Université de Sherbrooke
Rôle du cortex pariétal dans le traitement des erreurs de prédiction sensorielles et impact d’incitatifs sur l’activité cérébrale pendant la préparation d’un mouvement
Par
Félix-Antoine Savoie
Sciences des radiations et imagerie biomédicale
Thèse présentée à la Faculté de médecine et des sciences de la santé en vue de l’obtention du grade de philosophae doctor (Ph.D.)
en Science des radiations et imagerie biomédicale
Sherbrooke, Québec, Canada 28 avril 2020
Membres du jury d’évaluation Pierre-Michel Bernier, Ph.D.,
Université de Sherbrooke, Faculté des sciences de l’activité physique Département de kinanthropologie.
Kevin Whittingstall, Ph.D.,
Université de Sherbrooke, Faculté de médecine et sciences de la santé Département de médecine nucléaire et radiobiologie
John Francis Kalaska, Ph.D.,
Université de Montréal, Faculté de Médecine Département de neurosciences
Jean-François Lepage, Ph.D.,
Université de Sherbrooke, Faculté de médecine et sciences de la santé Département de pédiatrie
Darel Hunting, Ph.D.,
Université de Sherbrooke, Faculté de médecine et sciences de la santé Département de médecine nucléaire et radiobiologie
If the brain were simple enough for us to understand it, we would be too simple to understand it. Ken Hill
My Ph.D. was a prediction error. Félix-Antoine Savoie
v
R
ESUMERôle du cortex pariétal dans le traitement des erreurs de prédiction sensorielles et impact d’incitatifs sur l’activité cérébrale pendant la préparation d’un mouvement
Par
Félix-Antoine Savoie
Programme de Sciences des radiations et imagerie biomédicale
Thèse présentée à la Faculté de médecine et des sciences de la santé en vue de l’obtention du diplôme de philosophiae doctor (Ph.D.) en sciences des radiations et imagerie biomédicale, Faculté de médecine et des sciences de la santé, Université de Sherbrooke,
Sherbrooke, Québec, Canada, J1H 5N4
Selon la théorie du contrôle rétroactif optimal, l’objectif du cerveau est de produire des mouvements qui maximisent la probabilité d’atteindre un objectif tout en minimisant les coûts énergétiques associés. Pour y arriver, le cerveau doit savoir s’adapter à de nouvelles relations sensorimotrices, car celles-ci occasionnent des erreurs qui mettent en péril l’atteinte de l’objectif moteur et nécessitent des corrections coûteuses. À cet effet, le premier objectif de cette thèse était d'investiguer le rôle du cortex pariétal dans le traitement des erreurs qui alimentent l'adaptation implicite à une nouvelle relation sensorimotrice. Pour y arriver, deux projets ont été réalisés. Dans le premier projet, l'électroencéphalographie (EEG) a été enregistrée chez 15 participants lors d'une tâche de pointage faisant usage d’une perturbation visuomotrice pour identifier la signature neuronale des erreurs qui induisent l'adaptation implicite. Les résultats ont révélé de fortes modulations d’activité EEG au-dessus des régions pariétales en réponse à ces erreurs, ce qui suggère que le cortex pariétal est impliqué dans leur traitement. Dans le deuxième projet, 28 participants ont reçu une stimulation magnétique transcrânienne (TMS) à impulsion unique au-dessus du cortex pariétal pour tenter d'établir un lien causal entre l’activité EEG observée dans le premier projet et les ajustements moteurs implicites occasionnés par une perturbation visuomotrice. Contrairement à notre hypothèse, la TMS à impulsion unique n'a pas eu d’impact détectable sur les ajustements moteurs encourus par une perturbation visuomotrice. Ceci suggère que l'activité neuronale observée dans le premier projet n’était peut-être pas causalement liée au processus d'adaptation implicite. Ensemble, ces résultats suggèrent que le cortex pariétal est sensible aux erreurs à l’origine de l’adaptation implicite, malgré qu’il ne soit peut-être pas directement impliqué dans l’émergence des ajustements moteurs implicites occasionnés par une rotation visuomotrice. Les réponses pariétales observées dans le premier projet pourraient donc être liées à une autre facette de l’adaptation sensorimotrice, telle que l’adaptation explicite, le contrôle en ligne ou encore la recalibration proprioceptive. Dans tous les cas, ces résultats sont un premier pas vers une meilleure compréhension du rôle des différentes régions cérébrales dans le processus d’adaptation sensorimotrice.
En théorie, le cerveau devrait être plus enclin à mobiliser des ressources sensorimotrices lorsqu’un objectif a une plus grande valeur. En l’occurrence, le deuxième objectif de cette thèse était d'investiguer comment la valeur ajoutée d’un incitatif monétaire
vi influence l’activité cérébrale pendant la préparation d’un mouvement. Pour ce faire, l'activité EEG fut enregistrée chez 23 participants alors qu’ils accomplissaient une tâche dans laquelle l'obtention de récompenses ou l'évitement de punitions dépendait de la précision d’un mouvement de pointage manuel vers une cible. Les résultats ont révélé de fortes modulations de puissance spectrale dans l’étendu de fréquence bêta (13 - 30 Hz) sur un vaste territoire pariéto-frontal dans des conditions avec, vs. sans incitatifs et ces modulations étaient corrélées avec la durée des mouvements (c.-à-d., la vigueur). Il est à noter que, bien que les incitatifs positifs et négatifs aient ultimement donné lieu à des profils de puissance spectrale bêta similaires, les incitatifs positifs ont modulé la puissance spectrale bêta nettement plus tôt que les incitatifs négatifs. Ainsi, ces résultats démontrent un lien entre l'activité oscillatoire bêta, la motivation et la vigueur. Par ailleurs, ils supportent des résultats récents montrant que le traitement de stimuli associés à des récompenses est plus rapide que celui de stimuli associés à des punitions.
Mots clés : Adaptation sensorimotrice, rotation visuomotrice, contrôle visuomoteur, erreur
vii
S
UMMARYRole of the parietal cortex in sensory prediction error processing and the impact of incentives on neural activity during movement preparation
By
Félix-Antoine Savoie
Radiation sciences and biomedical imaging program
Thesis presented to the Faculté de médecine et des sciences de la santé in order to obtain a philosophiae doctor (Ph.D.) diploma in radiation sciences and biomedical imaging from the
Faculté de médecine et des sciences de la santé, Université de Sherbrooke, Sherbrooke, Québec, Canada, J1H 5N4
In the past two decades, many neuroscientists have framed motor control as an optimal feedback control problem. In this framework, it is thought that the brain aims to produce movements that maximize the probability of reaching the task goal and minimize motor costs. To do so, the brain must be able to adapt to novel sensorimotor relationships, as these cause errors that both jeopardize task success and necessitate costly corrective movements. In this light, the first goal of this thesis was to investigate whether the parietal cortex is involved in processing the errors thought to drive implicit adaptation to a novel sensorimotor relationship. To address this question, two projects were carried out. In the first project, electroencephalography (EEG) was recorded in 15 participants during a visuomotor perturbation task to identify the neural signature of implicit adaptation-driving errors. The results revealed strong parietal EEG responses to such errors, suggesting that the parietal cortex is involved their processing. In the second project, 28 participants received single-pulse transcranial magnetic stimulation (TMS) over the parietal cortex to try to establish a causal relationship between the EEG responses observed in the first project and the implicit motor output changes incurred by a visuomotor perturbation. In contrast to our hypothesis, single-pulse TMS had no detectable impact on the motor output changes incurred by the visuomotor perturbation, suggesting that the neural activity observed in the first project may not have been causally related to implicit adaptation per se. Overall, these results demonstrate that the parietal cortex is sensitive to the errors that drive implicit adaptation, although it may not contribute to implicit changes in motor output. Parietal responses to visuomotor perturbations may thus subtend some other aspect of performance improvement during adaptation, such as explicit adaptation, online control or proprioceptive recalibration. In any case, these results are a first step towards understanding how different brain areas contribute to performance improvement during adaptation.
In theory, the brain should be willing to mobilize more sensorimotor resources to reach more valuable motor goals. Given this, the second goal of this thesis was to investigate how the added value of a monetary incentive influences neural activity during movement preparation. To do so, EEG was recorded as 23 participants undertook a monetary incentive delay task in which reward obtainment and/or punishment avoidance depended on the accuracy of a goal-directed reaching movement. During the delay period, strong beta frequency (13 – 30 Hz) spectral power modulations were observed over a broad parieto-frontal area in conditions with, vs. without incentives and these modulations correlated with
viii movement times (i.e., vigor). Of note, although both positive and negative incentives gave rise to similar spectral beta power profiles late in the delay period, positive incentives modulated spectral beta power noticeably earlier compared to negative incentives. Thus, these findings provide a link between beta oscillatory activity, incentive motivation and vigor. Moreover, they support recent work showing that the processing of reward-predicting stimuli is quicker than that of punishment-predicting stimuli.
Keywords: Sensorimotor adaptation, visuomotor rotation, visuomotor control, sensory
ix
T
ABLE DES MATIERESRésumé... v
Summary ... vii
Table des matières ...ix
Liste des figures ... xiii
Liste des abréviations ... 1
Introduction ... 2
Chapitre 1 : Le contrôle moteur ... 4
1.1 Transformations sensorimotrices et le modèle inverse ... 4
1.2 Théorie du contrôle rétroactif optimal (OFC) ... 4
1.2.1 Les champs moteurs frontaux comme contrôleurs ... 6
1.3 Estimation d’état ... 7
1.3.1 Le cervelet comme siège des modèles directs ... 9
1.3.2 Le cortex pariétal comme estimateur d’état ... 10
Chapitre 2 : L’adaptation sensorimotrice ... 11
2.1 Méthodes pour investiguer l’adaptation sensorimotrice ... 12
2.2 Les processus explicite et implicite en adaptation sensorimotrice ... 13
2.3 L’adaptation sensorimotrice est stimulée par les erreurs ... 15
2.3.1 Erreurs explicites ... 15
2.3.2 Erreurs de prédiction sensorielles (EPS) ... 17
2.4 Substrats potentiels pour l’adaptation explicite/implicite ... 19
2.4.1 Le cervelet ... 19
2.4.2 Le cortex pariétal ... 22
Chapitre 3 : Projet Laphroaig ... 28
3.1 Problématique de recherche, objectif et hypothèse ... 28
3.1.1 EEG ... 29
3.1.2 Marqueurs potentiels d’EPS en EEG... 31
x 4.1 Résumé... 34 4.2 Abstract... 36 4.3 Introduction ... 37 4.4 Methods ... 39 4.4.1 Participants... 39 4.4.2 Experimental set-up ... 39 4.4.3 Experimental task ... 40
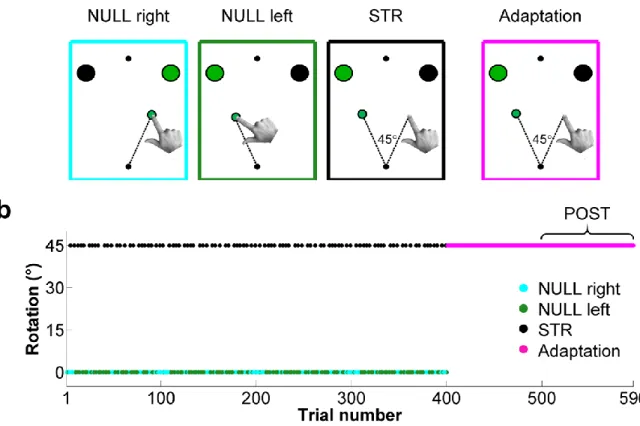
4.4.4 Experimental protocol and conditions ... 41
4.4.5 Kinematic data recording and analysis ... 43
4.4.6 Kinematic-based trial rejection ... 43
4.4.7 EEG data acquisition, processing and time-frequency decomposition ... 44
4.4.8 EEG data analysis ... 46
4.4.9 Statistical analyses ... 47
4.4.10 Data availability statement ... 49
4.5 Results ... 49
4.5.1 Kinematic results ... 49
4.5.2 Right Parietal ROI ... 51
4.5.3 Mid-Frontal ROI ... 58
4.6 Discussion ... 61
4.7 Acknowledgements ... 67
Chapitre 5 : Projet Aberlour ... 68
5.1 Problématique de recherche, objectif et hypothèse ... 68
Chapitre 6: Article 2 (Aberlour)... 70
6.1 Résumé... 71 6.2 Abstract... 72 6.3 Introduction ... 74 6.4 Methods ... 76 6.4.1 Participants... 76 6.4.2 Experimental set-up ... 76 6.4.3 Experimental task ... 77
6.4.4 Movement-related data recording and analysis ... 80
6.4.5 Movement-related trial rejection ... 81
6.4.6 TMS protocol ... 81
xi
6.5 Results ... 85
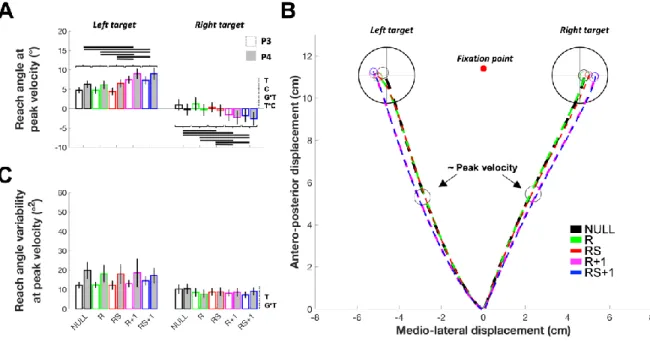
6.5.1 Reach angle at peak velocity ... 85
6.5.2 Reach angle variability at peak velocity ... 87
6.5.3 Finalx ... 87 6.5.4 Finalx variability ... 89 6.5.5 Finaly ... 90 6.5.6 Finaly variability ... 90 6.5.7 RT ... 91 6.5.8 MT ... 92 6.6 Discussion ... 93
Chapitre 7 : Incitatifs, activité cérébrale et performance motrice ... 99
7.1 La valeur influence le contrôle moteur ... 99
7.2 Les incitatifs modulent l’activité cérébrale ... 100
Chapitre 8 : Projet Diplomatico ... 103
8.1 Problématique de recherche, objectif et hypothèse ... 103
Chapitre 9: Article 3 (Diplomatico) ... 106
9.1 Résumé... 107 9.2 Abstract... 108 9.3 Introduction ... 110 9.4 Methods ... 111 9.4.1 Participants... 111 9.4.2 Experimental set-up ... 112 9.4.3 Experimental task ... 113
9.4.4 Movement-related data recording and analysis ... 117
9.4.5 Movement-related trial rejection ... 118
9.4.6 EEG data acquisition, processing and time-frequency decomposition ... 118
9.4.7 Experimental design and statistical analysis ... 120
9.5 Results ... 121
9.5.1 Motor behavior ... 121
9.5.2 Beta power responses... 124
9.5.3 Relationship between beta power and models ... 128
9.5.4 Relationship between beta power and movement-related variables ... 130
9.6 Discussion ... 132
xii
Chapitre 10: Discussion ... 138
10.1 Projet 1 - Laphroaig ... 138
10.1.1 Résumé ... 138
10.1.2 Les différences d’ERP entre STR et POST ... 138
10.1.3 Différences de puissance spectrale entre STR et POST ... 139
10.1.4 Un circuit pariéto-cérébelleux à l’origine de l’EPS? ... 141
10.1.5 Différences de FRN entre STR et POST... 142
10.2 Projet 2 - Aberlour ... 143
10.2.1 Résumé ... 143
10.2.2 Limites du protocole TMS utilisé dans Aberlour... 144
10.2.3 Interprétations alternatives de Laphroaig à la lumière des résultats d’Aberlour ... 144
10.2.3.1 Contrôle en ligne ... 144
10.2.3.2 Recalibration proprioceptive ... 145
10.3 Projet 3 - Diplomatico ... 147
10.3.1 Résumé ... 147
10.3.2 Puissance spectrale beta et motivation ... 148
10.3.3 Oscillations beta et interactions entre les noyaux gris centraux et le néocortex 149 10.3.4 Oscillations beta et inférence active ... 150
10.4 Conclusion et perspectives ... 151
Remerciements ... 154
xiii
L
ISTE DES FIGURESFigure 4.1 - Experimental conditions and perturbation schedule. ... 40
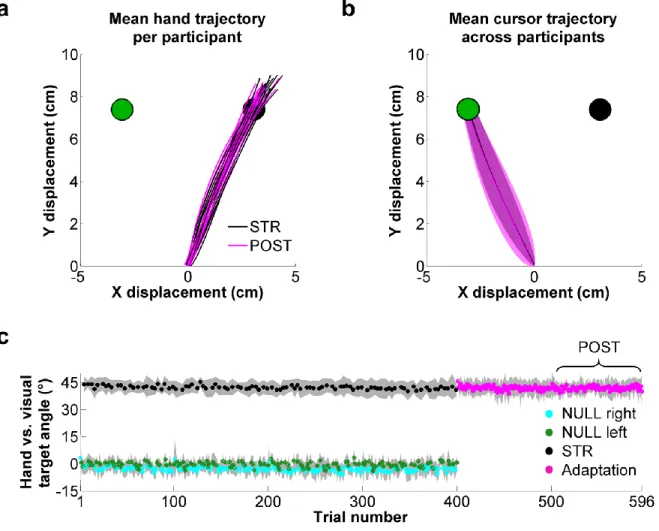
Figure 4.2 - Reach trajectories and hand vs. visual target angle at peak velocity. ... 50
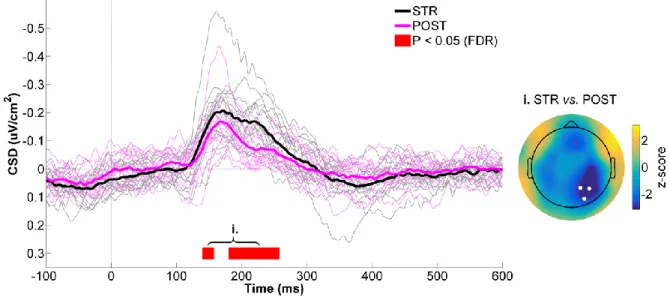
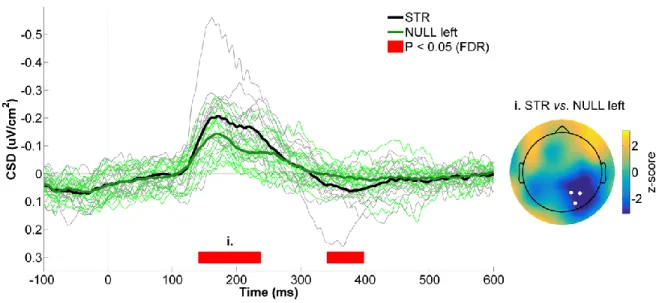
Figure 4.3 - CSD activity recorded at the right parietal ROI for STR vs. POST. ... 52
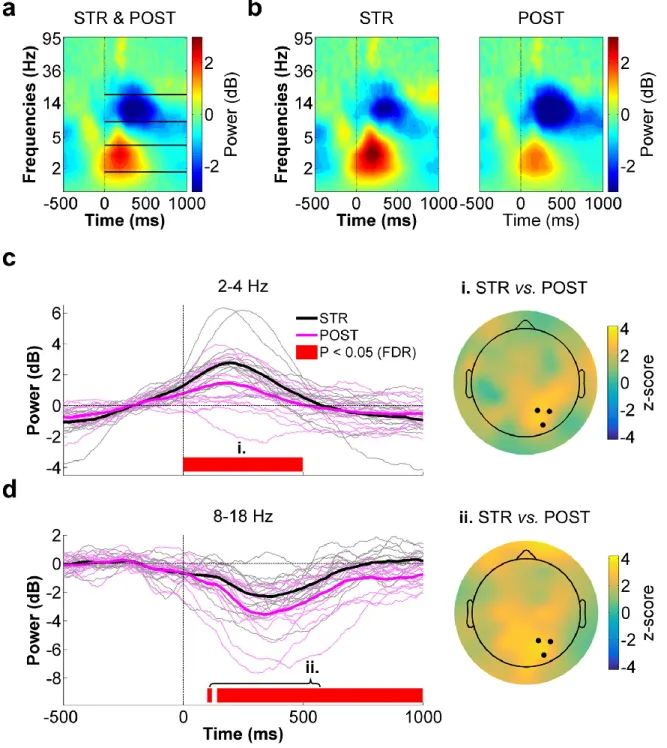
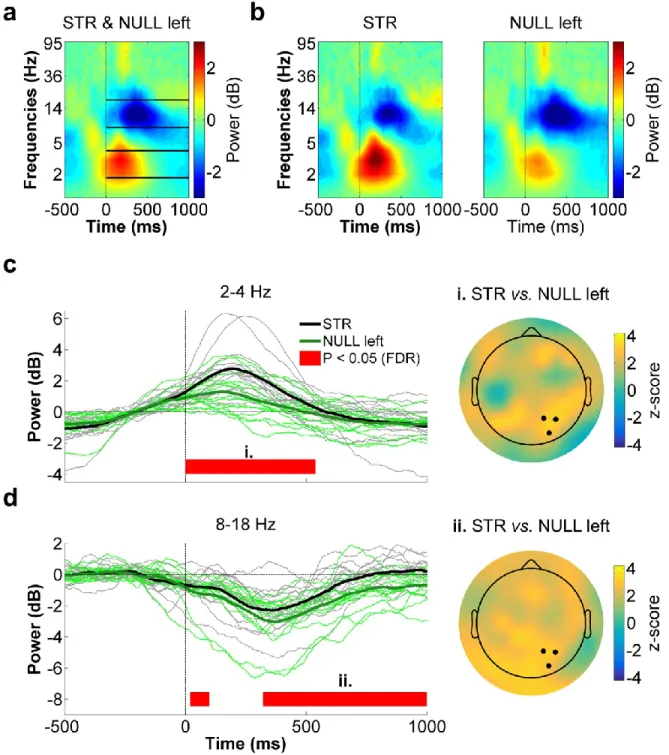
Figure 4.4 - Right parietal ROI time-frequency data for STR vs. POST. ... 53
Figure 4.5 - CSD activity recorded at the right parietal ROI for STR vs. NULL left. ... 56
Figure 4.6 - Right parietal ROI time-frequency data for STR vs. NULL left. ... 57
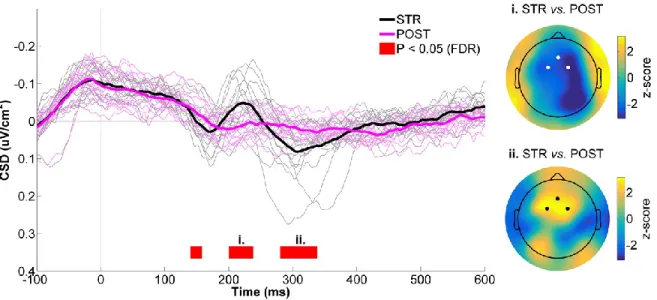
Figure 4.7 - CSD activity recorded at the mid-frontal ROI for STR vs. POST. ... 59
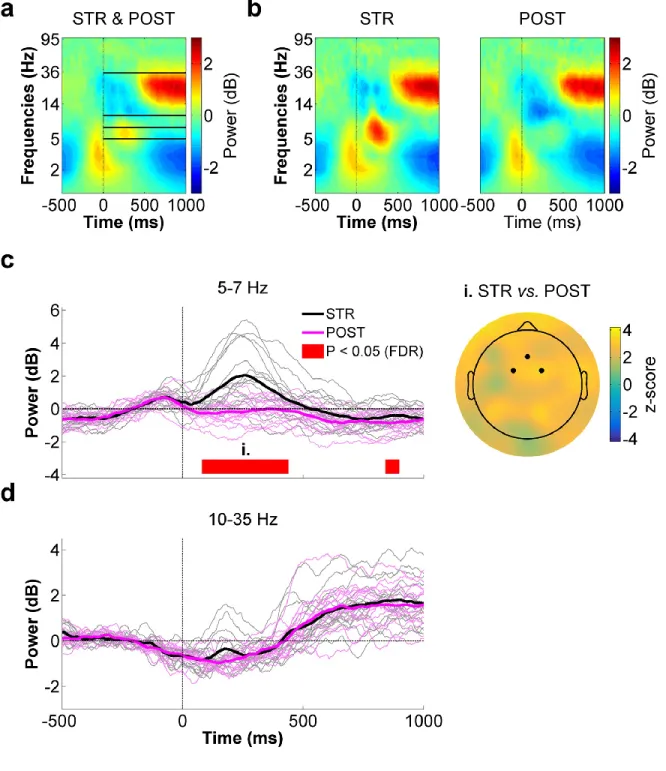
Figure 4.8 - Mid-frontal ROI time-frequency data for STR vs. POST. ... 60
Figure 6.1 - Study design. ... 78
Figure 6.2 - Hand direction data. ... 86
Figure 6.3 - Final hand position data. ... 88
Figure 6.4 - Movement onset and duration data. ... 92
Figure 9.1 - Schematic representation of the experimental apparatus, trial timeline and conditions. ... 113
Figure 9.2 - Movement-related variables. ... 123
Figure 9.3 - Paired comparisons of beta power across INCENTIVE levels during the delay period. ... 126
Figure 9.4 - Paired comparisons of beta power across PROBABILITY and INCENTIVE × PROBABILITY levels during the delay period. ... 127
Figure 9.5 - Spearman correlations between the models (i.e., expected value and motivational salience) and delay period beta power across INCENTIVE levels. ... 130
Figure 9.6 - Spearman correlations between the movement-related variables and delay period beta power across INCENTIVE levels. ... 132
1
L
ISTE DES ABRÉVIATIONSANOVA Analyse de variance (analysis of variance)
BP Puissance spectrale pendant la période de base (baseline spectral power) CAD Dollars canadiens (canadian dollars)
CSD Densité de courant source (current source density) EEG Électroencéphalographie (electroencephalography)
EPR Erreur de prédiction de récompense (reward prediction error) EPS/SPE/PE Erreur de prédiction sensorielle (sensory prediction error) ERP Potentiel évoqué par un évènement (event-related potential) FDR Taux de faux positifs (false discovery rate)
Finalx Position finale de la main dans l’axe médiolatéral (final medio-lateral hand position)
Finaly Position finale de la main dans l’axe antéropostérieur (final antero-posterior hand position)
FRN Négativité de rétroaction (feedback-related negativity)
ICA Analyse de composantes indépendantes (independent component analysis) IRMf/fMRI Imagerie par résonance magnétique fonctionnelle (functional magnetic
resonance imaging)
M1 Cortex moteur primaire (primary motor cortex)
MEG Magnétoencéphalographie (magnetoencephalography) MT Temps de mouvement (movement time)
NULL Essai sans rotation (null rotation trial)
OFC Contrôle rétroactif optimal (optimal feedback control) POST Essai post-adaptation (post-adaptation trial)
R Essai avec rotation (rotation trial)
R+1 Essai post-rotation (post-rotation trial) ROI Région d’intérêt (region of interest)
RP Puissance spectrale médiane (median spectral power)
RS Essai avec rotation + stimulation (rotation + stimulation trial)
RS+1 Essai post-rotation + stimulation (post-rotation + stimulation trial) RT Temps de réaction (reaction time)
SARA Échelle d’évaluation et de l’Ataxie (Scale for the Assessment and Rating of Ataxia) SE Erreur standard (standard error)
SD Écart type (standard deviation) STR Essai stratégie (strategy trial)
tDCS Stimulation transcrânienne par courant direct (transcranial direct current stimulation) TMS Stimulation magnétique transcrânienne (transcranial magnetic stimulation)
2
I
NTRODUCTIONLa motricité est la seule voie par laquelle nous pouvons interagir avec l’environnement. Sans elle, il est impossible de se nourrir, de communiquer ou d’éviter une situation menaçante. Bien que la motricité soit d’une importance fondamentale dans nos vies, nous la prenons bien souvent pour acquise. Après tout, il est si facile de tendre le bras pour prendre une tasse de café lorsqu’on est enfermé dans une petite pièce sombre en train d’écrire une thèse. Sachez, toutefois, qu’après 5 ans d’études dans le domaine de l’apprentissage et du contrôle moteur, un geste aussi simple soulève bien des questions.
Dans les pages qui suivent, il deviendra évident que le cerveau doit prédire les conséquences sensorielles des mouvements qu’il produit s’il veut que ceux-ci soient exécutés de manière optimale (Wolpert and Miall, 1996). Pour qu’elles soient utiles au contrôle moteur, toutefois, il est impératif que ces prédictions calquent précisément les réafférences du mouvement exécuté. Conséquemment, on considère que le cerveau compare continuellement ses prédictions aux réafférences sensorielles, et utilise les différences (c.-à-d., les erreurs) pour réviser ses prédictions ultérieures (Shadmehr et al., 2010a). Dans la présente thèse, l’objectif des deux premiers projets était d’investiguer le rôle du cortex pariétal dans le processus de révision des prédictions sensorielles.
Pour que les prédictions sensorielles mentionnées ci-haut optimisent le contrôle moteur, il faut d’abord bouger. Mais pourquoi bouge-t-on? Selon plusieurs, c’est la valeur associée à l’atteinte d’un objectif moteur qui pousse à l’action et qui détermine comment celle-ci sera exécutée (Shadmehr et al., 2019). Conséquemment, il n’est pas surprenant que plusieurs auteurs aient rapporté que la valeur d’une action influence l’activité cérébrale qui précède son exécution (p.ex.,(Iyer et al., 2010). Depuis environ dix ans, une question qui a fait couler beaucoup d’encre concerne ce que représentent ces modulations d’activité cérébrale. L’objectif du troisième projet de cette thèse était d’adresser cette question.
Le présent document se divise en 7 chapitres. Question d’établir des concepts importants pour la suite des choses, le chapitre 1 fait un bref résumé de la théorie du contrôle
3 rétroactif optimal (optimal feedback control), qui servira de cadre conceptuel pour les travaux présentés. Le chapitre 2, quant à lui, met la table pour les deux premiers projets de cette thèse (chapitres 3 et 4, respectivement). Plus précisément, ce chapitre concerne les notions importantes en adaptation sensorimotrice en mettant un accent particulier sur le traitement des erreurs et le rôle de celles-ci dans la révision des prédictions sensorielles. Dans le chapitre 5, l’influence d’incitatifs sur l’activité cérébrale et la performance motrice sera développée afin de mettre la table pour le troisième projet de cette thèse (chapitre 6). Enfin, pour mieux situer l’impact des travaux menés pendant cette thèse, les résultats des trois projets seront discutés dans le chapitre 7.
4
C
HAPITRE1 :
L
E CONTRÔLE MOTEUR1.1 Transformations sensorimotrices et le modèle inverse
Produire des mouvements précis et coordonnés en vue d’atteindre un objectif n’est pas une mince affaire. Selon les théories actuelles, le cerveau doit accomplir ce qu’on appelle des transformations sensorimotrices, c’est-à-dire convertir l’information véhiculée par les sens en commandes motrices qui activeront la musculature périphérique pour générer des mouvements (Kalaska and Crammond, 1992; Kandel et al., 2012). Pour y arriver, le cerveau utiliserait un modèle inverse, soit un répertoire de correspondances entre des états désirés et des commandes motrices (Wolpert and Miall, 1996; McNamee and Wolpert, 2019). Le modèle inverse déterminerait comment des commandes motrices (ut) appliquées à un
effecteur dans un état xt change l’état prochain de cet effecteur (xt+1). Dans ce modèle, l’état
(xt) de l’effecteur peut référer à toute variable cinématique pouvant lui être attribuée (angles
articulaires, vitesses angulaires articulaires, etc.), alors que les commandes motrices (ut)
réfèrent aux moments articulaires (McNamee and Wolpert, 2019).
1.2 Théorie du contrôle rétroactif optimal (OFC)
Imaginez un instant que vous désirez prendre une tasse de café à portée de main. Dans cet exemple, une infinité de mouvements différents pourraient vous permettre d’atteindre la tasse (autrement connu comme le problème des degrés de liberté,(McNamee and Wolpert, 2019). Malgré cet éventail de possibilités, le système moteur génère généralement des mouvements relativement droits avec un profil de vitesse en forme de cloche. Comment le cerveau fait-il pour arriver à une solution motrice aussi stéréotypée? Selon la théorie du contrôle rétroactif optimal (optimal feedback control, OFC), le cerveau établirait une loi de contrôle optimale, dans le sens où elle minimise les coûts métaboliques tout en maximisant les chances d’atteindre l’objectif moteur. Cette loi () spécifie essentiellement les commandes motrices (u) qui devraient être produites à l’instant (t) en fonction de l’état de l’effecteur (x) et d’une cible (y) [ut = (xt,yt),(Todorov and Jordan, 2002; Scott, 2004;
5 Franklin and Wolpert, 2011; Scott, 2012; Shadmehr and Mussa-Ivaldi, 2012; Haith and Krakauer, 2013; Scott et al., 2015; Scott, 2016; McNamee and Wolpert, 2019)]. Conséquemment, selon l’OFC, le cerveau ne préprogrammerait pas une trajectoire de mouvement désirée avant l’initiation du mouvement. Plutôt, les transformations sensorimotrices qu’il accomplit à chaque instant du mouvement naissent de son estimation continuelle de l’état de l’effecteur et de la cible. L’OFC s’apparente donc à un servomécanisme qui estime l’état des variables pertinentes pour implémenter des actions optimales.
En OFC, la loi de contrôle dicte les transformations sensorimotrices à accomplir en mettant en place des « gains sensoriels » (feedback gains,(Scott, 2004, 2012). Pour être optimaux, ces gains pondèrent à la baisse les afférences peu importantes pour l’accomplissement de l’objectif moteur et vice versa pour celles qui sont déterminantes pour la performance (c.-à-d., les erreurs qui mettent en péril l’atteinte de l’objectif). En pondérant ainsi les afférences sensorielles, les transformations sensorimotrices effectuées par le contrôleur sont dictées par leur impact sur l’optimalité du mouvement (principe de l’intervention minimale,(Todorov and Jordan, 2002). En effet, lorsqu’une perturbation inattendue altère l’état de l’effecteur ou de la cible lors d’un mouvement, la magnitude des réponses correctrices engendrées par le système dépend de l’impact de cette perturbation sur l’atteinte de l’objectif moteur (Diedrichsen, 2007; Liu and Todorov, 2007; Franklin and Wolpert, 2008; Knill et al., 2011; Nashed et al., 2012). Par exemple, Nashed et al. (2012) ont investigué l’impact d’une perturbation mécanique latérale sur le contrôle en ligne pendant des mouvements de pointage vers une petite cible ronde (petite marge d’erreur) et d’une longue cible rectangulaire (grosse marge d’erreur). En accord avec l’OFC, ces auteurs ont montré que les participants apportaient des corrections plus importantes pour des mouvements vers la petite cible ronde que la longue cible rectangulaire. En évitant de corriger les erreurs sans importance, le système évite l’engagement inutile de fibres musculaires supplémentaires qui occasionnerait des coûts métaboliques supplémentaires (McNamee and Wolpert, 2019). Par ailleurs, cette stratégie d’optimisation diminue la variabilité du mouvement, causée par le bruit neuromusculaire qui croît avec l’amplitude des efférences motrices (Jones et al., 2002; Faisal et al., 2008).
6
1.2.1 Les champs moteurs frontaux comme contrôleurs
Quelles régions cérébrales pourraient jouer le rôle du contrôleur dans le schème de l’OFC? Vu ce qui précède, un contrôleur optimal devrait 1) avoir accès à une estimation de l’état de l’effecteur et de la cible pendant l’exécution du mouvement; et 2) être en mesure de véhiculer de l’information aux motoneurones de la moelle épinière (c.-à-d., avoir des connexions cortico-spinales). Parmi les régions cérébrales impliquées dans la sensorimotricité, les régions caudales du cortex frontal, notamment les cortex prémoteur et moteur primaire (M1), semblent des candidats intéressants pour mettre en place une loi de contrôle (Shadmehr and Krakauer, 2008). Premièrement, ces deux régions sont dotées de connexions réciproques avec les régions pariétales (Johnson et al., 1996; Wise et al., 1997; Rizzolatti and Luppino, 2001) probablement impliquées dans l’estimation d’état (voir section 1.3.2). Deuxièmement, il est connu que le cortex prémoteur/M1 compte des projections cortico-spinales (Dum and Strick, 2002), leur conférant tous deux la capacité d’influencer l’activité des interneurones et motoneurones spinaux qui dicte l’activité des muscles (Kalaska, 2009).
Avec le développement de méthodes d’enregistrement intracortical, plusieurs chercheurs ont tenté de déterminer ce qu’encodent les neurones des champs moteurs frontaux avant et pendant l’exécution de mouvements (Kalaska, 2009). Dans certaines études, l’objectif était de déterminer si l’activité des neurones représentait le mouvement dans un cadre de référence extrinsèque (c.-à-d., en termes de déplacement de l’effecteur terminal) ou intrinsèque (c.-à-d., en termes de motions articulaires) (p.ex,(Scott and Kalaska, 1997). D’autres encore, ont cherché à déterminer si l’activité des neurones du cortex prémoteur et/ou du M1 représente la cinématique ou la cinétique d’un mouvement (p.ex. (Kalaska et al., 1989). Toutefois, et malgré plusieurs études sur le sujet, il n’y a toujours pas de consensus quant aux paramètres d’un mouvement que représente l’activité de neurones individuels en contexte de contrôle moteur (Shenoy et al., 2013; Kalaska, 2019).
7 Selon la maxime bien connue, « le tout est plus grand que la somme de ses parties ». Cette idée résonne bien avec un cadre théorique qui a refait surface depuis quelques années en contrôle moteur et qui conceptualise l’activité neurale comme un système dynamique (Scott, 2008; Shenoy et al., 2013). Selon cette perspective, les neurones des cortex prémoteurs et M1 (c.-à-d., les parties) forment un système interconnecté dont les changements d’état (c.-à-d., le tout) déterminent les commandes motrices qui émergent (Churchland et al., 2012; Shenoy et al., 2013; Kalaska, 2019). Imaginez, par exemple, l’enregistrement multi-unitaire de n neurones du M1. L’activité de tous les neurones est représentée par des vecteurs avec des directions différentes, mais une même origine. À tout moment, on peut donc imaginer que l’activité collective des n neurones se déplace dans un espace d’états à n dimensions. C’est essentiellement grâce à ce déplacement que le mouvement est orchestré. À noter, l’état futur du système dynamique est déterminé par son état actuel et les « inputs » qui lui parviennent (Shenoy et al., 2013). Conséquemment, le cadre théorique des systèmes dynamiques est tout à fait compatible avec celui de l’OFC, dans lequel une loi de contrôle détermine comment l’état de l’effecteur et de la cible influence le contrôleur (c.-à-d., le déplacement de l’origine dans l’espace n-dimensionnel).
1.3 Estimation d’état
Vu la section précédente, il est évident que l’estimation de l’état de l’effecteur et de la cible est un aspect fondamental pour l’OFC, car ces variables déterminent les efférences produites par le contrôleur. Intuitivement, l’information véhiculée par le sensorium (notamment la vision et la proprioception) constitue une source d’information importante pour estimer l’état de l’effecteur et de la cible dans l’environnement. Toutefois, et tel que mentionné plus haut, les réafférences sont contaminées par du bruit (neuronal, mécanique et autre,(Faisal et al., 2008), ce qui augmente l’incertitude de l’estimation d’état. De plus, les réafférences sont assujetties à des délais de transduction, de conduction et d’intégration au sein du système nerveux central. Ceci peut causer de l’instabilité pour un servomécanisme comme le cerveau, car l’implémentation d’une commande motrice en réponse à une erreur passée pourrait empirer, plutôt qu’améliorer, la performance du système (Wolpert and Miall, 1996; McNamee and Wolpert, 2019). Pour pallier ces problématiques, le cerveau utiliserait
8 un autre type de modèle, dit direct, qui utiliserait des copies des efférences motrices issues du contrôleur pour prédire les conséquences sensorielles des mouvements en cours d’exécution (ou sur le point d’être exécutés,(Wolpert et al., 1995; Wolpert and Miall, 1996; Crapse and Sommer, 2008; Shadmehr and Krakauer, 2008; Shadmehr and Mussa-Ivaldi, 2012). Selon Shadmehr and Mussa-Ivaldi (2012), le modèle direct s’apparente à un modèle génératif (probabiliste), qui établit des correspondances entre des commandes motrices et les changements d’état d’un effecteur, ainsi que des correspondances entre ces états et leurs conséquences sensorielles. En combinant ces prédictions avec les réafférences sensorielles de manière optimale au sens bayésien, le cerveau peut estimer la position de l’effecteur en temps réel et avec moins d’incertitude qu’avec les afférences/réafférences seules (Kording and Wolpert, 2004; Shadmehr and Mussa-Ivaldi, 2012; McNamee and Wolpert, 2019). Par exemple, plusieurs auteurs ont démontré que l’humain est plus rapide pour anticiper les conséquences sensorielles causées par soit comparativement à celles induits par une source externe (Flanagan and Wing, 1993; Diedrichsen et al., 2003; Nowak et al., 2007a), ce qui suggère que le système utilise les efférences motrices pour prédire les conséquences sensorielles d’actions. Évidemment, ceci implique que la détection d’évènements sensoriels non prédits par le système nerveux central est forcément en retard par rapport à leur moment d’occurrence. À titre d’exemple, l’amorce d’une correction (tâche-dépendante) en réponse à une erreur motrice nécessiterait au moins 60 ms lorsque celle-ci est détectée par le système proprioceptif et au moins 90 ms lorsqu’elle détectée par le système visuel (Scott et al., 2015; Scott, 2016).
Il est plus difficile de démontrer que l’intégration bayésienne de prédictions et de réafférences sensorielles améliore l’estimation d’état d’un effecteur en mouvement, car une fois en mouvement, l’état de l’effecteur est automatiquement estimé avec ces deux sources d’information. Par exemple, les mouvements de personnes atteintes d’une neuropathie sensorielle (c.-à-d., une perte du sens proprioceptif) sont plus variables que ceux des participants témoins (Miall et al., 2018). Malgré que ceci suggère que la proprioception est une variable importante pour l’estimation d’état, cela ne veut pas dire que les prédictions sensorielles améliorent la capacité du cerveau à localiser un effecteur en mouvement au-delà de ce qui est possible avec les réafférences seules. Pour démontrer la valeur ajoutée des
9 prédictions sensorielles pour estimation d’état, Vaziri et al. (2006), ont mis à profit l’encodage par anticipation lors de saccades oculaires. Plus précisément, lorsqu’une personne fait une saccade oculaire, le champ visuel entier est déplacé. Les éléments du champ visuel sont donc encodés par des neurones différents pré- et post-saccade. Fait intéressant, les neurones dans plusieurs régions du cerveau, notamment les cortex pariétaux (Duhamel et al., 1992), occipitaux (Nakamura and Colby, 2002), et le colliculus supérieur (Walker et al., 1995), amorcent l’encodage post-saccade avant même que l’œil n’ait bougé. Autrement dit, le cerveau prédit la prochaine position rétinienne des éléments du champ visuel en prévision du déplacement de l’œil. Conséquemment, le cerveau bénéficie de deux sources d’information pour estimer la position d’un objet après une saccade oculaire : l’encodage par anticipation (c.-à-d., la prédiction d’où devrait se trouver l’objet) ainsi que l’emplacement réel de l’objet, dicté par sa position sur la rétine. Vaziri et al. (2006) ont essentiellement montré que lors d’un pointage manuel vers une cible périphérique après une saccade oculaire (c.-à-d., prédiction + vision) le mouvement est moins variable que s’il n’est pas précédé d’une saccade (c.-à-d., vision seule). Malgré que ces résultats ne démontrent pas que les prédictions d’un modèle direct améliorent l’estimation d’état d’un effecteur, ils démontrent clairement qu’elles réduisent l’incertitude des observations faites par le sensorium.
1.3.1 Le cervelet comme siège des modèles directs
Plusieurs lignes d’évidence pointent vers le cervelet comme siège des modèles directs (Nowak et al., 2007b; Shadmehr and Krakauer, 2008; Therrien and Bastian, 2015; Ishikawa et al., 2016; Sokolov et al., 2017). En effet, des lésions cérébelleuses, autant réelles que virtuellement induites avec des appareils de neurostimulation, provoquent des déficits moteurs s’apparentant à une incapacité à anticiper les conséquences sensorielles de nos actions (Miall et al., 2007; Nowak et al., 2007a). Par exemple, le fait de perturber temporairement le cervelet pendant l’exécution d’un mouvement du membre supérieur provoque un retard dans l’estimation de la position du membre (Miall et al., 2007). Outre les évidences comportementales, la connectivité du cervelet, ainsi que le moment d’activation des neurones dans ses secteurs moteurs, répondent aux prérequis d’un modèle direct
10 (Ishikawa et al., 2016). Dans un premier temps, le cervelet reçoit des projections proprioceptives et cutanées des voies spinocérébelleuses (Oscarsson, 1965; Cooke et al., 1971; Ekerot and Larson, 1972). Ceci lui procure l’information nécessaire pour connaitre la configuration du corps avant et pendant un mouvement. Deuxièmement, le cervelet a des connexions réciproques avec le cortex moteur primaire (Kelly and Strick, 2003; Lu et al., 2007) qui pourrait véhiculer les copies d’efférences permettant l’estimation du prochain état sensoriel de l’appareil locomoteur. Finalement, lors de mouvements du membre supérieur, les fibres moussues qui projettent vers les régions motrices du cervelet s’activent après le cortex moteur primaire, mais avant l’initiation du mouvement (van Kan et al., 1993; Ishikawa et al., 2014). En l’occurrence, il est plausible que cette activité reflète l’arrivée de copies d’efférences au cervelet ou encore la formulation de prédictions sensorielles par le modèle direct.
1.3.2 Le cortex pariétal comme estimateur d’état
Si le cervelet s’occupe de prédire les conséquences sensorielles des mouvements à venir, quelles régions du cerveau pourraient s’occuper de les combiner aux réafférences sensorielles pour estimer l’état de l’effecteur en mouvement? À la lumière du paragraphe précédent, le cervelet semble aussi être un bon candidat. En effet, en plus de recevoir des projections sensorielles provenant de la périphérie (Oscarsson, 1965; Cooke et al., 1971; Ekerot and Larson, 1972), le cervelet reçoit des projections visuelles (Glickstein et al., 1972; Glickstein et al., 1994; Kralj-Hans et al., 2007). Conséquemment, si on accepte qu’il soit le siège du modèle direct (ce qui implique qu’il a accès aux prédictions sensorielles), le cervelet détiendrait tous les éléments nécessaires pour formuler une estimation de l’état du corps dans l’espace. Néanmoins, les modèles actuels en contrôle moteur suggèrent que le cortex pariétal, plutôt que le cervelet, soit le siège de l’estimation d’état (Shadmehr and Krakauer, 2008). L’étude de cas de la patiente PJ est peut-être la plus convaincante du rôle joué par le cortex pariétal dans l’estimation de l’état du corps dans l’espace (Wolpert et al., 1998). Cette patiente, atteinte d’une lésion importante au lobe pariétal supérieur gauche, rapportait avoir l’impression que son bras droit disparaissait graduellement si elle le perdait de vue. Dans la même veine, Rushworth et al. (1997) ont montré que des lésions aux lobes pariétaux
11 supérieurs et inférieurs provoquent des déficits moteurs lors de mouvements de pointage chez le macaque. Remarquablement, ces lésions ne semblent pas avoir d’effets délétères lors de tâches motrices ne requérant pas le guidage d’un effecteur vers une cible (p.ex. tirer sur un levier en réponse à un stimulus). Somme tout, ces résultats suggèrent que les lésions pariétales affectent les transformations sensorimotrices à une étape qui précède celle de l’implémentation de commandes motrices, tel que la mise à jour de la représentation du corps dans l’espace (c.-à-d., l’estimation d’état).
Comme le cervelet, le cortex pariétal semble détenir tous les éléments nécessaires pour estimer l’état du corps dans l’espace. Plus précisément, les cortex pariétaux reçoivent des projections visuelles (Baizer et al., 1991; Galletti et al., 2001) et proprioceptives (Prevosto et al., 2011). De plus, il semble que le cortex pariétal ait accès aux prédictions sensorielles du modèle direct. En effet, Mulliken et al. (2008) ont démontré que le cortex pariétal postérieur du macaque encode la position passée, présente et future du membre supérieur lors de mouvements de pointage, ce qui serait impossible sans l’accès aux prédictions sensorielles permettant l’anticipation du mouvement. Donc, selon les théories actuelles, le cortex pariétal recevrait les prédictions sensorielles du cervelet, possiblement via des projections issues des noyaux cérébelleux profonds (Prevosto et al., 2010), qu’il intégrerait avec les réafférence sensorielles pour estimer l’état du corps dans l’espace (Shadmehr and Krakauer, 2008).
C
HAPITRE2 :
L’
ADAPTATION SENSORIMOTRICEL’adaptation sensorimotrice est un type d’apprentissage moteur qui permet l’optimisation d’habiletés motrices déjà existantes (Krakauer et al., 2019). Pour l’étudier, on utilise des perturbations sensorimotrices qui altèrent la correspondance habituelle entre les commandes motrices et les changements d’états attendus. À la lumière de l’OFC, la perturbation sensorimotrice force l’apprenant à remédier à deux problématiques s’il veut optimiser son comportement. Premièrement, il doit modifier sa loi de contrôle, car celle-ci ne pointe plus vers les commandes motrices optimales. Deuxièmement, il doit mettre à jour
12 son modèle direct, car les prédictions issues de ce dernier sont cruciales pour que le système puisse évaluer l’état de l’effecteur en temps réel et avec certitude pendant l’exécution du mouvement. Conséquemment, les protocoles d’adaptation sensorimotrice offrent une vitrine qui donne sur l’optimisation du contrôle rétroactif (c.-à-d., comment FC devient OFC).
2.1 Méthodes pour investiguer l’adaptation sensorimotrice
En laboratoire, on note deux grandes familles de perturbations sensorimotrices. L’une d’entre elles consiste à altérer la dynamique de nos mouvements en entravant leur exécution avec des forces externes (Shadmehr, 2017). Ce type de perturbation peut être produit par une plateforme rotative induisant des forces de Coriolis (Lackner and Dizio, 1994), ou encore par des robots articulés permettant de simuler un champ de force (Shadmehr and Mussa-Ivaldi, 1994). Dans tous les cas, en altérant les forces qui agissent sur l’effecteur, la perturbation altère la correspondance habituelle entre une cinématique intrinsèque désirée et les torques articulaires nécessaires pour l’accomplir. La deuxième famille de perturbations consiste à altérer la correspondance habituelle entre la cinématique extrinsèque et intrinsèque des mouvements. Ceci peut être accompli en déplaçant, de façon subliminale, une cible à atteindre en cours de mouvements. Par exemple, en déplaçant la cible d’une saccade pendant l’accomplissement de celle-ci, il est possible d’adapter l’amplitude des saccades ultérieures (McLaughlin, 1967). Cette même méthode peut être utilisée pour adapter les mouvements du membre supérieur (Magescas and Prablanc, 2006; Cameron et al., 2010). Une autre méthode consiste à manipuler le retour visuel des mouvements exécutés plutôt que de berner les participants en déplaçant la cible. La plus ancienne méthode utilisant cette tactique est le port de lunettes prismatiques (Stratton, 1897), qui déplacent ou inversent le champ visuel en déviant la lumière sur la rétine. Malgré que les lunettes prismatiques soient encore utilisées aujourd’hui (Striemer et al., 2019), l’arrivée des ordinateurs permet de manipuler le retour visuel des mouvements de façon « virtuelle ». Par exemple, on peut implémenter un gain de position à un curseur sur un écran, augmentant ainsi la distance qu’il parcourt par rapport à la main de l’apprenant (Krakauer et al., 2000). Une autre méthode, appelée rotation visuomotrice, consiste à induire un décalage angulaire entre la position de la main de l’apprenant et celle du curseur la représentant (Prablanc et al., 1975). Dans tous les cas, pour
13 retrouver un niveau de performance optimal (c.-à-d., préperturbation), l’apprenant doit, lors d’une phase d’acquisition, adapter ses mouvements de façon à contrer la perturbation.
2.2 Les processus explicite et implicite en adaptation sensorimotrice
Une courbe d’adaptation normale s’apparente à une fonction logarithmique, et montre essentiellement une amélioration monotone de la performance (c.-à-d., une réduction des erreurs) en fonction de la pratique. Après avoir observé que les post-effets induits par l’adaptation prismatique étaient inférieurs aux changements de comportement pendant la période d’acquisition, Redding and Wallace (1996) ont suggéré que deux processus contribuent à l’adaptation sensorimotrice. L’un d’eux correspond à l’implémentation d’une stratégie cognitive (p.ex. viser à l’opposé de la déviation) alors que l’autre correspondrait au réalignement des cartes sensorimotrices (parfois appelé le « vrai apprentissage »). Toutefois, la démonstration directe de cette hypothèse est difficile via l’adaptation prismatique, car l’apprenant voit sa main réelle, plutôt qu’un curseur virtuel la représentant. En effet, ce n’est qu’en utilisant la rotation visuomotrice que les travaux des laboratoires de Richard B. Ivry et de Jordan A. Taylor, ont permis de quantifier la contribution de ces deux processus pendant l’adaptation (Taylor et al., 2014; Bond and Taylor, 2015; McDougle et al., 2015; Morehead et al., 2015).
Dans la première de ces études, Taylor et al. (2014) demandaient à des participants de leur indiquer, à l’aide de repères visuels positionnés sur l’espace de travail, où ils visaient avant chaque essai d’un protocole d’adaptation à une rotation visuomotrice. Tel que suggéré par Redding and Wallace (1996), les participants rapportaient viser volontairement dans la direction opposée à la perturbation plutôt que la cible à atteindre pendant la phase d’acquisition. Ainsi, ces résultats mettent clairement en évidence l’identification et l’implémentation d’une stratégie motrice explicite permettant à l’apprenant de contrecarrer une perturbation sensorimotrice rapidement. Toutefois, ces résultats montrent qu’à elle seule, la stratégie motrice n’explique pas complètement le changement de comportement durant la phase d’acquisition. Taylor et al. (2014) ont donc raisonné que la fraction du comportement moteur non attribuable à l’implémentation de la stratégie devait être le fruit du deuxième
14 processus correspondant à la recalibration du modèle direct. Pour quantifier la contribution de cette recalibration au changement de performance pendant l’acquisition, ils ont simplement soustrait la fraction du comportement moteur attribuable à la stratégie au comportement moteur mesuré. On désigne aujourd’hui « d’explicite » la fraction de l’adaptation attribuable à l’implémentation d’une stratégie cognitive et « d’implicite » celle attribuable à la recalibration du modèle direct (McDougle et al., 2015).
Une fois les processus explicite et implicite isolés, deux conclusions principales peuvent être tirées quant à leurs contributions respectives en contexte d’adaptation sensorimotrice. Premièrement, l’adaptation explicite contribue largement plus à l’amélioration de la performance en début d’acquisition que l’adaptation implicite (Smith et al., 2006; Taylor et al., 2014). D’ailleurs, Bond and Taylor (2015) ont montré qu’en contexte d’adaptation visuomotrice, la taille de la rotation imposée détermine l’amplitude de la stratégie explicite utilisée par l’apprenant, mais pas le taux d’adaptation implicite. Ce dernier cesserait de croître à partir de rotations visuomotrices d’environ 6° (Morehead et al., 2017; Kim et al., 2018). On pourrait donc respectivement qualifier les processus explicite et implicite de « rapide » et de « lent » (Smith et al., 2006). Deuxièmement, malgré sa contribution importante en début d’adaptation, l’implication du processus explicite tend à diminuer au fil des essais, alors que celle du processus implicite croît de façon monotonique. Ceci suggère que l’adaptation explicite est un phénomène flexible qui s’articule autour de l’adaptation implicite pour maximiser la performance (Taylor and Ivry, 2011; Bond and Taylor, 2015).
Selon Schmidt and Lee (2011), un apprentissage devrait laisser une trace relativement permanente chez un apprenant. À la lumière de ce postulat, l’adaptation sensorimotrice devrait laisser une trace au sein du système sensorimoteur une fois la perturbation retirée. La trace d’apprentissage la plus notable en adaptation sensorimotrice est de loin les changements de comportements moteurs observés lorsqu’on retire la perturbation suite à la période d’acquisition. Ces « post-effets » se traduisent par des mouvements avec un biais directionnel suite à l’adaptation visuomotrice (p.ex.(Canaveral et al., 2017), et par la production de forces compensatoires pour des perturbations dynamiques (p.ex.(Scheidt et
15 al., 2000). À noter, ces post-effets s’observent malgré que l’apprenant sache que la perturbation a été retirée, ce qui suggère que de réels changements ont pris place dans son système nerveux central. La magnitude des post-effets est généralement proportionnelle au niveau d’adaptation implicite atteint pendant l’adaptation (Taylor et al., 2014; Bond and Taylor, 2015). Conséquemment, ils sont généralement considérés comme un reflet pur de la mise à jour du modèle direct (Taylor et al., 2014).
À la lumière de ce qui précède, il est évident que le processus explicite représente une modification de la loi de contrôle, car il provoque des changements de comportement évidents. On pourrait aussi conclure que le processus implicite contribue à la modification de la loi de contrôle, car il provoque aussi des changements de comportement moteur. Une possibilité intéressante, toutefois, est que contrairement au processus explicite, le processus implicite pourrait influencer la loi de contrôle indirectement. En effet, en modifiant l’estimation d’état essai par essai (via la mise à jour du modèle direct), le processus implicite pourrait rendre sous-optimale toute modification de la loi de contrôle basée sur une erreur de tâche antérieure. Ceci expliquerait d’ailleurs pourquoi le processus explicite semble accommoder le processus implicite pendant l’adaptation à une perturbation sensorimotrice (Taylor and Ivry, 2011; Bond and Taylor, 2015).
2.3 L’adaptation sensorimotrice est stimulée par les erreurs
Pour améliorer sa performance en contexte d’adaptation sensorimotrice, le cerveau doit modifier sa loi de contrôle ainsi que l’estimation qu’il fait de l’état de l’effecteur pendant le mouvement (qu’il accomplit, notamment, en modifiant son modèle direct). Pour y arriver, il doit être informé que sa loi de contrôle/estimation d’état est sous-optimale/erronée. Ce sont les erreurs commises lors de l’exécution motrice qui procurent cette information au cerveau.
2.3.1 Erreurs explicites
Lorsque confrontée à une perturbation sensorimotrice, la loi de contrôle utilisée par l’apprenant n’est plus optimale, car elle ne pointe plus vers les bonnes transformations
16 sensorimotrices. Conséquemment, l’apprenant n’arrive (généralement) plus à atteindre l’objectif moteur. Ceci provoque ce qu’on appelle une « erreur de prédiction de récompense (EPR) » (c.-à-d., des rétroactions binaires indiquant « succès/échec ») (Izawa and Shadmehr, 2011; Walsh and Anderson, 2012; Nikooyan and Ahmed, 2015; Schultz, 2016). Ce type d’erreur est primordial pour l’adaptation du comportement moteur, car il indique à l’apprenant qu’il doit changer sa loi de contrôle pour améliorer sa performance. Étant donné qu’elles sont de nature « scalaire » (Izawa and Shadmehr, 2011; Shmuelof et al., 2012; Krakauer et al., 2019), les EPR procurent peu d’information concernant la façon dont l’apprenant devrait changer son comportement pour améliorer sa performance. Conséquemment, si l’apprenant n’a accès qu’aux EPR pour identifier une nouvelle loi de contrôle, l’adaptation sera longue et les mouvements plus variables (Izawa and Shadmehr, 2011; Shmuelof et al., 2012; Haith and Krakauer, 2013). Ce type d’adaptation tombe essentiellement sous l’ombrelle de l’apprentissage par renforcement (Sutton and Barto, 2018). Dans ce schème, le contrôleur (M1/prémoteur) apprend directement une loi de contrôle (Haith and Krakauer, 2013), probablement en associant des mouvements à des récompenses (Nikooyan and Ahmed, 2015). À noter, malgré que les EPR seules suffisent pour adapter un comportement moteur, elles ne donnent pas lieu à une recalibration des cartes sensorimotrices (Izawa and Shadmehr, 2011; Nikooyan and Ahmed, 2015). Conséquemment, elles semblent contribuer à l’adaptation explicite, mais pas (ou peu) à l’adaptation implicite (mais voir section 2.3.2).
Lorsque l’apprenant a accès aux retours sensoriels de ses mouvements (ce qui est généralement le cas dans des situations écologiques), l’apprenant pourra déterminer l’amplitude et la direction de l’erreur motrice qui a causé l’EPR (Shmuelof et al., 2012). Plus précisément, en déterminant l’écart entre la position finale du mouvement qu’il vient d’exécuter et la cible, il pourra utiliser un vecteur d’erreur qu’on appelle « erreur de tâche » (Krakauer et al., 2019) ou encore « erreur de cible » (Taylor et al., 2014) pour identifier la nature de la perturbation. Ce faisant, il pourra plus rapidement modifier sa loi de contrôle pour compenser pour la perturbation. Selon plusieurs auteurs, l’information tirée des erreurs de tâche, comme les EPR, nourrit le processus d’adaptation explicite (Mazzoni and
17 Krakauer, 2006; Taylor and Ivry, 2014; Taylor et al., 2014; Krakauer et al., 2019), permettant ainsi à l’apprenant de compenser rapidement pour des perturbations sensorimotrices.
2.3.2 Erreurs de prédiction sensorielles (EPS)
Étant donné que les prédictions du modèle direct deviennent erronées lorsqu’un apprenant est soumis à une perturbation sensorimotrice, sa capacité à estimer l’état d’un effecteur en mouvement est diminuée. Heureusement, le cerveau peut utiliser ses erreurs de prédiction (c.-à-d., l’écart entre le mouvement désiré et celui effectué (Mazzoni and Krakauer, 2006; Shadmehr et al., 2010a; Taylor and Ivry, 2014) pour ajuster ses prédictions ultérieures (c.-à-d., modifier son modèle direct). Malgré que toutes les erreurs de prédiction soient le fruit d’afférences sensorielles erronément prédites (y compris les EPR), on appelle les erreurs de prédictions du modèle direct les « erreurs de prédictions sensorielles (EPS) ». À l’opposé des erreurs de tâche, les EPS semblent être à l’origine de l’adaptation implicite et des post-effets qu’elle engendre. En effet, malgré que l’adaptation implicite puisse être observée suite à une erreur de tâche causée par le déplacement non perçu d’une cible (Magescas and Prablanc, 2006; Cameron et al., 2010), elle s’observe généralement en réponse aux EPS induites par une perturbation sensorimotrice. D’ailleurs, des post-effets peuvent être observés ne serait-ce qu’après un seul essai contraint à une perturbation sensorimotrice (Diedrichsen et al., 2005; Torrecillos et al., 2015). En contrepartie, peu ou pas de post-effets sont observables suite à l’erreur de tâche induite par un saut de cible perçu (Diedrichsen et al., 2005; Torrecillos et al., 2015), probablement parce que l’erreur n’occasionne pas d’EPS.
En début d’adaptation, les EPS et les erreurs de tâche sont confondues du fait que leur amplitude est idem. Néanmoins, ces deux types d’erreurs semblent relativement indépendants. Ceci fut démontré dans une étude de Mazzoni and Krakauer (2006), dans laquelle des participants devaient adapter leurs mouvements à une rotation visuomotrice de 45. À la différence d’autres études d’adaptation, les auteurs dévoilaient aux participants comment contrecarrer la rotation à l’aide d’une stratégie (ou loi de contrôle) qui consistait à viser la cible voisine (positionnée à -45 par rapport à la cible à atteindre). Mazzoni and
18 Krakauer (2006) ont d’abord noté que l’implémentation de la stratégie permettait aux participants d’éliminer complètement les erreurs de tâche en début d’adaptation. Au fil des essais, toutefois, ces derniers se mettaient graduellement à faire des erreurs de tâche dans la direction opposée à la perturbation (c.-à-d., ils surcompensaient), minant ainsi leur performance. Vu son indépendance vis-à-vis les erreurs de tâche, les auteurs ont suggéré que cette surcompensation devait être attribuable à l’adaptation du modèle direct causée par l’EPS perçu entre la trajectoire du curseur (vers la cible désirée) et celle de la main (vers la cible voisine). Ces données, maintenant répliquées à plusieurs reprises (Taylor et al., 2010; Taylor and Ivry, 2011; Schween et al., 2014; Rand and Rentsch, 2015), suggèrent que contrairement à son homologue explicite, qui est tributaire des erreurs de tâche, l’adaptation implicite du modèle direct dépend des EPS.
Malgré que les évidences actuelles suggèrent que les EPS et les erreurs de tâche sont relativement indépendantes, des données récentes suggèrent que les EPR modulent la sensibilité du système moteur aux EPS (Gaveau et al., 2014; Leow et al., 2018; Kim et al., 2019). Par exemple, Kim et al. (2019) ont utilisé un protocole d’adaptation visuomotrice où ils demandent aux participants de pointer des cibles tout en ignorant un curseur qui se déplace en même temps que leur main, mais systématiquement à 3.5 de la cible à atteindre. Lorsque confronté à ce type de retour visuel dit « clampé », la direction des mouvements produits par les participants dérive dans le sens opposé à la perturbation, probablement parce que la perturbation induit des EPS invariables qui provoquent une adaptation implicite du modèle direct (Morehead et al., 2017; Kim et al., 2018). Dans leur étude, Kim et al. (2019) ont modifié la taille des cibles de façon à ce que le curseur clampé les rate complètement, les frôle ou les frappe complètement lors de mouvements produits par les participants. Étonnement, ils montrent que malgré que le curseur soit clampé à 3.5 dans toutes les conditions, l’adaptation implicite est nettement supérieure lorsque le curseur rate ou frôle les cibles comparativement à lorsqu’il les frappe. De plus, l’absence de différence d’adaptation implicite pour les conditions « ratée » et « frôlée » suggère que cet effet est binaire, et donc tributaire de l’EPR plutôt que l’erreur de tâche. Dans tous les cas, il est à noter que l’adaptation implicite a eu lieu peu importe que le curseur rate, frôle ou frappe la cible, ce
19 qui suggère que l’adaptation implicite est modulée, plutôt qu’abolit, par l’absence d’EPR (c’est aussi ce que suggère l’étude de(Mazzoni and Krakauer, 2006).
2.4 Substrats potentiels pour l’adaptation explicite/implicite
Étant donné leurs présumés rôles dans l’élaboration de prédictions sensorielles/estimation d’état en contrôle moteur, le cervelet et le cortex pariétal semblent des substrats prometteurs pour prendre en charge l’adaptation sensorimotrice. En effet, les caractéristiques qui font de ces régions de bons candidats pour l’estimation d’état (c.-à-d., l’accès aux afférences visuelles, proprioceptives, et aux prédictions sensorielles), en font aussi de bons candidats pour détecter/traiter les erreurs de tâche et les EPS qui contribuent respectivement à l’adaptation explicite et implicite.
2.4.1 Le cervelet
Le cervelet est peut-être la région la plus souvent impliquée dans l’adaptation sensorimotrice (et l’apprentissage, globalement). En effet, plusieurs études de neuroimagerie ont montré une augmentation d’activité dans différents secteurs du cervelet en réponse à diverses perturbations sensorimotrices (Shadmehr and Holcomb, 1997; Imamizu et al., 2000; Krakauer et al., 2004; Graydon et al., 2005; Seidler et al., 2006; Danckert et al., 2008; Luaute et al., 2009; Chapman et al., 2010b; Schlerf et al., 2012; Werner et al., 2014). Par ailleurs, plusieurs études ont montré que des personnes avec une atteinte cérébelleuse démontrent des difficultés d’adaptation sensorimotrice (Martin et al., 1996; Pisella et al., 2005; Smith and Shadmehr, 2005; Tseng et al., 2007; Synofzik et al., 2008; Rabe et al., 2009; Werner et al., 2009; Criscimagna-Hemminger et al., 2010; Taylor et al., 2010; Werner et al., 2010; Donchin et al., 2012; Izawa et al., 2012; Gibo et al., 2013; Butcher et al., 2017; Morehead et al., 2017). Par exemple, Butcher et al. (2017), ont montré que des participants atteints d’ataxie spinocérébelleuse ont peine à identifier une stratégie pour contrer une rotation visuomotrice et ce, malgré qu’ils soient capables d’appliquer une stratégie donnée par un agent externe (Taylor et al., 2010). Malgré son rôle dans l’adaptation explicite, on associe surtout le cervelet à l’adaptation implicite. Par exemple, Tseng et al. (2007) ont montré que les
post-20 effets, suite à un protocole d’adaptation visuomotrice, sont atténués chez des participants atteints d’ataxie spinocérébelleuse comparativement à des participants témoins. Par ailleurs, il a été montré que la dérive de la main typiquement observée lorsque l’on donne une stratégie explicite à des participants en contexte d’adaptation visuomotrice (voir(Mazzoni and Krakauer, 2006) est réduite chez des personnes atteintes d’ataxie (Taylor et al., 2010). Enfin, d’autres études ont montré que le décours temporel d’adaptation implicite est inférieur chez des personnes atteintes d’ataxie (Butcher et al., 2017; Morehead et al., 2017). Pris avec les résultats de neuroimagerie décrits ci-haut, ces évidences suggèrent fortement que le cervelet joue un rôle de premier plan dans l’adaptation implicite.
Malgré que son rôle dans l’adaptation implicite soit largement reconnu, quelques études ont montré que des personnes avec une atteinte cérébelleuse sont capables de contrer, au même titre qu’un groupe témoin, une perturbation sensorimotrice graduellement introduite (Criscimagna-Hemminger et al., 2010; Izawa et al., 2012; Henriques et al., 2014). Par exemple, Izawa et al. (2012) ont montré que des personnes présentant une atteinte cérébelleuse sont capables de s’adapter à une rotation visuomotrice graduellement imposée dans une tâche de pointage avec une seule cible. Par contre, ces participants montraient peu ou pas de post-effets comparativement aux participants témoins, ce qui suggère que l’adaptation n’était probablement pas implicite. D’autres études, toutefois, montrent que les personnes présentant des lésions cérébelleuses ont des post-effets comparables à ceux de participants témoins suite à un protocole d’adaptation graduel, ce qui suggère que la capacité d’adaptation à de petites erreurs pourrait être le fruit du processus implicite (Criscimagna-Hemminger et al., 2010; Henriques et al., 2014). Il est à noter, toutefois, que dans l’étude de Henriques et al. (2014), les patients présentaient des scores d’ataxie relativement faible sur l’échèle SARA (Scale for the Assessment and Rating of Ataxia). Conséquemment, il est possible que la gravité de l’atteinte cérébelleuse n’ait pas été suffisamment importante pour avoir un impact détectable sur l’adaptation. Quant aux résultats de Criscimagna-Hemminger et al. (2010), qui montrent que les personnes avec une atteinte cérébelleuse sont capables de s’adapter à un champ de force graduel dans le sens horaire, mais pas à un champ de force abrupt dans le sens antihoraire, ils pourraient être le fruit d’un artéfact méthodologique. En effet, Gibo et al. (2013) ont montré qu’en contexte d’adaptation à un champ de force, la
21 capacité d’adaptation de personnes avec une atteinte cérébelleuse pourrait dépendre des déficits moteurs initiaux des participants. Dans cette étude, les auteurs ont montré que les patients avaient de la difficulté à contrer les forces nécessaires pour déplacer le robot utilisé pour la collecte de données dans un champ de force nul. Conséquemment, lorsque la direction de la perturbation compensait (fortuitement) les forces intrinsèques du robot, la performance des patients s’améliorait, et ce, peu importe si la perturbation était implémentée, de manière abrupte ou graduelle (Gibo et al., 2013). Conséquemment, les résultats de Criscimagna-Hemminger et al. (2010) pourraient être expliqués par le fait que la direction du champ de force utilisé pour l’adaptation graduelle et abrupte n’ait pas été contrebalancée. Pris ensemble, ces résultats suggèrent que l’assistance du cervelet n’est peut-être pas nécessaire pour contrer une perturbation sensorimotrice graduellement imposée (c.-à-d., améliorer sa performance pendant la phase d’acquisition). Toutefois, le cervelet semble être nécessaire pour que l’adaptation implicite prenne place (c.-à-d., pour qu’il y ait une mise à jour d’un modèle direct).
Certaines études avec des patients présentant des lésions cérébelleuses ont permis d’identifier les secteurs du cervelet causalement impliqués dans l’adaptation sensorimotrice. Par exemple, les lésions touchant les régions plutôt antérieures du cervelet (lobes IV et V) affecteraient davantage l’adaptation à un champ de force que l’adaptation visuomotrice, alors que les lésions rostrales du cervelet postérieur (lobe VI) affecteraient principalement l’adaptation visuomotrice (Rabe et al., 2009; Donchin et al., 2012). En accord avec ces résultats, les lésions touchant l’artère cérébelleuse supérieure, qui irrigue la moitié supérieure du cervelet (donc les lobes IV, V et VI) auraient des impacts délétères plus importants sur l’adaptation sensorimotrice que des lésions à son homologue postérieure (Werner et al., 2009; Werner et al., 2010; Donchin et al., 2012)1. À noter, les études de Rabe et al. (2009)
et Donchin et al. (2012) montrent aussi que, chez les patients, la performance lors d’une tâche d’adaptation visuomotrice et à un champ de force n’est pas significativement corrélée, ce qui suggère que ces tâches pourraient mettre en jeu des régions relativement distinctes du cervelet (mais voir(Diedrichsen et al., 2005).