Thérapie cellulaire pour le glaucome : régénération tissulaire grâce aux cellules souches mésenchymateuses
201
0
0
Texte intégral
Figure
+7
Outline
La MSC en thérapie cellulaire : utilisation dans la régénération tissulaire
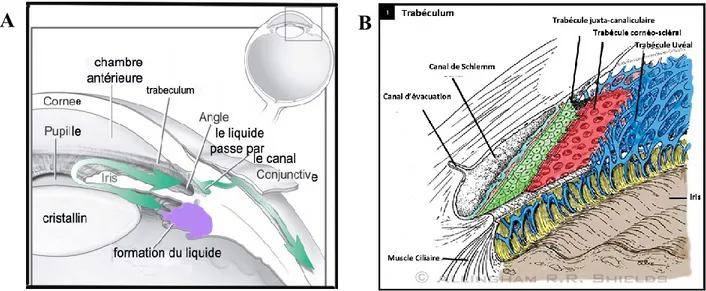
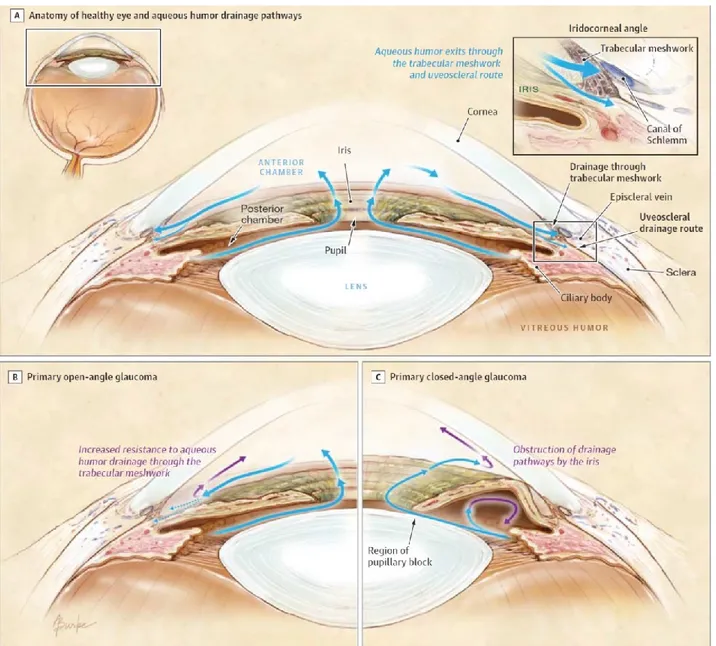
Le Glaucome
Les traitements contre le glaucome
Les cellules souches oculaires et la régénération tissulaire
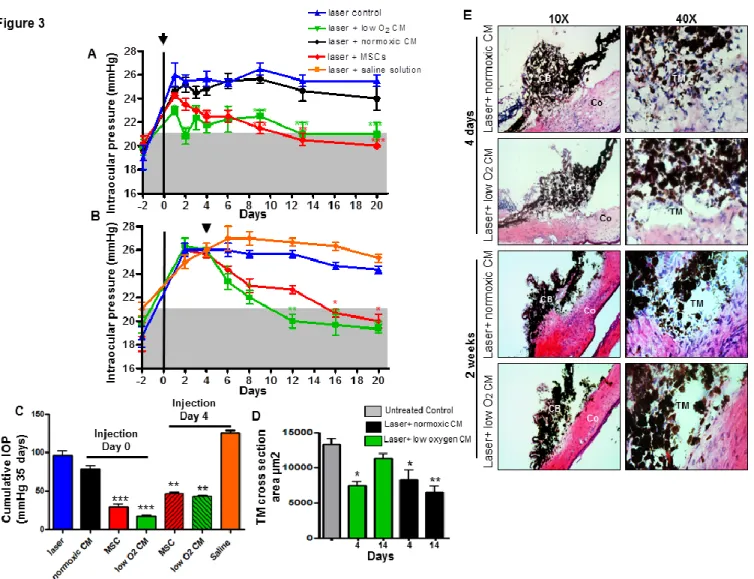
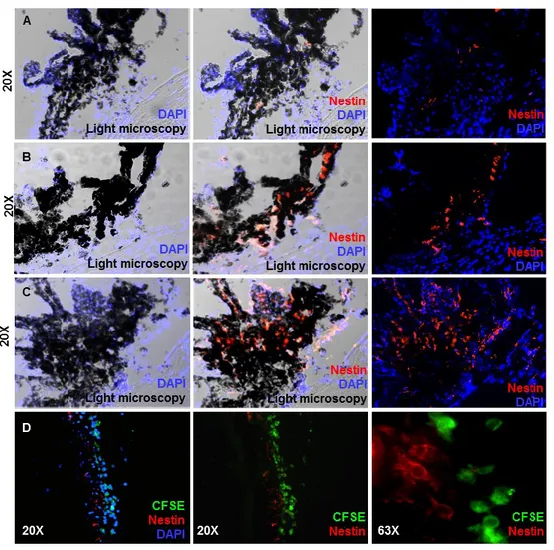
Mesenchymal stem cells promote ocular regeneration through macrophage recruitment
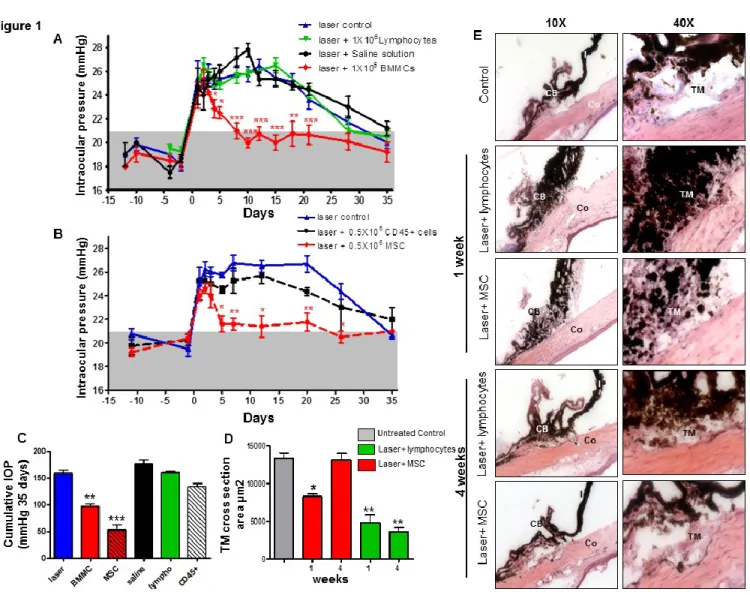
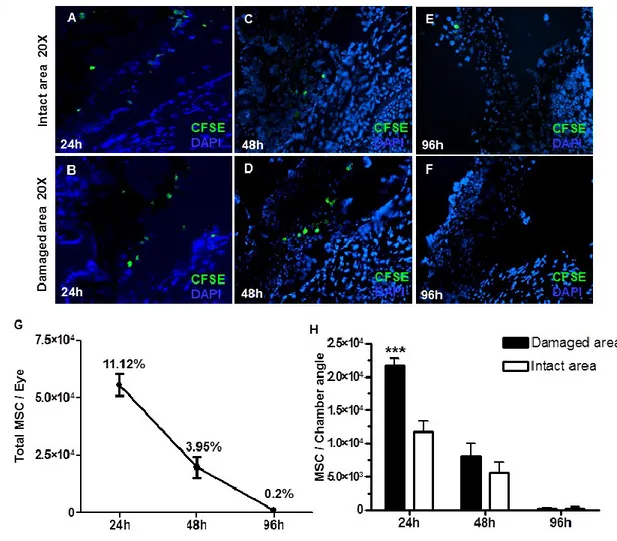
Les MSC en thérapie cellulaire pour le glaucome : mécanismes d’action
Transfert vers la clinique
Modèle proposé des mécanismes de régénération par les MSC
Documents relatifs