Effet des engrais verts et de leur période
d’enfouissement sur la nutrition azotée et les
rendements du blé
Mémoire
Frédéric Verville
Maîtrise en biologie végétale
Maître ès sciences (M. Sc.)
Québec, Canada
iii
Résumé
L’objectif de ce projet était de déterminer l’effet de cinq engrais verts (mélange trèfle rouge/trèfle blanc, pois fourrager, moutarde blanche, radis huileux, avoine) et de leur période d’enfouissement à l’automne ou au printemps sur la nutrition azotée et les rendements d’une culture subséquente de blé sous les conditions du Québec. L’expérience a été répétée à deux reprises sur trois années (2011-2012 et 2012-2013) à deux stations expérimentales. Lors de tous les essais, l’engrais vert de trèfle rouge et trèfle blanc implanté en intercalaire d’une culture d’orge a été celui qui a produit le plus de biomasse, qui a accumulé le plus d’azote dans ses tissus et qui a permis d’augmenter significativement le rendement de la culture de blé subséquente. Le traitement d’enfouissement a eu peu d’effets significatifs et ces effets ne semblent pas liés à une influence sur la minéralisation des résidus d’engrais verts.
v
Table des matières
Résumé ... iii
Table des matières ... v
Liste des tableaux ... vii
Liste des figures ... ix
Remerciements ... xi
Avant-propos ... xiii
1. Introduction ... 1
2. Revue de littérature ... 3
2.1 L’azote dans les sols ... 3
2.2 Définition et mise en contexte de l’utilisation des engrais verts ... 5
2.3 Avantages et défis des engrais verts ... 6
2.4 Les engrais verts comme outil de gestion de l’azote : éléments à prendre en considération ... 9
2.4.1 Rendements et composantes de l’engrais vert ... 9
2.4.1.1 Effets des espèces choisies ... 9
2.4.1.2 Effets de l’environnement... 13
2.4.1.3 Effets des pratiques culturales ... 15
2.4.2 Minéralisation de l’azote dans les sols ... 19
2.4.2.1 Effets de l’engrais vert ... 19
2.4.2.2 Effets de l’environnement... 21
2.4.2.3 Effets des pratiques culturales ... 22
2.5 Prélèvement de l’azote par la culture suivante ... 24
2.5.1 Effets de l’utilisation des engrais verts sur les rendements de la culture suivante : quelques exemples issus de la littérature ... 24
2.5.2 Valeur fertilisante associée à l’engrais vert ... 33
3. Hypothèses et objectifs ... 37
4. Effet des engrais verts et de leur période d’enfouissement sur la nutrition azotée et les rendements du blé ... 41
Résumé ... 41
Introduction ... 42
Matériels et Méthodes ... 44
Description des sites et des traitements ... 44
Collecte de données ... 46
Analyses des échantillons de végétaux et de sols ... 47
Analyses statistiques ... 48
Résultats et discussion ... 48
Caractéristiques et biomasses des engrais verts ... 49
Rendements et prélèvements en azote du blé ... 51
Azote des sols ... 54
Conclusion ... 56
Remerciements ... 57
Références ... 57
5. Discussion générale ... 69
5.1 Effets des traitements sur les caractéristiques et les rendements des engrais verts ... 69
5.2 Effets des traitements sur les rendements du blé ... 73
5.2.1 Effets du traitement d’engrais vert ... 73
5.2.2 Effets du traitement d’enfouissement ... 76
5.3 Effets des traitements sur l’azote des sols ... 78
6. Conclusion générale ... 81
Bibliographie ... 83
Annexe A – Schéma du dispositif expérimental ... 95
vii
Liste des tableaux
Tableau 1: Caractéristiques des principaux engrais verts de climat tempéré : revue de différentes études. ... 36 Tableau 2: Dates des principales opérations culturales et d’échantillonnages lors des deux
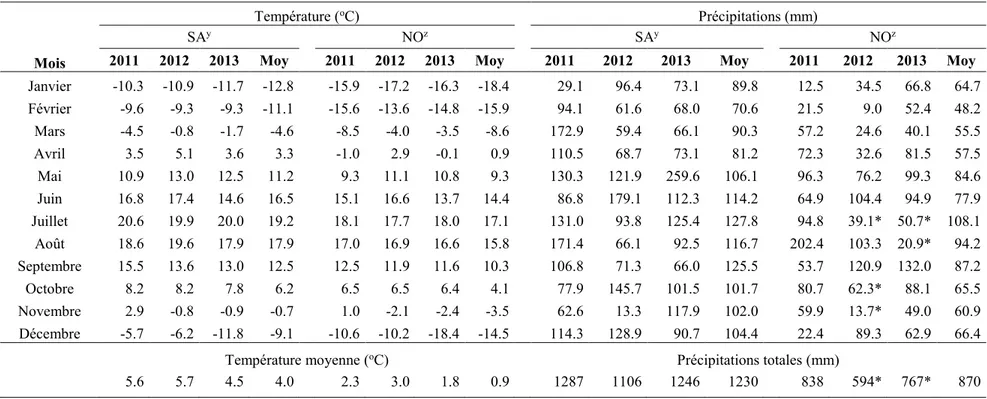
essais (2011-2012; 2012-2013) aux sites de Saint-Augustin (SA) et de Normandin (NO). ... 61 Tableau 3: Température et précipitations mensuelles lors des trois années d'essais pour les
sites de Saint-Augustin (SA) et de Normandin (NO) et température moyenne (Moy) pour les années 1971-2000. ... 62 Tableau 4: Concentrations en azote (N), en carbone (C) et rapport C/N de la biomasse
aérienne des engrais verts (EV) pour les sites de Saint-Augustin (SA) et de Normandin (NO) lors des deux années d'essais (2011-2012; 2012-2013). ... 63 Tableau 5: Rendements et accumulation en azote (N) de la biomasse aérienne des engrais
verts (EV) pour les sites de Saint-Augustin (SA) et de Normandin (NO) lors des deux années d'essais (2011-2012; 2012-2013). ... 64 Tableau 6: Rendements en grains et en paille de blé selon les traitements d'enfouissement
(ENF) et d’engrais verts (EV) pour les sites de Saint-Augustin (SA) et de Normandin (NO) lors des deux années d'essais (2011-2012; 2012-2013). ... 65 Tableau 7: Prélèvements en N retrouvé dans les grains et la paille de blé selon les
traitements d'enfouissement (ENF) et d’engrais verts (EV) pour les sites de Saint-Augustin (SA) et de Normandin (NO) lors des deux années d'essais (2011-2012; 2012-2013). ... 66 Tableau 8: Teneur en azote minéral (NO3-N + NH4-N) dans les sols (0-20 cm) selon les
traitements d'enfouissement (ENF) et d’engrais verts (EV) lors des quatre échantillonnages réalisés durant les deux années d'essais (2011-2012; 2012-2013) au site de Saint-Augustin. ... 67 Tableau 9: Teneur en azote minéral (NO3-N + NH4-N) dans les sols (0-20 cm) selon les
traitements d'enfouissement (ENF) et d’engrais verts (EV) lors des quatre échantillonnages réalisés durant les deux années d'essais (2011-2012; 2012-2013) au site de Normandin ... 68
ix
Liste des figures
xi
Remerciements
Je voulais tout d’abord remercier ma directrice, Anne Vanasse, la principale responsable de mon choix de prendre en main ce projet de maitrise. Sans sa bonne humeur contagieuse et son énergie débordante, je ne crois pas que j’aurais trouvé la motivation de me lancer dans cette aventure. Merci Anne de m’avoir fait confiance et de m’avoir laissé toute cette liberté au cours des deux dernières années, j’espère avoir été à la hauteur de cette confiance et ne pas avoir été une source de stress trop grande. Finalement, merci d’avoir partagé avec moi tes connaissances, ton sens critique et ta passion, je suis convaincu que ce bagage que tu m’as donné m’accompagnera longtemps et fera de moi une meilleure personne. Merci aussi à Martin Chantigny, qui m’a codirigé durant ces deux années. Merci Martin pour tes conseils, tes commentaires, ta disponibilité, ton efficacité et ton support. Tu as aussi su faire preuve de confiance et de patience et je l’apprécie beaucoup. J’espère aussi avoir été à la hauteur de tes attentes dans ce projet.
Ce projet n’aurait pu aboutir sans le soutien de nombreuses autres personnes qui m’ont épaulé tout au long du chemin. Je tiens à remercier tout particulièrement Marie-Ève Bernard qui, par sa rigueur, son sens de l’organisation et sa grande gentillesse, a su remplacer à de maintes reprises plusieurs parties de mon corps : mes jambes, mes bras, mais particulièrement ma tête. Merci Marie-Ève pour ta bonne humeur, pour ton aide précieuse et intarissable, je crois sincèrement que je n’y serais pas parvenu sans toi. Merci aussi à Valérie Bélanger, Annie Brégard, Francis Gagnon ainsi qu’à toutes et tous mes collègues avec qui j’ai travaillé à la ferme, vous avez été d’un grand soutien et d’une très agréable compagnie, vous me manquez toutes et tous beaucoup.
Merci à toute l’équipe d’Agriculture et Agroalimentaire Canada à Québec. Merci tout particulièrement à Annie Robichaud, Johanne Tremblay, Gabriel Lévesque et Alain Larouche pour leur accompagnement et leur assistance plus que nécessaire dans le laboratoire et sur le terrain. Merci à l’équipe d’Agriculture et Agroalimentaire Canada à Normandin, et plus particulièrement à Denis Pageau et Isabelle Morasse, dont l’efficacité a
été exemplaire tout au long de ce projet. Merci à l’équipe du CÉROM à Saint-Mathieu de Beloeil. Merci plus particulièrement à Gilles Tremblay, Stéphane Beaulieu et Julie Guérin. Finalement, merci à ma famille et à mes ami-es qui m’ont soutenu et accompagné tout au long de ces deux années. Merci à ma mère Lise, à mon père Marcel et à ma deuxième mère Marcelle d’avoir fait de moi la personne que je suis. Vous avez toujours été là pour moi et je vous en remercie beaucoup. Merci aussi à Sébastien, Jean-François, Marilyn et Julie, et un merci tout spécial à mes neveux Samuel et Antoine, ces adorables petits garçonnets, précieuses petites sources de distractions lors de ces longs moments où j’essayais de travailler. Merci tout le monde…!
xiii
Avant-propos
Ce mémoire compte six chapitres. Le premier chapitre consiste en une brève introduction du sujet. Le chapitre 2 rassemble les principales connaissances reliées aux facteurs qui vont influencer la production de biomasse par les engrais verts et la minéralisation des résidus suite à leur destruction. Il traite aussi des résultats de plusieurs recherches qui ont étudié les effets des engrais verts sur les rendements d’une culture subséquente. Le chapitre 3 énonce les hypothèses de recherche qui ont été formulées suite à la revue bibliographique en plus de faire mention des objectifs spécifiques du projet. Le chapitre 4 traite des principaux résultats de la recherche et est écrit sous la forme d’un article scientifique qui sera éventuellement traduit et soumis à une revue scientifique. L’auteur principal est Frédéric Verville, appuyé dans la rédaction par les coauteurs Anne Vanasse, Martin Chantigny, Denis Pageau et Gilles Tremblay. Le chapitre 5 correspond à un retour sur les hypothèses de recherche, qui seront davantage discutées à la lumière des résultats obtenus, alors que le chapitre 6 conclut de manière générale le document.
Les résultats du projet ont été diffusés sous forme de présentations orales lors de la Journée de la recherche de la Faculté des sciences de l’agriculture et de l’alimentation (FSAA) en novembre 2013, lors de la Journée d’information scientifique – Grandes cultures du Centre de références en agriculture et agroalimentaire du Québec (CRAAQ) en février 2014 et lors du congrès annuel de l’Association québécoise des spécialistes en sciences du sol (AQSSS) en mai 2014. Le financement du projet a été assuré par les bourses du Conseil de recherches en sciences naturelles et génie du Canada (CRSNG), du Fonds de recherche du Québec – Nature et technologie (FQRNT) et le financement du Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ), envers lesquels nous sommes reconnaissants.
1
1. Introduction
Plusieurs légumineuses, graminées et crucifères sont utilisées dans les systèmes culturaux comme engrais verts afin d’améliorer les propriétés chimiques, physiques et biologiques du sol (Fagaria et al., 2005), d’augmenter l’efficacité d’utilisation de l’azote (Thorup-Kristensen et al., 2003) et d’atténuer les émissions de gaz à effet de serre (Robertson et al., 2000). Ces engrais verts permettent aussi de réduire l’érosion des sols et de rompre le cycle des maladies, des insectes ravageurs et des plantes adventices (Reeves, 1994). Pour toutes ces raisons, l’utilisation des engrais verts dans les rotations peut contribuer à maintenir ou à augmenter les rendements des cultures tout en réduisant les apports en intrants externes (Tonitto et al., 2006).
Les engrais verts ont la possibilité de produire les mêmes rendements que les fertilisants azotés conventionnels à condition que ceux-ci accumulent suffisamment d’azote dans leurs tissus et que cet azote soit prélevé par la culture subséquente suite à une bonne synchronisation de sa minéralisation avec les besoins de la culture (Tonitto et al., 2006). Cependant, plusieurs facteurs vont gouverner ce processus. Il existe des différences fondamentales entre les types de plantes utilisées comme engrais verts qui vont influencer leur accumulation d’azote et leur minéralisation. Par exemple, les légumineuses sont en mesure de fixer l’azote de l’air alors que les non-légumineuses ne le peuvent pas. Les légumineuses ont généralement un rapport C/N bas, qui mènera à une minéralisation rapide des résidus, alors que les graminées ont à maturité un rapport C/N plus élevé, ce qui risque de causer une immobilisation temporaire de l’azote (Reeves, 1994).
L’environnement et les pratiques culturales vont également conditionner l’utilisation des engrais verts. Dans certaines régions plus tempérées, les engrais verts survivront à l’hiver alors que dans d’autres, plus nordiques, ils seront détruits. Dans les régions plus arides, il peut être avantageux d’enfouir les engrais verts plus rapidement à l’été (Foster, 1990) alors que dans les régions nordiques plus humides et où l’hiver apporte un couvert de neige, il semble préférable de les enfouir le plus tard possible à l’automne pour éviter les pertes sans nuire aux rendements de la culture subséquente (Lahti et Kuikman, 2003).
Étant donné ces nombreux facteurs, il devient primordial de mener des études sur les engrais verts dans un grand nombre de contextes climatiques et agronomiques différents. Il existe beaucoup de résultats de recherche sur les engrais verts, mais très peu sur les engrais verts utilisés en intercalaire ou à la dérobée d’une culture dans le contexte climatique québécois. Est-ce possible de cultiver efficacement des engrais verts au Québec, sans pour autant devoir se priver d’une récolte? Le présent mémoire a pour objectif de tenter de répondre à cette question.
3
2. Revue de littérature
2.1 L’azote dans les sols
L’azote est l’élément le plus abondant dans l’atmosphère, le quatrième élément le plus abondant dans la biomasse des plantes et possiblement l’élément nutritif le plus important en agriculture (Paul et Clark, 1989). On isole souvent le cycle de l’azote dans les sols de son cycle global. Cependant, les deux cycles demeurent très liés puisque l’azote des sols tire malgré tout son origine de l’atmosphère (figure 1).
Figure 1: Cycle de l’azote
Source : Tiré de McKague et al., 2005
Il existe deux sources naturelles pour les apports d’azote dans les sols : la fixation biologique et l’apport d’ammoniac (NH3), de nitrate (NO3-) et de nitrite (NO2-) par les eaux
de pluie. Les pertes, toujours en milieu naturel, proviennent du lessivage et de la volatilisation (Stevenson et Cole, 1999). En milieu agricole, l’apport de fertilisants azotés
sous forme d’engrais chimique (fixation industrielle) ou d’engrais de ferme représente une nouvelle source, alors que le ruissellement des eaux de surface et l’exportation de biomasse végétale représentent de nouvelles pertes.
La fixation biologique de l’azote est réalisée par des cyanobactéries, des bactéries ou par des plantes qui vivent en association avec ces organismes et elle permet de convertir le diazote gazeux atmosphérique (N2) en NH3 ou sous forme d’azote organique (protéines,
acides nucléiques, etc.). Par la suite, le processus par lequel l’azote organique se dégrade en forme minérale, soit en ammonium (NH4+) puis en NO3-, se nomme minéralisation. Celle-ci
se déroule en deux étapes : l’ammonification est la conversion de la forme organique en NH4+ et la nitrification est l’oxydation du NH4+ en NO3-. Finalement, on parle
d’immobilisation lorsque ces deux éléments sont utilisés par les organismes du sol et les plantes (Stevenson et Cole, 1999).
Le NH4+ et le NO3- sont les deux principales formes d’azote prélevées par les plantes.
Cependant, ces deux éléments sont très mobiles et sont susceptibles d’être perdus s’ils ne sont pas rapidement réutilisés par les plantes. Les pertes en NH4+ surviennent par
volatilisation (par l’intermédiaire du NH3) et par ruissellement suite à une application
importante d’engrais azoté (McKague et al., 2005) alors que le NO3- est principalement
perdu par lessivage. Le NO3- peut également s’accumuler dans les sols et se transformer en
formes gazeuses volatilisables (protoxyde d’azote (N2O) et N2) par le biais de la
dénitrification (Paul et Clark, 1989). C’est donc pourquoi en agriculture, il est important de synchroniser au maximum la minéralisation de l’azote avec les besoins des cultures pour éviter les pertes inutiles.
Il est possible que l’azote inorganique formé suite à la minéralisation soit rapidement utilisé par la microflore hétérotrophe des sols. Si l’utilisation de l’azote inorganique est plus importante que sa production, on parlera d’immobilisation nette. Au contraire, si la production d’azote inorganique est plus importante que l’utilisation par les micro-organismes, on parlera de minéralisation nette. La quantité de carbone (C) présent dans la matière organique décomposée est un facteur majeur dans l’équation, puisque les microorganismes utilisent l’azote pour sa dégradation. Ainsi, Stevenson et Cole (1999)
5 considèrent que la décomposition de résidus ayant un rapport C/N de moins de 20:1 mènera à une minéralisation nette alors que la décomposition de résidus ayant un rapport C/N supérieur à 30:1 mènera à une immobilisation nette. Un rapport C/N entre 20 et 30 mènera à des échanges stables malgré le fait que les deux phénomènes puissent se produire à des taux significatifs.
2.2 Définition et mise en contexte de l’utilisation des engrais
verts
On définit les engrais verts comme étant une culture destinée à être enfouie lorsque verte ou peu de temps après la maturité avec comme objectif l’enrichissement des sols (SSSA, 1997). En comparaison, on définit les cultures de couverture, ou culture piège, comme des plantes cultivées dans le but de protéger les sols contre l’érosion et la perte d’éléments nutritifs (Reeves, 1994) ou encore comme des cultures servant à améliorer la qualité des sols, de l’eau et de l’air, le captage, le recyclage et la gestion des éléments nutritifs et/ou la lutte intégrée aux ennemis des cultures (Delgado et al., 2006). Dans tous les cas, il n’y a aucun débouché commercial direct à ces cultures (Fagaria et al., 2005). Les termes d’engrais verts et de cultures de couverture sont souvent utilisés comme étant des synonymes (Richards et al., 1996), d’autant plus que la culture peut remplir plusieurs rôles à la fois, c’est pourquoi il n’y aura pas de distinction de faite entre les deux termes dans le cadre du présent document.
Traditionnellement, la fertilité des sols agricoles reposait sur une combinaison de pratiques comme les rotations de cultures, la combinaison de l’élevage, des pâturages et des cultures annuelles ainsi que l’utilisation de cultures de couverture et d’engrais verts de légumineuses qui assuraient le rôle de fixateurs d’azote atmosphérique. Cependant, suite à la découverte du procédé Haber-Bosch, permettant la production d’azote synthétique, au début du 20e siècle et étant donné le contexte technicoéconomique qui prévalait à la fin de
la 2e guerre mondiale, ces pratiques ont graduellement été abandonnées à mesure que se
concrétisait la « Révolution verte » et ses méthodes culturales industrielles (Dinnes et al., 2002). Malgré les gains d’efficacité et de productivité indéniables associés à ces méthodes,
l’agriculture agrochimique a aussi apporté son lot de problèmes associés à une mécanisation toujours plus intense des travaux et à des pratiques monoculturales très dépendantes des produits chimiques de synthèse et des intrants extérieurs (Gold, 1999). Le retour des engrais verts dans les systèmes culturaux dynamiques représente une méthode pour augmenter la durabilité de l’agriculture, et ce, pour de nombreuses raisons (Tanaka et al., 2002).
2.3 Avantages et défis des engrais verts
Les engrais verts peuvent contribuer à augmenter la durabilité et la résilience de l’agriculture de différentes façons. Tout d’abord, en diversifiant la rotation des cultures, les engrais verts peuvent permettre de briser le cycle de certaines maladies et de certains ravageurs (pourriture racinaire, nématodes : Abawi et Widmer, 2000; punaise terne : Bugg et al., 1990) à condition d’agir comme une plante non-hôte. Ils peuvent également réduire la pression des mauvaises herbes en entrant en compétition avec elles pour la lumière, les éléments nutritifs ou l’eau (Martin, 1996; Liebman et Davis 2000; Bàrberi et Mazzoncini, 2001), en plus de réduire la pression des ravageurs en offrant des ressources ou des habitats pour les organismes bénéfiques (Bugg et Waddington, 1994; Lewis et al., 1997; Nicholls et Altieri, 2001). Les engrais verts représentent aussi une façon de lutter contre les changements climatiques en permettant de stocker davantage de carbone dans les sols et en diminuant l’utilisation de carburants fossiles (Robertson et al., 2000; Tonitto et al., 2006). Il faut par contre noter que les avantages énumérés précédemment, bien qu’ils ne soient pas négligeables, ne sont généralement pas ceux qui vont motiver les agriculteurs à adopter les engrais verts (Cherr et al., 2006). Les bénéfices qui retiennent le plus l’attention concernent davantage leurs effets bénéfiques sur les sols et la possibilité de mieux gérer et conserver les éléments nutritifs au champ.
Dans certains contextes, les engrais verts sont utilisés avant tout comme culture de couverture hivernale pour lutter contre l’érosion des sols, particulièrement dans les régions où ces derniers sont normalement laissés à nu durant l’hiver et où le couvert de neige est très faible, voire inexistant. Les cultures sarclées ont été identifiées comme étant particulièrement sensibles à l’érosion et à la dégradation des sols au Canada (McKeague et
7 al., 1987) et l’inclusion d’engrais verts dans la rotation, soit en intercalaire ou à la dérobé des cultures, peut permettre d’atténuer ces effets (Dapaah et Vyn, 1998). En réduisant l’érosion, ils peuvent aussi contribuer à diminuer les pertes d’éléments nutritifs et de pesticides dans l’environnement (Delgado et al., 2001; Gaston et al., 2003).
Les effets bénéfiques des engrais verts sur les propriétés physiques, chimiques et biologiques du sol sont également importants (Abdollahi et Munkholm, 2014). Tout d’abord, les engrais verts ont la capacité d’augmenter le contenu en carbone et en matière organique des sols. Puget et Drinkwater (2001) ont démontré que le carbone contenu dans les racines, plus lignifiées, peut persister plus longtemps dans les sols que celui des parties aériennes plus labiles, contribuant ainsi à la hausse du niveau de matière organique des sols. Par contre, cette hausse suite aux engrais verts n’est pas nécessairement élevée, représentant généralement moins de 1% (Utomo et al., 1990; Reddy et al., 2003). D’autres études démontrent aussi une amélioration de la stabilité des agrégats et de la porosité des sols (Roberson et al., 1991), malgré le fait que ces recherches n’arrivent pas à noter de différences significatives selon les différents types de racines des plantes, soit superficielles ou pivotantes (Carof et al., 2007), ou encore selon la quantité de biomasse des résidus (Biederbeck et al., 1998). Les engrais verts stimulent également l’activité enzymatique et biologique des sols de façon plus importante que les engrais chimiques (Kirchner et al., 1993; Abdallahi et N’Dayegamiye, 2000). Il faut cependant souligner que la plupart des recherches sur le sujet étudient les effets sur le court terme et que des études sur le long terme pourraient démontrer des effets plus significatifs ayant des impacts plus considérables (Cherr et al., 2006).
Un autre effet très recherché par les agriculteurs lorsqu’ils implantent un engrais vert est la réduction des pertes en éléments nutritifs des sols. Comparativement aux sols laissés à nu durant l’automne et l’hiver, les engrais verts ont le potentiel de réduire les pertes d’azote par lessivage et volatilisation en immobilisant l’azote disponible dans leurs tissus au fur et à mesure de leur croissance (Jackson et al., 1993; Staver et Brinsfield, 1998; Drury et al., 2014). Par contre, il n’y a pas nécessairement de lien entre la quantité d’azote accumulé dans la biomasse de l’engrais vert et la quantité de NO3 résiduel dans le sol. Il est possible
que le sol où se trouve une biomasse plus importante d’engrais vert contienne tout de même plus de nitrates potentiellement lessivables qu’un autre engrais vert moins productif (Baggs et al., 2000). À l’inverse, une biomasse d’engrais vert modeste n’est pas nécessairement synonyme d’inefficacité à prélever l’azote. Un rendement moyen peut être amplement suffisant pour capturer la majorité de l’azote présent (Baggs et al., 2000). Une fois cet azote capturé dans la plante, celui-ci peut ensuite être rendu disponible à une culture suivante par le biais de la minéralisation suite à son incorporation.
Finalement, le dernier avantage des engrais verts, et sans doute le plus important, est la possibilité d’introduire de l’azote dans le système agricole par le biais de la fixation biologique. Pour ce faire, les légumineuses sont particulièrement intéressantes puisqu’elles sont capables de fixer l’azote de l’air à l’aide de bactéries symbiotiques de type Rhizobium qui se retrouvent dans les nodules de leurs racines (Tonitto et al., 2006). Cet apport d’azote peut permettre de réduire les besoins de la culture suivante de façon non négligeable, représentant potentiellement une économie d’argent pour le producteur (Singh et al., 2004). Étant donné que la production des engrais chimiques de synthèse nécessite des quantités importantes de carburants fossiles, l’utilisation des légumineuses comme engrais vert représente une façon de réduire considérablement les besoins en carburant du secteur agricole (Tonitto et al., 2006). Cet apport d’azote par les engrais verts peut également représenter une solution aux problèmes liés à une utilisation excessive d’engrais de ferme comme source de fertilisant (particulièrement en agriculture biologique), soit la saturation du sol en phosphore ou la salinisation (Cherr et al., 2006). La quantité d’azote fixée par les légumineuses peut varier fortement selon plusieurs facteurs, comme il sera présenté plus loin.
Il peut également y avoir quelques problèmes avec l’utilisation des engrais verts. Leur incorporation peut mener à une immobilisation de l’azote lors de sa décomposition, ce qui peut nuire à la culture qui le suivra (Vyn et al., 1999). Sous certaines conditions, il est également possible que l’engrais vert prélève de trop grandes quantités d’eau et que cela vienne nuire à la culture qu’il accompagne ou celle qui suivra (Tollenaar et al., 1993; Thelen et al., 2004). Finalement, on peut également nommer l’augmentation du temps de
9 travail et des coûts, les possibilités d’interférences avec d’autres travaux au champ, les difficultés de destruction de l’engrais vert en plus des risques d’augmentation des ravageurs et des maladies (Dabney et al., 2001).
Pour arriver à maximiser les bénéfices liés à la gestion de l’azote tout en évitant d’avoir des problèmes, il devient donc primordial de bien connaître et comprendre les facteurs qui influencent la dynamique de l’azote dans le cycle de l’utilisation des engrais verts.
2.4 Les engrais verts comme outil de gestion de l’azote : éléments
à prendre en considération
2.4.1 Rendements et composantes de l’engrais vert
Comme cela a été soulevé précédemment, les engrais verts accumulent de l’azote dans leurs tissus dans le but de le rendre à la culture subséquente de deux façons : en capturant l’azote excédentaire des sols lors de périodes où ceux-ci auraient normalement été laissés à nu après une récolte (tous les types d’engrais verts) ou encore en fixant l’azote à partir de l’atmosphère (légumineuses). La quantité d’azote accumulée par ces deux phénomènes peut être influencée par plusieurs facteurs, dont le rendement en biomasse de l’engrais vert et sa composition, qui eux-mêmes peuvent varier en fonction de trois facteurs principaux : l’espèce utilisée, les pratiques culturales et l’environnement dans lequel l’espèce va croître (Badaruddin et Meyer, 1990).
2.4.1.1 Effets des espèces choisies
Le Tableau 1 présente les espèces qui ont été utilisées dans le cadre de ce projet ou encore d’autres espèces très communément utilisées comme engrais vert au Québec. Ce tableau permet de constater la très grande variabilité dans les rendements et les caractéristiques des différents engrais verts, non seulement entre les espèces, mais également au sein même d’une espèce, étant donné les différentes conditions dans lesquelles ces recherches ont été menées. Les facteurs qui vont influencer les rendements sont majoritairement d’ordre climatique et technique, des aspects qui seront abordés plus tard dans la section 2.4.1.2 et 2.4.1.3.
Le Tableau 1 présente également la teneur en azote de différents engrais verts; teneur en N qui s’explique généralement par le type d’engrais vert, mais également par le stade phénologique de la plante au moment de l’échantillonnage (abordé à la section 2.4.1.3). Pour le type d’engrais vert, on remarque, avec les données du Tableau 1, que les engrais verts de non-légumineuses (graminées et crucifères) possèdent généralement une proportion d’azote plus faible dans leurs tissus que les engrais verts de légumineuses avec respectivement environ 1.5 à 2.0 % de N en moyenne par rapport à 2.5 à 3.5 % et même plus. Cela veut dire que, pour une même biomasse d’engrais vert, les légumineuses peuvent contenir facilement le double d’azote, ce qui les rend beaucoup plus efficaces dans leur accumulation et, potentiellement, dans leur transfert à une culture subséquente. Il est aussi important de noter que, dans le cas des légumineuses, une grande partie de cet azote aura été fixé de l’atmosphère, ce qui représente un gain par rapport aux non-légumineuses. L’azote contenu dans la plante provient en partie (légumineuses) ou totalement (graminées et crucifères) du sol. L’accumulation en N dans les résidus de non-légumineuses est donc dépendante de la disponibilité en azote du sol, qui elle est dépendante de la quantité d’azote résiduel et du taux de minéralisation de l’azote dans les sols (Wagger et Mengel, 1988). L’azote résiduel représente le N minéral qui reste dans les sols suite à une culture. Cet azote excédentaire, à risque de lessivage et de volatilisation, est capturé dans la biomasse et mis à l’abri des pertes lorsqu’un engrais vert est utilisé. Même si les non-légumineuses accumulent moins d’azote dans leur biomasse que les non-légumineuses, 100% de l’azote qu’elles prélèvent provient de l’azote résiduel et des réserves du sol. C’est pourquoi, en général, on reconnait que les graminées et les crucifères sont les plantes les plus efficaces pour récupérer l’azote du sol, même si les légumineuses sont également en mesure d’en récupérer une certaine partie (Thorup-Kristensen et al., 2003; Snapp et al., 2005). Dans une méta-analyse sur le sujet, Meisinger et al. (1991) compilent des résultats qui nous permettent de conclure qu’en moyenne, les graminées, les crucifères et les légumineuses réduisent le lessivage de l’azote de respectivement 61 %, 62 % et 25 %. Dans une méta-analyse plus récente, Tonitto et al. (2006) tirent sensiblement les mêmes conclusions en évaluant que la capture moyenne de l’azote excédentaire post-récolte par les non-légumineuses varie de 20 à 60 kg N ha-1, ce qui est cohérent avec le Tableau 1 et qui
11 représente une réduction du lessivage de 40 à 70 % comparativement à un sol laissé à nu. Cependant, au sein même de ces familles, il est aussi normal de voir des variations en terme de biomasses et de prélèvements d’azote puisque toutes les plantes ne vont pas croître ni prélever les éléments de la même façon. Par exemple, les céréales de printemps produisent généralement plus de biomasse aérienne que les céréales d’automne lorsqu’utilisées à la dérobé suite à une récolte puisque ces dernières stockent des réserves dans leurs racines en prévision de survivre à l’hiver. À titre d’exemple, Andraski et Bundy (2005) ont montré que l’avoine a produit significativement plus de biomasse aérienne qu’un triticale et un seigle d’automne lors de deux années sur trois. Dans une étude comparant plusieurs types de trèfles, den Hollander et al. (2007) ont démontré que dans des conditions semblables, un trèfle rouge et un trèfle incarnat allaient produire davantage de biomasse qu’un trèfle blanc, mais que la concentration en N du trèfle blanc, plus élevée que celle du trèfle incarnat, permettait malgré tout une accumulation d’azote équivalente à ce dernier (Tableau 1). Reeves (1994) avance également que le trèfle incarnat produit une biomasse élevée plus rapidement que la vesce velue, une affirmation qui peut partiellement se confirmer par les résultats présentés dans le Tableau 1, où on remarque que le trèfle incarnat peut arriver à produire une biomasse de 5 t MS ha-1 en l’espace de seulement deux mois (Dyck et al.,
1995).
Contrairement au prélèvement d’azote dans les sols, qui est à la portée de tous les types de plantes, les légumineuses sont les seules plantes herbacées qui peuvent réaliser la fixation biologique de l’azote dans les zones tempérées (Smith et al., 1987). La quantité d’azote fixée et stockée dans la biomasse va varier selon l’espèce choisie, mais aussi selon l’azote résiduel dans les sols, les conditions climatiques et les méthodes culturales (Reeves, 1994), critères qui seront abordés un peu plus loin aux sections 2.4.1.2 et 2.4.1.3. À titre d’exemple concernant la variabilité entre les espèces, Karpenstein-Machan et Stuelpnagel (2000) ont comparé les rendements en azote fixé par un pois fourrager et un trèfle incarnat poussant dans les mêmes conditions en culture pure durant un été complet et ont obtenu respectivement des résultats de 178 et 242 kg N ha-1. Deux études font la revue de résultats
les espèces et au sein même d’une espèce allant de 25 à 300 kg N ha-1, malgré que la
moyenne semble plutôt se situer entre 60 et 150 kg N ha-1 (Phillips, 1980 ; Fagaria et al.,
2005).
Un autre aspect qui semble important, généralement moins étudié étant donné les difficultés inhérentes à l’échantillonnage, sont les racines des plantes qui servent d’engrais verts. Ici encore, on trouve une très grande variabilité dans le profil et la biomasse racinaire des différentes espèces, ce qui peut avoir un impact sur leur prélèvement, mais aussi leur rendement en azote pour la culture suivante. Reeves (1994) a recensé plusieurs profils racinaires de plantes cultivées ou utilisées comme engrais verts et a noté des variations en profondeur allant de 90 centimètres pour le pois à 6 mètres pour la luzerne, avec des variations latérales allant de 15 centimètres pour la luzerne et le blé à 1 mètre pour le maïs. Dans une étude de trois ans portant sur les racines, Griffin et al. (2000) ont rapporté que les racines de la luzerne, du seigle et d’un mélange de vesce velue et de seigle pouvaient représenter respectivement 56, 46 et 38% de la biomasse totale de la plante alors que ces mêmes racines contenaient 32, 28 et 19% de l’azote total accumulé par l’engrais vert. Kirchmann (1988) a également comparé la proportion d’azote contenue dans la portion racinaire chez six espèces de trèfle et a remarqué des variations assez importantes allant de 3 % pour le trèfle de Perse à 45 % pour le trèfle blanc. Finalement, Ramos et al. (2001) ont aussi mesuré une proportion de 39 et 49 % de l’azote total contenu dans les racines de deux espèces de légumineuses, respectivement Canavalia ensiformis et Mucuna aterrima, mais ont aussi mesuré que 10 et 12 % de tout l’azote accumulé par ces deux mêmes espèces a été directement transféré au sol par les racines, le «turnover» des nodules et les exsudats racinaires.
Un dernier point qui apparait dans le Tableau 1, qui va beaucoup varier selon le type d’engrais vert et qui est très important lorsque vient le temps de gérer l’azote au champ, est le rapport C/N de l’engrais vert. On peut noter que les légumineuses ont tendance à avoir un rapport C/N plus faible que les graminées et les crucifères étant donné, entre autres, que le rapport est fortement influencé par la teneur en N de l’engrais vert. Le rapport C/N est aussi beaucoup influencé par le stade de croissance de la plante au moment de sa
13 destruction, un facteur qui dépend beaucoup des pratiques culturales, abordées à la section 2.4.1.3. Reeves (1994) a fait le recensement de plusieurs études et indique que de façon générale, le rapport C/N des graminées se situera au-dessus de 30:1 alors que celui des légumineuses sera généralement sous la barre des 20:1, ce qui semble cohérent avec les données du Tableau 1 et ce qui aura un impact important sur la minéralisation de l’azote provenant de ces biomasses (section 2.4.2). Outre l’impact sur la minéralisation de l’azote, le rapport C/N aura aussi un impact sur les qualités du sol. Selon Hadas et al. (1998), les résidus d’engrais verts plus murs et donc avec un rapport C/N plus élevé vont favoriser la stabilité des agrégats par la formation de polysaccharides dans les sols, en plus d’être utilisés comme source d’énergie microbienne et favoriser la formation d’humus plus stable et la hausse de la matière organique des sols. À l’opposé, les résidus jeunes, fermentescibles et avec un rapport C/N plus faible vont stimuler l’activité de la microflore et la formation des agrégats pour une courte durée.
Ainsi, tous ces éléments (capacité à capturer l’azote résiduel, fixation de l’azote par les légumineuses, biomasse aérienne et racinaire, concentration en azote, rapport C/N) sont en partie influencés par l’espèce d’engrais vert qui sera utilisée. C’est donc pourquoi il est important de choisir ses engrais verts selon ses besoins et les objectifs fixés. Cependant, comme cela a été souligné précédemment, de nombreux autres facteurs vont aussi affecter ces éléments et les performances des engrais verts, soit ceux liés à l’environnement et aux pratiques culturales.
2.4.1.2 Effets de l’environnement
Il est très important de sélectionner des espèces d’engrais vert adaptées aux conditions du milieu où elles vont croître si on souhaite maximiser leur production, car le climat est possiblement le facteur qui limite le plus la croissance des engrais verts (Cherr et al., 2006). Les variations de biomasses importantes pour une même étude présentée dans le Tableau 1 dépendent généralement des conditions climatiques. Par exemple, dans Andraski et Bundy (2005), les rendements les plus élevés pour l’avoine (3.0 t MS ha-1 vs 0.6 t MS ha-1) ont été
obtenus durant l’année 2000, lorsque les températures en septembre et octobre ont été particulièrement chaudes. Toujours dans la même étude, les rendements les plus élevés de
seigle d’automne (1.4 t MS ha-1 vs 0.6 t MS ha-1) ont été obtenus en 2001, lorsque le mois
d’avril fut plus chaud. Dans Ranells et Wagger (1996), l’automne de l’année 1994 fut particulièrement chaud et humide, ce qui a contribué à augmenter de presque quatre fois les rendements du trèfle incarnat (5.0 t MS ha-1 vs 1.4 t MS ha-1) et du seigle d’automne (5.7 t
MS ha-1 vs 1.5 t MS ha-1) (Tableau 1). Les rendements de la vesce velue, dans les travaux
de Gabriel et Quemada (2011), menés dans un climat méditerranéen plutôt sec, ont été particulièrement influencés par les précipitations abondantes suite au semis de l’engrais vert durant l’année 2006, ce qui a mené à un rendement pratiquement dix fois plus élevé que l’année suivante (5.0 t MS ha-1 vs 0.6 t MS ha-1).
De façon générale, plus le climat est chaud et humide, plus les options d’engrais verts et leurs bénéfices potentiels sont élevés (Dabney et al., 2001). Les engrais verts vont normalement se classer en deux catégories : les espèces tropicales et les espèces tempérées. Les espèces tropicales vont tolérer des températures très élevées (> 35°C) mais vont être particulièrement sensibles au gel, alors que les espèces tempérées vont généralement moins bien performer à des températures élevées (> 30°C), mais vont, dans certains cas, résister à des températures inférieures au point de congélation (jusqu’à -10°C) (Cherr et al., 2006). Ainsi, dans nos climats nordiques, on ne choisira pas les mêmes espèces pour un engrais vert d’été et un engrais vert d’automne, puisque les engrais verts d’automne requièrent de résister davantage au froid. Le seigle d’automne est reconnu comme étant l’engrais vert de zone tempérée le plus résistant au froid, suivi par la vesce velue dans la catégorie des légumineuses (Dabney et al., 2001). C’est pourquoi, comme on peut le voir dans le Tableau 1, ces engrais verts sont généralement utilisés durant plusieurs mois comme engrais vert d’hiver, alors qu’on utilisera surtout l’avoine, le radis, la moutarde et le trèfle incarnat comme engrais vert de courte durée. La vesce velue serait plus résistante au froid que le trèfle incarnat, qui n’est pas conseillé d’utilisation sous les -18oC (Reeves, 1994). La
moutarde, le trèfle blanc et le raygrass seraient également identifiés comme des engrais verts peu résistants au gel à l’automne (Shepherd et May, 1992; Miller et al., 1994).
Certaines espèces vont également mieux performer en sol plus sec ou en sol plus humide. Par exemple, la luzerne semble mieux performer que le trèfle rouge en période de
15 sécheresse (Peterson et al., 1992). Dans les climats semi-arides, l’utilisation d’espèces qui consomment moins d’eau aura aussi l’avantage de ne pas épuiser les sols pour la culture suivante. En Saskatchewan, le pois et la gesse commune ont utilisé l’eau plus efficacement que la lentille ou la gesse de Tanger, malgré qu’en général, ces engrais verts consomment les mêmes quantités d’eau qu’un blé et davantage qu’une jachère (Townley-Smith et al., 1993; Biederbeck et Bouman, 1994). Dans les climats plus arides, on aura aussi avantage à opter pour des espèces à grosses graines, qui peuvent être semées plus en profondeur, afin d’augmenter les chances de germination en cas de sécheresse prolongée (Keeling et al., 1996). Les stress hydriques peuvent également réduire de façon significative la fixation biologique de l’azote (Gardner et al., 1985).
Finalement, l’azote résiduel présent dans le sol est un dernier facteur environnemental qui pourrait influencer les performances des engrais verts. Un niveau d’azote résiduel élevé dans le sol aura différents impacts. D’abord, il permettra une meilleure croissance des plantes qui ne fixent pas l’azote de l’air telles les graminées et les crucifères (Möller et al., 2008), alors que la réponse des légumineuses sera faible ou nulle (Cline et Silvernail, 2001). De plus, ce niveau élevé d’azote résiduel va diminuer la capacité des légumineuses à fixer l’azote de l’air étant donné que la fixation biologique est stimulée lorsque la légumineuse n’est pas en mesure de prélever suffisamment d’azote en provenance du sol, soit par absence ou par compétition avec d’autres plantes (Gardner et al., 1985; Karpenstein-Machan et Stuelpnagel, 2000). Ainsi, si on fait pousser une légumineuse sur un sol riche, on perd l’avantage de la fixation biologique tout en augmentant les chances de pertes d’azote plus élevées par le biais du lessivage ou de la volatilisation.
2.4.1.3 Effets des pratiques culturales
Plusieurs pratiques culturales vont influencer la croissance et le rendement des engrais verts. On peut entre autres identifier la régie d’implantation et de fertilisation ainsi que le calendrier de culture de l’engrais vert, du semis à l’enfouissement.
Il a été mentionné précédemment que les graminées et les crucifères vont réduire les pertes d’azote et que les légumineuses vont fixer l’azote atmosphérique pour l’intégrer dans leurs
tissus. Ainsi, il est reconnu que la mise en place d’un mélange d’engrais verts incluant des graminées/crucifères avec des légumineuses permettrait de profiter de ces deux avantages conjugués en plus de fournir une biomasse supérieure et une plus grande stabilité dans les rendements par rapport à ce qu’on obtiendrait normalement avec la monoculture de chaque espèce (Ranells et Wagger, 1996; Karpenstein-Machan et Stuelpnagel, 2000; Sainju et al., 2005). Toutefois, un mélange n’est pas nécessairement synonyme de bénéfices supérieurs à une culture pure sur tous les plans (Smith et al., 2014). Karpenstein-Machan et Stuelpnagel (2000) ont comparé la culture hivernale du trèfle incarnat et du pois en culture pure et en mélange avec le seigle d’automne comme précédent d’un maïs l’été suivant. Lorsque les espèces étaient mélangées, les doses optimales de chacune en semis pur (seigle = 300 grains m-2, pois = 70 grains m-2, trèfle incarnat = 25 kg ha-1) étaient affectées par un ratio de
manière à ce que le total des deux doses de semis réduites donne l’équivalent d’une dose normale (ex : ¼ pois et ¾ seigle, ½ pois et ½ seigle, etc.). Ils ont utilisé le concept du ratio de surface équivalente (Land equivalent ratio, LER), qui est défini par la surface relative de terre nécessaire à des cultures pures pour produire le même rendement que ces cultures implantées en intercalaire ou en mélange (Mead et Willey, 1980). Par exemple, un LER de 1,5 signifie qu’il faudrait un equivalent de 1,5 hectare pour produire en monoculture de deux cultures ce qu’un seul hectare permet de produire lorsqu’on les cultive conjointement. Ils ont ainsi calculé un LER de 1,1 pour les rendements en biomasse de la combinaison de ½ pois - ½ seigle, de 1,2 pour les combinaisons de ¼ pois - ¾ seigle et ¾ pois - ¼ seigle et de 1,3 pour les trois combinaisons utilisant le trèfle incarnat et le seigle. Les légumineuses en mélange ont également fixé plus efficacement l’azote de l’air que les légumineuses en semis pur, malgré le fait qu’au total, une plus petite quantité d’azote ait été fixée étant donné la proportion plus faible de légumineuses dans les mélanges. La meilleure efficacité en mélange s’explique par le fait que le seigle s’est chargé de récupérer l’azote résiduel dans le sol, ce qui a obligé les légumineuses à fixer davantage d’azote pour subvenir à leurs besoins. Schipanski et Drinkwater (2011) arrivent à des conclusions semblables dans une étude comparant les rendements d’un trèfle établi au printemps en semis pur et au printemps en intercalaire d’une céréale d’automne récoltée. Le trèfle intercalaire a obtenu 72% de son azote par le biais de la fixation biologique comparativement à 63% pour le
17 trèfle en semis pur, pour un total d’azote fixé de 57 kg ha-1 pour le trèfle en intercalaire
contre 47 kg ha-1 en semis pur.
Les mélanges d’engrais verts peuvent avoir d’autres avantages ou encore des inconvénients. Une légumineuse grimpante mais sensible à la verse, comme le pois ou la vesce, profitera d’un mélange avec une faible dose de seigle qui offrira un support à la légumineuse, ce qui favorisera sa croissance en plus de stimuler la fixation d’azote (Karpenstein-Machan et Stuelpnagel, 2000). À l’inverse, une dose trop forte de seigle dans un mélange avec une légumineuse à petites graines et à établissement lent, comme le trèfle incarnat, risque d’étouffer cette dernière. Finalement, les mélanges peuvent également permettre de trouver un équilibre dans le rapport C/N global de notre engrais vert. La culture en mélange d’une légumineuse avec un rapport C/N bas et d’une graminée avec un rapport C/N élevé peut résulter en un rapport intermédiaire (Möller et al., 2008).
Deux autres pratiques culturales peuvent influencer l’azote résiduel, soit la fertilisation de l’engrais vert ou encore le semis de l’engrais vert dans les résidus de la culture précédente. La fertilisation apportera une quantité importante d’azote à l’engrais vert, ce qui favorisera la croissance des non-légumineuses mais qui n’affectera pas nécessairement les légumineuses. Cline et Silvernail (2001) ont testé la fertilisation d’engrais verts composés d’une vesce velue ou d’un seigle pur et d’un mélange de ces deux espèces, et arrivent à la conclusion que l’apport d’azote inorganique favorise la croissance du seigle pur ou en mélange mais ne va pas affecter les rendements de la vesce. Dans une recherche menée en Ontario, Vyn et al. (2000) ont comparé les rendements de différents engrais verts non fertilisés. Les rendements des engrais verts d’avoine, de seigle et de radis huileux implantés à la dérobée n’ont pas été très élevés étant donné l’absence de fertilisation (en moyenne 1.1, 1.2 et 1.0 t MS ha-1 respectivement), alors que les rendements du trèfle rouge, également
implanté en intercalaire d’une céréale, ont été plus considérables, avec 1.9 à 4.0 t MS ha-1
(Tableau 1). Il faut également s’attendre à une fixation biologique plus faible suite à la fertilisation de légumineuses, comme le démontrent plusieurs autres études (Hansen et Vinther, 2001; Hardarson et Atkins, 2003). À l’opposé, l’enfouissement des résidus de la culture précédente peut causer une immobilisation de l’azote résiduel du sol, ce qui nuira à
la croissance des non-légumineuses mais favorisera la fixation biologique des légumineuses (Patterson et LaRue, 1983).
Le calendrier de culture aura aussi un impact sur les rendements finaux et la composition des engrais verts. Il est préférable de semer les engrais verts le plus tôt possible dans la saison, de façon à profiter d’un meilleur établissement et d’un meilleur rendement (Richards et al., 1996). Les pratiques qui permettent aussi un établissement plus hâtif, comme l’implantation en intercalaire d’une autre culture qui sera récoltée ou l’utilisation d’espèces qui survivent à l’hiver, peuvent aussi favoriser un meilleur rendement et donc un apport en azote plus important (Oyer et Touchton, 1990; Snapp et al., 2005).
Le stade phénologique au moment de l’enfouissement, qui sera déterminé par la date d’enfouissement de l’engrais vert, a aussi son importance. De façon générale, un enfouissement plus tardif d’un engrais vert fera diminuer la concentration en N et ainsi, augmenter le rapport C/N de ses tissus, puisque les proportions de lignine et de glucides structuraux augmentent à mesure que la plante s’approche de la maturité (Ranells et Wagger, 1993). On remarque dans le Tableau 1 que les engrais verts ayant poussé durant de courtes périodes (2.5 mois) ont généralement une concentration en N plus élevée. En retardant de deux semaines en juin l’enfouissement d’engrais verts de moutarde, de sarrasin et de pois semés en avril, Miller et al. (2011) ont observé une augmentation du rapport C/N des résidus de respectivement 8, 4 et 5. Cela s’explique aussi par le fait qu’un engrais vert jeune possèdera une plus grande proportion de feuilles et cette proportion diminuera au profit des tiges à mesure que l’engrais vert arrive à maturité (Gallaher, 1991). Ceci peut fortement affecter la vitesse de décomposition de la biomasse puisque les feuilles peuvent se décomposer jusqu’à cinq fois plus rapidement que les tiges (Cobo et al., 2002).
Un enfouissement plus tardif favorisera aussi une biomasse plus élevée (Reeves, 1994), malgré que dans certains cas la biomasse maximale peut s’observer de trois à cinq semaines avant la maturité (Karpenstein-Machan et Stuelpnagel, 2000). De plus, cette biomasse plus importante ne signifie pas nécessairement un apport d’azote plus important. Au Kentucky, en retardant de huit jours au mois de mai la destruction d’un engrais vert d’hiver de vesce velue, Cline et Silvernail (2001) ont remarqué une augmentation de la biomasse (4.2 t MS
19 ha-1 vs 3.3 t MS ha-1) mais une baisse significative de la quantité d’azote foliaire (3.4 % vs
4.2 %), résultant en une accumulation totale d’azote semblable (141 kg N ha-1 vs 137 kg N
ha-1); la vesce velue était au stade végétatif à début de floraison lors de la destruction hâtive
et au stade fin de floraison lors de la destruction tardive, huit jours plus tard. Dans le cas du seigle, dans cette même étude, la destruction retardée n’a eu aucun effet significatif. En Caroline du Nord, Wagger (1989) a démontré que de retarder de deux semaines en avril la destruction d’un seigle, d’un trèfle incarnat ou d’une vesce velue augmentait respectivement la matière sèche de la biomasse de 39 %, 41 % et 61 % et son contenu en azote de 14 %, 23 % et 41 %. Dans ce cas-ci, la vesce velue était au stade de début de floraison lors de la destruction tardive. La destruction retardée a aussi causé une augmentation du rapport C/N et de la concentration en cellulose, hémicellulose et lignine pour les trois types d’engrais verts. Un allongement de la période de croissance de l’engrais vert peut donc mener ou non à une augmentation de la biomasse et de son contenu en azote.
2.4.2 Minéralisation de l’azote dans les sols
La récupération d’un maximum d’azote stocké dans l’engrais vert par la culture subséquente est d’une importance capitale, particulièrement en agriculture biologique puisque les sources d’azote sont moins nombreuses (Baggs et al., 2000). C’est donc pourquoi il est important de synchroniser les besoins de la culture avec la minéralisation de l’azote des résidus (Wagger, 1989). La minéralisation sera surtout influencée par trois catégories de facteurs : la qualité des résidus, les conditions environnementales et les pratiques culturales (Baggs et al., 2000).
2.4.2.1 Effets de l’engrais vert
Certaines caractéristiques propres à l’engrais vert même vont influencer la vitesse de décomposition et le transfert de l’azote à la culture suivante.
Fox et al. (1990) ont identifié le ratio (lignine + polyphénol) / N (LP/N) comme étant significativement et négativement corrélé avec la vitesse de minéralisation d’un résidu. En effet, après seulement deux semaines d’incorporation des résidus au sol en laboratoire,
l’engrais vert ayant le ratio LP/N le plus bas (Medicago sativa L.; LP/N = 1,48) avait déjà minéralisé 23,5% de son azote alors que l’engrais vert avec le ratio le plus élevé (Cassia
rotundifolia Pers.; LP/N = 7,52) avait causé une immobilisation équivalente à 12,8% de son
azote total. Il aura fallu neuf semaines pour que C. rotundifolia en arrive à un bilan N nul, alors que M. sativa avait déjà minéralisé 46,7% du N accumulé. Après douze semaines, l’azote minéralisé était respectivement de 47,2% et 11,2% pour les espèces M. sativa et C.
rotundifolia, la minéralisation de M. sativa ayant considérablement ralentie durant les trois
dernières semaines de l’expérience.
Le rapport C/N semble fortement influencer la minéralisation des engrais verts. En général, le rapport de 20:1 représente la limite sous laquelle on peut s’attendre à une minéralisation rapide des résidus et un apport important à la culture subséquente (Janzen et al., 1990) et au-dessus de laquelle il y a un risque d’immobilisation et de réduction de l’azote disponible à la culture subséquente (Aulakh et al., 1991). Dans une étude analysant les résultats combinés de huit expériences différentes, Vigil et Kissel (1991) ont obtenu un R2 de 0,75
lorsqu’ils essayaient de prédire la quantité d’azote minéralisé à l’aide de la variable de C/N, ce qui en faisait la meilleure variable testée. Dans son étude comparant l’effet d’un retardement de deux semaines dans la destruction d’un seigle, d’un trèfle incarnat et d’une vesce velue, Wagger (1989) a mesuré la vitesse de décomposition des résidus à l’aide d’un sac de mailles de nylon placé sur le sol au champ. Les résultats suivent la même tendance malgré que l’amplitude varie d’une année à l’autre. Par exemple, pour l’année 1984 et pour la date de destruction la plus hâtive, le pourcentage d’azote qui se trouve toujours dans le sac après huit semaines par rapport à la quantité initiale est de 54% pour le seigle (C/N = 36), de 25% pour le trèfle incarnat (C/N = 13) et de 5% pour la vesce velue (C/N = 8) alors que pour la destruction tardive après huit semaines, ces pourcentages sont de 60% pour le seigle (C/N = 44), de 44% pour le trèfle incarnat (C/N = 15) et de 16% pour la vesce velue (C/N = 9). Il faut aussi rappeler que le retardement de la destruction avait causé une augmentation de la concentration en cellulose, en hémicellulose et en lignine de tous les engrais verts et ce sont ces différences dans la composition des engrais verts que l’auteur identifie comme étant la cause de ces variations dans la vitesse de décomposition.
21
2.4.2.2 Effets des conditions environnementales
La température et l’humidité du sol sont des facteurs qui ont un impact considérable sur la vitesse de décomposition et de minéralisation de la matière organique. On reconnait que la libération de l’azote est lente, voire presque nulle, sous les 5 °C, qu’elle atteint son maximum aux environs de 35 °C pour recommencer à diminuer avec des températures plus élevées et que l’humidité idéale est celle de la capacité au champ avec un ralentissement à des teneurs en humidité inférieures ou supérieures à ce seuil (Cherr et al., 2006; Liebman et al., 2011). Plusieurs recherches étudiant la décomposition des résidus organiques viennent appuyer ces affirmations (Schomberg et al., 1994; Andersen et Jensen, 2001; Agehara et Warncke, 2005). Il peut cependant avoir certaines nuances à apporter à ces généralités. Andersen et Jensen (2001) observent que pour des résidus de raygrass évoluant dans les mêmes types de sol, une température de 3 °C mène à une minéralisation nette comparativement à une immobilisation nette à des températures de 9 et 15 °C peu de temps après l’enfouissement. Il semble que l’immobilisation soit davantage ralentie par la baisse de température que la minéralisation dans le contexte de cette expérience. Cependant, Cookson et al. (2002) arrivent à des résultats opposés et remarquent une hausse de l’immobilisation avec un passage des températures de 15 °C à 2 °C. Les auteurs proposent que cette observation puisse être due à une différence dans l’adaptabilité de la microflore présente dans les sols. De plus, plusieurs recherches semblent contredire l’idée selon laquelle la libération de l’azote est très lente en sol froid, leurs résultats démontrant une minéralisation et une nitrification considérable même pour des sols dont la température avoisine les 0 °C comme c’est le cas généralement en hiver (Breland, 1994; Chantigny et al., 2002; Cookson et al., 2002). C’est le cas de Magid et al. (2001), dont les résultats démontrent que même à 3oC, 25 à 40 % de l’azote accumulé par différents engrais verts de
légumineuses incorporés à un sol en laboratoire a été minéralisé après seulement 35 jours. Les résultats de leur analyse de variance suggèrent même que le type de résidus contribue jusqu’à dix fois plus à la régression que la température. Leurs résultats sur la nitrification étaient suffisamment élevés pour qu’ils estiment de 50 à 100 kg N ha-1 les quantités d’azote
pouvant être perdues par lessivage durant l’hiver, ce qui peut avoir de grandes implications sur les stratégies d’incorporation des résidus.
L’humidité est également un facteur important pour la minéralisation de l’azote, particulièrement en zone semi-aride. Dans leur étude menée au Montana, Miller et al. (2011) ont enfoui au mois de juin des engrais verts plantés en avril en prévision d’un semis de blé d’automne, et ce, durant deux années. Lors de la première année, les échantillons de sol prélevés dans les parcelles d’engrais verts de moutarde et de sarrasin au moment de semer le blé (soit plusieurs semaines après l’enfouissement de l’engrais vert) contenaient moins de nitrates que le témoin (jusqu’à -16 kg NO3-N ha-1), ce qui laisse penser à une
immobilisation. Au contraire, les parcelles de pois contenaient légèrement plus de nitrates que le témoin (3 à 9 kg NO3-N ha-1). Lors de la deuxième année, la majorité des parcelles
d’engrais verts démontraient une minéralisation nette, celle-ci allant de 8 kg NO3-N ha-1 à
29 kg NO3-N ha-1 pour le pois. Les auteurs attribuent cette différence à des précipitations
beaucoup plus faibles durant la première année, ce qui aurait ralenti la minéralisation.
2.4.2.3 Effets des pratiques culturales
Comme il a été vu précédemment, les mélanges d’engrais verts peuvent comporter plusieurs avantages. L’un de ces avantages est l’équilibrage du rapport C/N des résidus d’engrais verts. Puisque dans certains cas, l’enfouissement d’une légumineuse pure peut entrainer une minéralisation trop rapide alors que l’enfouissement d’une non-légumineuse pure peut mener à une immobilisation, un mélange peut permettre de trouver un équilibre profitable. Par exemple, Ranells et Wagger (1996) ont démontré qu’un mélange de seigle avec un trèfle incarnat (C/N = 24 à 28) ou de seigle et d’une vesce velue (C/N = 14 à 21) peut permettre de ralentir de près de trois semaines la minéralisation de l’azote par rapport à un engrais vert de trèfle incarnat pur (C/N = 17) ou de vesce velue pure (C/N = 11) sans pour autant causer une immobilisation aussi importante que le seigle pur (C/N = 38 à 42). Suite à des recherches sur la vitesse de minéralisation de différents mélanges de résidus en laboratoire, Kuo et Sainju (1998) ont établi que la proportion de seigle (C/N = 23) dans un mélange avec de la vesce velue (C/N = 10) ne devrait pas dépasser 60 % (C/N = 15) si on souhaite utiliser l’engrais vert pour une augmentation de la disponibilité en azote et profiter d’une minéralisation nette.
23 Le type de travail de sol au moment de détruire les engrais verts influencera aussi la libération de l’azote provenant de ceux-ci. De façon générale, un enfouissement des résidus d’engrais vert accélérera leur décomposition comparativement à les laisser en surface des sols dans un contexte de semis direct (Dou et al., 1994 ; Schomberg et al., 1994), en plus de faire augmenter la moyenne de minéralisation nette (Drinkwater et al., 2000). Par contre, tous les travaux de sol ne sont peut-être pas équivalents. Breland (1994) a observé une différence dans la vitesse de décomposition des résidus d’engrais verts de trèfle durant l’hiver norvégien selon la profondeur de ceux-ci. Après 167 jours, les résidus contenaient encore 73 %, 62 % et 56 % de leur azote initial lorsque laissés à la surface, enfouis à 5 centimètres ou enfouis à 30 centimètres, respectivement. Il est aussi possible que le type de travail de sol affecte le niveau de perturbation des sols et la vitesse de minéralisation de l’azote et ces éléments devraient être étudiés et adaptés à l’effet que l’on recherche lorsqu’on prévoit l’enfouissement de nos engrais verts (Tonitto et al., 2006).
Le moment de l’enfouissement des engrais verts est également un élément important à considérer, généralement en lien avec les conditions de températures et d’humidité du sol et le rythme de libération de l’azote recherché. Dans le climat froid et aride de la Saskatchewan, Foster (1990) a testé trois moments d’incorporation d’un engrais vert de mélilot en été (15 juin, 1er juillet et 15 juillet) pour la culture du blé de printemps l’année
suivante. Dans ce cas-ci, l’enfouissement le plus hâtif, soit celui du 15 juin, est celui qui a résulté en une plus grande quantité d’azote dans les sols au moment de semer le blé le printemps suivant, soit 82 kg NO3-N ha-1, comparativement à 75 et 62 kg NO3-N ha-1 pour
les incorporations du 1er et du 15 juillet, respectivement. Ceci s’explique sans doute par le
fait que les précipitations sont plus réduites sous ces climats semi-arides (moyenne annuelle = 355 mm) et donc, qu’il est plus difficile de perdre l’azote par lessivage durant l’hiver. Des résultats semblables ont été observés sous un climat semi-aride au Montana, où un enfouissement d’engrais verts de moutarde et de pois deux semaines plus tôt au mois de juin a augmenté parfois jusqu’à 12 kg NO3-N ha-1 la quantité d’azote disponible dans les
Au contraire, sous des climats plus humides comme ceux de la Scandinavie, la minéralisation de l’azote est ralentie et les pertes semblent être réduites au maximum lorsqu’on reporte l’enfouissement le plus tard possible à l’automne. Dans une étude menée en Finlande, Känkänen et al. (1998) ont testé l’effet d’une incorporation automnale hâtive (début septembre) et automnale tardive (fin octobre) de trois engrais verts (vesce velue, trèfle rouge et raygrass) ainsi que de résidus d’orge sur l’azote des sols. La vesce est l’engrais vert ayant accumulé le plus d’azote et celui où les nitrates se sont accumulés en plus grande quantité dans les sols suite aux enfouissements. Déjà à la fin de l’automne, les sols où la vesce avait été enfouie hâtivement contenaient de 48 à 56 kg NO3-N ha-1 dans la
couche 0-30 cm, comparativement à 21 à 30 kg NO3-N ha-1 dans les parcelles où la vesce
avait été enfouie tardivement. Dans la couche 30-60 cm, les accumulations étaient en moyenne respectivement de 22 et de 12 kg NO3-N ha-1, ce qui laisse penser que le lessivage
des nitrates vers les couches plus profondes du sol était déjà plus avancé dans les parcelles avec enfouissement hâtif et que celui-ci allait certainement se poursuivre au cours de l’hiver et du printemps suivant. Les mêmes tendances s’observaient pour les autres engrais verts, malgré que les quantités fussent beaucoup moins importantes.
Des exemples supplémentaires traitants de ces facteurs et tirés de la littérature scientifique sont présentés dans la prochaine partie, qui traite plus particulièrement de leur effet sur les rendements d’une culture subséquente.
2.5 Prélèvement de l’azote par la culture suivante
2.5.1 Effets de l’utilisation des engrais verts sur les rendements de la
culture suivante : quelques exemples issus de la littérature
Badaruddin et Meyer (1990), dans une étude menée au Dakota du Nord, ont comparé l’effet de cinq espèces de légumineuses utilisées en engrais verts annuels : une luzerne annuelle, une luzerne vivace, un mélilot, un trèfle rouge et une vesce velue; et de deux témoins : un blé fertilisé avec 112 kg N ha-1 et une jachère, sur les rendements d’une culture subséquente
de blé soumise à différentes régies de fertilisation (0, 75 et 150 kg N ha-1) l’année suivante.
Les sols étaient une argile limoneuse et un loam argileux limoneux. L’expérience fut répétée à deux reprises à deux sites, pour un total de quatre répétitions. Les engrais verts