvent recevoir des signaux rétrogrades et les transférer aux cellules cibles [109, 125]. Les MV ont été détectées dans plusieurs types de cancer. Elles sont souvent riches en facteurs tissulaires, permettant d’expliquer l’augmentation du risque de problèmes thrombotiques chez ces patients [126]. Il a aussi été démontré que des MV provenant de cellules cancéreuses étaient en charge de transporter le récepteur de l'EGF aux CE et contribuaient ainsi à la stimulation de l'angiogénèse [127]. En raison de leurs nombreuses propriétés, les MV sont engagées dans le contrôle de nombreux processus importants pour l'homéostasie physiologique ou pathologique des populations cellulaires.
1-5-1-5 Définition des exosomes
Les Exo sont des vésicules de diamètre de 30-100nm sécrétées par les cellules. Ils contiennent des protéines, de l'ADN, de l'ARN, des peptides et des dérivés lipidiques, entourés par une membrane bicouche lipidique. Ils proviennent des CMV qui fusionnent avec la membrane plasmique et libèrent leurs contenus par exocytose [128]. Il a été démontré que les Exo sont produits par une variété de types cellulaires incluant les cellules T [129], les mastocytes, les cellules dendritiques, les cellules tumorales, les cellules souches mésenchymateuses et les CE. Les Exo sont actuellement
considérés comme des médiateurs intercellulaires transportant la signalisation cellulaire agissant de façon indépendante mais en synergie avec les facteurs de croissance solubles. [130].
La méthode la plus commune pour isoler les Exo est l'ultracentrifugation à 100 000 à 120 000 × g [128, 129]. Cependant, l'inconvénient de cette méthode est son inefficacité à séparer les sous-types de EV l'un de l'autre [129]. Pour éviter le co-isolement des autres types de EV, une série d'étapes de centrifugation différentielle finissant avec l'ultracentrifugation et l'isolement des exo est généralement effectuée. Une centrifugation différentielle entre 10 000 et 20 000 x g précédant l'ultracentrifugation différentielle est capable d'éliminer les plus grosses VE tels que les MV et les CMV. Une filtration avec des pores de 0,1 ou 0,2 μm est également efficace. L'ultrafiltration et la microfiltration ont été utilisées pour isoler les Exo de l'urine [131, 132]. Plusieurs études ont mis en évidence le fait que l'ultracentrifugation peut être utilisée en conjonction avec d'autres méthodes d'isolement, y compris la centrifugation par gradient de densité (saccharose, saccharose-oxyde de deutérium (D2O) et OptiPrep ™ (iodixanol) [133], les méthodes d'immunoaffinité (immunoprécipitation avec des billes magnétiques recouvertes d’anticorps [134], et la cytométrie en flux [135].
1-5-1-6 Formation des exosomes
Le processus de formation des CMV et des Exo est coordonné par le complexe de criblage endosomal nécessaire pour le transport (ESCRT : endosomal sorting complexes required for transport). Ce complexe multiprotéique se compose de quatre ensembles de protéines solubles appelés ESCRT-0, ESCRT-I, ESCRT-II et ESCRT-III. Les complexes ESCRT recrutent des protéines ubiquitinylées et s’assemblent de manière définie à la membrane des endosomes. Ils entrainent la formation de bourgeonnements de tailles comparables qui sont clivés sous forme de vésicules intraluminales dans la lumière des endosomes appelées CMV [136]. Les connaissances actuelles de la façon dont ces CMV fusionnent avec la membrane plasmique pour libérer les Exo dans le milieu extracellulaire sont préliminaires. Cependant, la production et la libération des Exo dépendent de l'activité des différents Rab-GTPases et de leurs effecteurs, connus pour réguler le transport vésiculaire entre des organites spécifiques [137].
1-5-1-7 La composition des exosomes
Les Exo contiennent des ARNm et des microARN (miARN) qui peuvent être transférés entre les cellules et modulent l'expression génique des cellules réceptrices [138, 139]. Le développement d'ExoCarta, une base de données manuelle qui répertorie les protéines, l'ARN et les lipides identifiés dans les Exo [140, 141], et Vesiclepedia, un compendium d'annotation communautaire pour toutes les EV [142], ont permis aux chercheurs de fournir avec succès un aperçu de leur composition moléculaire [140]. Ces deux bases de données ont révélé que les protéines des Exo sont mieux étudiées que celles des MV. Similaire aux MV, les Exo expriment des protéines uniques à eux même, ainsi que des protéines spécifiques de leur cellule parente [79, 143]. De plus, la composition de leur bicouche lipidique diffère de la composition lipidique de la membrane plasmique de leur cellule d'origine[144, 145].
Le contenu luminal des Exo contient principalement des protéines cytosoliques dérivées de la cellule mère [79]. D'autres protéines souvent rencontrées sont liés au système endosomique (par exemple, différents Rab-GTPases ou Tsg101 du complexe ESCRT I, Alix), au niveau du cytosquelette (par exemple β-actine, moésine, cofillin et tubuline), à l'organisation des microdomaines dans les radeaux lipidiques (par exemple, la tétraspanines CD9, CD63, CD81 et CD82 ou les flotillins FLOT1 et FLOT2), à l'adhésion (par exemple les intégrines). De façon compatible avec leur fonction proposée comme vésicules présentatrice d’antigène [129], de nombreux Exo contiennent des protéines du CMH de classe I et II. Ils contiennent des protéines de choc thermique tels que HSP70 et HSP90 et sont également enrichis en certains lipides: principalement en céramide, en cholestérol, en Phosphatydilsérine et en sphingolipides [146]. De plus, ils comtiennent des protéines impliquées dans les voies de signalisation cellulaires comme β-caténine, WNT5B ou Notch Delta-like ligand 4 [147]. Les Exo ont également des signatures polysaccharidiques et glycérique sur leur surface externe, comprenant principalement du mannose, des acides α-2,3 et α-2,6-sialiques, des glycanes N-liés complexes et de la polylactosamine [148, 149].