ANALYSE DU PROCESSUS DE RENOUVELLEMENT PAR TROUÉES DE LA FORÊT FEUILLUE TEMPÉRÉE NATURELLE ET JARDINÉE
THÈSE PRÉSENTÉE
COMME EXIGENCE PARTIELLE DU DOCTORAT EN BIOLOGIE
PAR
JEAN-FRANÇOIS SÉNÉCAL
UNIVERSITÉ DU QUÉBEC À MONTRÉAL Service des bibliothèques
Avertissement
La diffusion de cette thèse se fait dans le respect des droits de son auteur, qui a signé le formulaire Autorisation de reproduire et de diffuser un travail de recherche de cycles
supérieurs (SDU-522 – Rév.07-2011). Cette autorisation stipule que «conformément à
l’article 11 du Règlement no 8 des études de cycles supérieurs, [l’auteur] concède à l’Université du Québec à Montréal une licence non exclusive d’utilisation et de publication de la totalité ou d’une partie importante de [son] travail de recherche pour des fins pédagogiques et non commerciales. Plus précisément, [l’auteur] autorise l’Université du Québec à Montréal à reproduire, diffuser, prêter, distribuer ou vendre des copies de [son] travail de recherche à des fins non commerciales sur quelque support que ce soit, y compris l’Internet. Cette licence et cette autorisation n’entraînent pas une renonciation de [la] part [de l’auteur] à [ses] droits moraux ni à [ses] droits de propriété intellectuelle. Sauf entente contraire, [l’auteur] conserve la liberté de diffuser et de commercialiser ou non ce travail dont [il] possède un exemplaire.»
Je tiens d'abord à remercier chaudement ma compagne, Audrey qui m ' a soutenu durant mes études graduées. Sans sa grande patience, cette thèse n'aurait pas pu être menée à terme. Je remercie aussi toute ma famille élargie qui a fourni beaucoup de support au
fil
des ans sans trop poser de questions.Merci à mon directeur de thèse, Frédérik Doyon pour m 'avoir fait confiance encore une fois pour un nouveau projet. Ton enthousiasme et tes nombreuses idées ont certainement contribué à rendre ces dernières années intéressantes! Merci aussi à mon codirecteur de thèse, Christian Messier, qui malgré son horaire toujours chargé a pris beaucoup de temps pour tenter de m'inculquer ce qu'est un chercheur ( et aussi ce que ce n' est pas). La simplicité et la complexité de ton discours m 'a beaucoup inspiré.
Merci aussi aux professeurs, au personnel et aux étudiants de l'Isfort. C'est certainement le meilleur environnement de travail/étude qui soit; surtout en raison des gens qu'on y retrouve. J'adresse un merci particulier à Angélique Dupuch pour l'aide précieuse en statistiques et son constant sourire chaque fois que j 'allais lui poser une dernière question (souvent trois-quatre fois de suite).
LISTE DES FIGURES ... IX LISTE DES TABLEAUX ... XIII RÉSUMÉ ... .... ... ... ... ... . -XV
INTRODUCTION ... ... 1
0.1 Définition des trouées ... 1
0.2 Dynamique des trouées ... ... ... 3
0.3 Autres types d'ouvertures de la canopée ... ... 5
0.4 Effets des trouées sur la croissance des arbres ... ... ... .. 7
0.5 Forêts aménagées ... 9
0.6 Lidar ... ... ... ... 10
0.7 Objectifs de l'étude ... 11
CHAPITRE! MANAGEMENT IMPLICATIONS OF V ARYING GAP DETECTION HEIGHT THRESHOLDS AND OTHER CANOPY DYNAMICS PROCESSES IN TEMPERATE DECIDUOUS FORESTS ... ... 13
1.1 Abstract ... 15
1.2 Introduction ... 17
1.3 Study area and Methods ... ... . 19
1.3.1 Study area ... 19
1. 3 .2 LiD AR data processing ... 20
1.3.3 Canopy dynamics processes ... 22
1.4 Results ... 24
1.4.1 Mean canopy height .. ... ... ... ... 24
1.4.2 Canopy processes ... 26
1.4.4 Gap extinction rate ... 29
1.5 Discussion ... ... 29
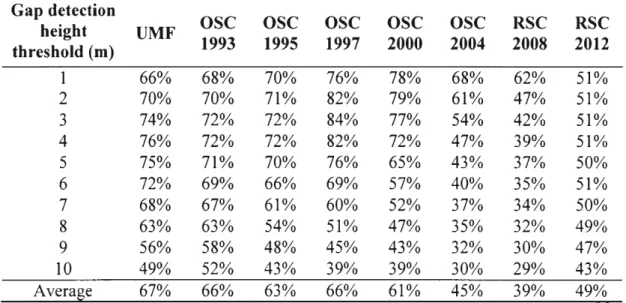
1.5.1 Canopy dynamics processes and gap detection height treshold ... 29
1.5.2 New gap formation and canopy erosion ... 31
1.5.3 Canopy dynamics in managed forests ... 32
1.6 Conclusion ... ... 35
CHAPITRE II DISENTANGLING THE CAUSES OF CANOPY HEIGHT INCREASE IN MANAGED AND UNMANAGED TEMPERATE DECIDUOUS FORESTS USING MULTI-TEMPORAL LIDAR DATA ... ... 37
2.1 Abstract ... 39
2.2 Introduction ... .. ... 41
2.3 Method ... ... ... ... 43
2.3.1 Study area ... ... ... ... 43
2.3.2 LiDAR data processing ... .. ... 44
2.3.3 Canopy height change classification ... 45
2.3 .4 Vertical growth rate analysis ... 4 7 2.4 Results ... 49
2.4.1 Estimated density of LiDAR points per pixels in the 2007 and 2013 canopy height models ... 49
2.4.2 Chronosequence of canopy processes .... ... 49
2.4.3 Canopy processes description ... ... 50
2.4.4 Vertical growth rate analysis ... 52
2.5 Discussion ... 55
2.6 Conclusion ... 63
CHAPITRE III WHAT CAUSES GAPLESS TREE DEATH: AN INVESTIGATION OF CANOPY DYNAMICS OF NORTHERN TEMPERATE DECIDUOUS FORESTS3 ... 65
3.1 Abstract ... .' ... 67
3 .2 Introduction ... 69
3.3 Methods ... ... ... ... ... 71
3.3.2 LiDAR data processing ... ... 72
3 .3 .3 Field sampling ... 73
3.4 Statistical analysis ... 75
3.5 Results ... ... ... : ... 77
3 . 5 .1 S tatisti cal anal y sis ... · ... 77
3.5.2 Tree damage mechanisms ... 78
3.5.3 Canopy structure ... .... 78
3.6 Discussion ... 80
3.6.1 Tree damage mechanisms ... 80
3 .6.2 Best explanation of canopy gap presence ... 85
3 .6.3 Implications ... 85
3.7 Conclusion ... 87
CONCLUSION GÉNÉRALE ... 89
3.8 Retour sur les résultats ... 89
3.8.1 Cadre conceptuel de la dynamique des trouées ... 89
3. 8 .2 Processus de canopée ... 94
3.8.3 Mécanismes de dégâts des arbres versus structure ... 96
3.8.4 Rétablissement de la canopée dans les forêts aménagées et non-aménagées ... 98
3.8.5 Derniers mots ... ... ... ... ... 104
3. 9 Perspectives de recherche ... 1 0 5 3.9.1 Suite à donner à la modélisation du comblement latéral.. ... 105
3.9.2 Succès de la régénération du bouleau jaune en peuplements jardinés .. . 106
3. 9 .3 Distribution de tailles ... ... 107
3.9.4 Végétation statique près du sol.. .... ... ... ... ... ... .. 108
3.9.5 Facteurs affectant la répartition spatiale des classes de structure de la canopée ... 11 0 APPENDICE A SUPPLEMENTARY MATERIAL TO CHAPTER 1 ... 113 APPENDICEB
APPENDICE C
SUPPLEMENTARY MATERJAL TO CHAPTER 3 ... 121 LISTE DES RÉFÉRENCES ... 125
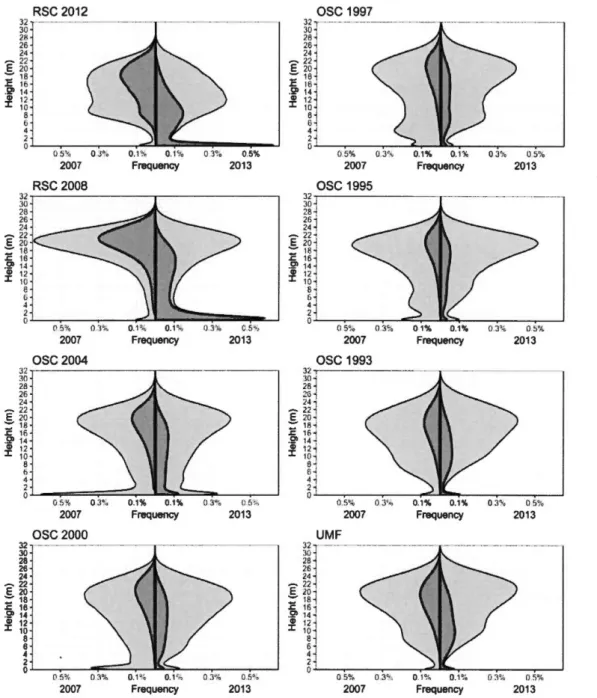
Figure Page Figure 1.1 Change in canopy height distribution of the study sites from 2007 (left
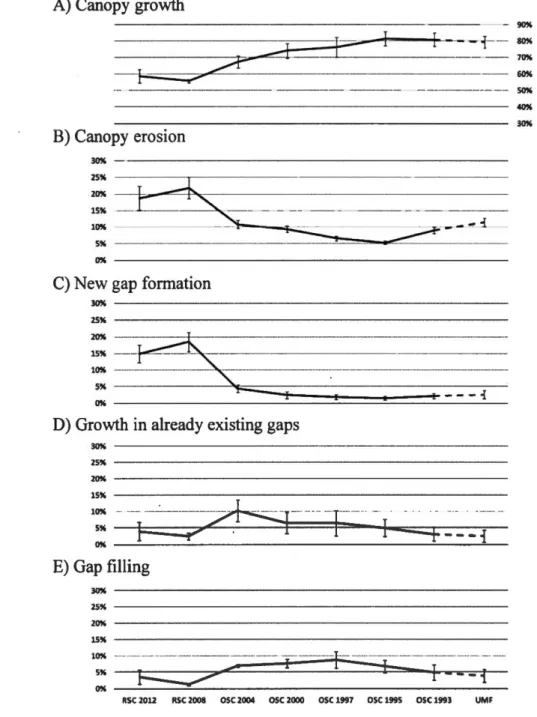
of each graph) to 2013 (right of each graph) by canopy processes (dark grey: height reduction > 1 m; light gray: height increases). RSC refers to recent selection cuts-stands that were eut between 2007 and 2013. OSC refers to old selection cuts-stands that were eut the given year, before 2007. UMF refers to unmanaged forests-forests that have never been eut. ... 27 Figure 1.2 Mean of canopy processes frequencies ( canopy growth, canopy erosion,
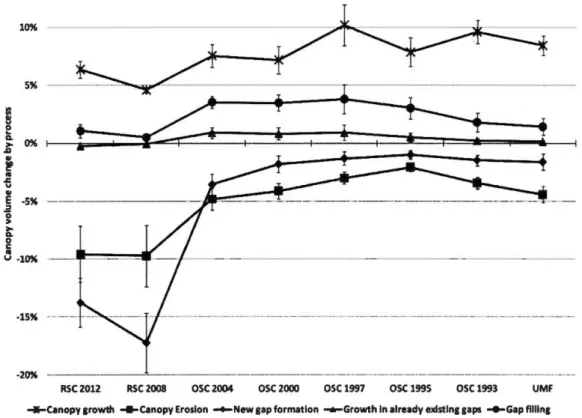
new gap formation, gap filling and growth in already existing gaps) with 95% intervals of variations around the mean due to GDHT variation between 1 and 10 meters. These are meant to illustrate variability only. Actual values can be found in the appendices. RSC refers to recent selection cuts-stands that were eut between 2007 and 2013. OSC refers to old selection cuts-stands that were eut the given year, before 2007. UMF refers to unmanaged forests ... 28 Figure 1.3 Mean of canopy volumes for each canopy process with the 95%
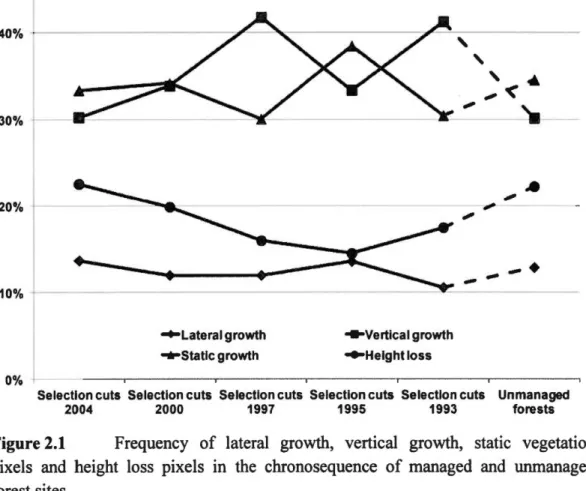
confidence intervals of variations due to gap detection height threshold from 1 to 10 m. Actual values can be found in the appendices. OSC refers to old selection cuts-stands that were eut the given year, before 2007. UMF refers to unmanaged forests-forests that have never been eut. ... 3 0 Figure 2.1 Frequency of lateral growth, vertical· growth, static vegetation pixels and
height loss pixels in the chronosequence of managed and unmanaged forest sites ... ... ... ... .... ... .. ... ... 50 Figure 2.2 Canopy height models of a portion of an unmanaged forest in A) 2007
and B) 2013. C) Canopy height difference between 2007 and 2013 (grayscale gradient of height loss in black to height growth in white) and D)
classification maps of the height changes as height loss ( dark grey), static growth (medium gray), vertical growth (light gray) and lateral growth (white). The pixels have a 50 cm resolution and the represented area is 30 x 30 meters ... 53
Figure 2.3 Canopy height models of a portion of a managed forest stand eut in 2004 in A) 2007 and B) 2013. C) Canopy height difference between 2007 and 2013 (grayscale gradient of height loss in black to height growth in white) and D) classification maps of the height changes as height loss (dark grey), static growth (medium gray), vertical growth (light gray) and lateral growth (white). The pixels have a 50 cm resolution and the represented area is 30 x 30 meters ... ... 54 Figure 2.4 Change in relative frequency ofheight loss (dark grey), static vegetation
(medium grey), vertical growth (light grey) and lateral growth (white) from 2007 (left side of central axis) to 2013 (right side of central axis) ... 56 Figure 2.5 Smoothing functions of vertical growth rate in cm per year as a function
of initial height (2007), fitted to vertical growth and static vegetation with height increase between 2007 and 2013 ... 57 Figure 2.6 Smoothing functions of vertical height growth rate in cm per year as a
function of initial height (2007) and maximum height difference with the irnmediate neighbors, fitted to vertical growth and static vegetation pixels with positive height change values. Smoothing functions have been restricted to the actual spread of the data ... 58 Figure 3 .1 Example of field-measured leaf coverage by height of an unmanaged
forest in a) 2007 and b) 2013. The numbers represent the estimated cover percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automatically. The superimposed points represent Lidar points for the same space ... 82 Figure 3 .2 Example of field-measured leaf coverage by height of a site eut in 1993
in a) 2007 and b) 2013. The numbers represent the estimated cover
percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automatically. The superimposed points represent Lidar points for the same space ... 83 Figure C.1 Field-measured leaf coverage by height of an unmanaged site in 2007.
The numbers represent the estimated cover percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automaticall y ... 121 Figure C.2 Field-measured leaf coverage by height of an unmanaged site in 2013.
The numbers represent the estimated cover percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automatically. The superimposed points represent Lidar points for the same space. The numbers at the top represent the number of vegetation layers in the canopy ... 122
Figure C.3 Field-measured leaf coverage by height of a site eut in 1993 in 2007. The numbers represent the estimated cover percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automatically. The superimposed points represent Lidar points for the same space ... ... 123 Figure C.4 Field-measured leaf coverage by height of a site eut in 1993 in 2013.
The numbers represent the estimated cover percentage of leaf material at a given height within the 5x5 m plots in 2014. The shading represents breaks assigned automatically. The superimposed points represent Lidar points for the same space. The numbers at the top represent the number of vegetation layers in the canopy ... 124
Tableau Table 1.1
Page LiDAR acquisition parameters ... 21 Table 1.2 Mean canopy height (± 1 standard deviation), area and frequency of
height reduction for the investigated study sites in 2007 and 2013. The frequency of negative canopy height change is the proportion of all the pixels that experienced a decrease in height of 1 meter or more between 2007 and 2013. RSC refers to recent selection cuts-stands that were eut between 2007 and 2013. OSC means old selection cuts-stands that were eut the given year, be fore 2007. UMF stands for unmanaged forests-forests that have never been eut ... 25 Table 1.3 The gap extinction rate, per site, according to the gap detection height
threshold. See Figure 1.1 for table headers definitions ... .. ... 31 Table 2.1 LiDAR acquisition parameters ... 44 Table 2.2 Mean density and standard deviation of first-returns LiDAR points per
pixels in the 2007 and 2013 canopy height models ... 49 Table 3.1
Table 3.2
LiDAR acquisition parameters ... 72 Model formulas for the MECHANISMS model and the STRUCTURES model. Every variables are binary parameters of presence or absence except for Area, which is continuous. GAP indicates new canopy gaps pixels within the height reduction areas, Standing indicate standing dead trees, Bent indicate bent trees, Broken indicate broken bole, Branch indicate large broken bran ch and Adj acency indicates presence of older gaps at the edge of the height reduction areas ... 77 Table 3.3 Model selection results for the mechanisms and structure models ... 78 Table 3.4 Contingency tables of presence or absence of gaps, adjacency to older
gaps and tree damage mechanisms within HRA. HRA can have more than one tree damage mechanism ... 79
Table 3.5 Percent of sites with 1, 2 or 3 canopy layers occupied by leafy material as determined visually with the 2013 Lidar data and leaf cover field data. This does not include the ground layer. The numbers in brackets are the total percentage of single layered canopy structures ... 80 Table A.1 Percentages of the main canopy, canopy erosion, new gaps formation,
existing gaps and filled gaps for unmanaged and partially-logged sites for gap detection height thresholds 1 to 10 m ... 113 Table B.1 Rate and shape values of the canopy extension gamma distribution.
The slope of the logistic regression is also given. All parameters are
Cette thèse visait à mieux comprendre les processus de renouvellement des forêts feuillues tempérées aménagées et non-aménagées en abordant de façon critique le cadre conceptuel de la dynamique des trouées. Au début du projet, les connaissances sur la dynamique des trouées de la forêt feuillue tempérée était limitées par certaines incertitudes liées à des failles dans le cadre conceptuel de la dynamique des trouées. Certaines sources de ces incertitudes ont été investiguées dans cette étude, à savoir les effets du choix de définition opérationnelle de trouées sur la mesure des propriétés des régimes de trouées, l'effet de la croissance latérale et verticale sur la structure de hauteur de la canopée et l'effet ( ou l'absence d' effet) des mécanismes de mortalité des arbres sur la création de trouées. Ces sources d'incertitude ont été analysées pour des forêts anciennes et des forêts amémagées par coupe de jardinage
Ce travail a été rendu possible grâce à l' acquisition de données Lidar ( «Light detection and ranging») multi-temporelle. Cela a permis de mesurer la hauteur de la canopée dans un intervalle de 6 ans, de 2007 à 2013, et ainsi de mesurer les changements de hauteur de la canopée. Dans le premier chapitre, nous nous sommes intéressés la dynamique de la canopée des forêts aménagées et non-aménagées. Nous avons ainsi trouvé un processus qui avait été peu étudié, l'érosion de la canopée. Ce phénomène se produit lorsqu'un arbre est endommagé ou meurt mais que cela ne crée pas de trouées. Les autres processus étaient liés à la croissance dans la canopée ou à la création et à la fermeture de trouées dans la canopée. L'érosion de la canopée était plus fréquente que les trouées dans les sites aménagés et non-aménagés, à l'exception des sites coupés durant l' étude. Nous avons aussi étudié l' effet de changer le seuil de hauteur utilisé pour détecter les trouées sur la fréquence des cinq processus. Nous avons montré que la perception des caractéristiques du régime de trouées est sensible au seuil de hauteur de détection des trouées. La structure de la canopée devrait prendre au moins 30 ans pour se rétablir suite à la coupe de jardinage.
Dans le deuxième chapitre, j ' ai appliqué un modèle stochastique Bayésien afin d'étudier l' effet de la croissance latérale et de la croissance verticale sur la structure de la canopée des forêts aménagées et non-aménagées. Nous avons ainsi montré que la fréquence de croissance latérale était assez similaire entre les forêts aménagées et
non-aménagées durant la période d'étude. Les résultats démontrent que la récupération de structure des forêts aménagées après les coupes est le fruit d'une diminution de la mortalité et d'une accélération de la croissance verticale des arbres de moins de 10 m de haut, comparativement aux forêts non-aménagées. Aussi, les résultats indiquent que les arbres dominants ont une croissance verticale lente et que la coupe de jardinage ne semble pas diminuer la fréquence des arbres à croissance verticale lente.
Dans le troisième chapitre, nous nous sommes penchés sur les causes de la faible fréquence de trouées lors de la mort ou de dégât aux arbres. Avant cette thèse, il était attendu que la mort d'arbres crée les trouées en forêt feuillue tempérée. Nous avons cependant montré au premier chapitre que l'érosion de la canopée était un phénomène très fréquent dans les sites étudiés. Nous avons donc testé deux hypothèses pouvant expliquer, ensemble ou séparement, la présence ou l'absence de trouées lors de morts ou de dégâts aux arbres; 1) soit que les trouées étaient dues aux mécanismes de <légats des arbres (i.e. branches cassées, tronc cassé, tronc plié, arbre mort sur pied, arbre déraciné); 2) soit que l'organisation spatiale de la structure verticale de la canopée explique ou non l'absence de trouées. Pour réaliser cela, 178 zones de perte de hauteur détectées à l'aide de Lidar multi-temporel ont été visitées à l' été 2014 pour déterminer les mécanismes de dégât des arbres présents dans la zone perturbée. Des mesures de structure de la canopée ont été prises dans des forêts aménagées et non -aménagées qui ont par la suite été combinées avec les données Lidar afin de déterminer la fréquence des structures de canopée pouvant donner lieu à des trouées en cas de perturbation de la canopée. Les résultats montrent que la structure préexistante de la canopée est une bien meilleure explication de la présence ou l'absence de trouées que les mécanismes de dégât des arbres. De plus, la distribution de fréquence de structure de canopée pouvant donner lieu à des trouées permet d' expliquer la fréquence de trouées dans les zones de perte de hauteur. Les trouées ne sont donc pas créées par la mort d'arbres; elle sont révélées par les perturbations dans la canopée.
La synthèse des résultats indique que le cadre conceptuel de la dynamique des trouées ne décrit pas complètement les processus de renouvellement de la forêt feuillue tempérée.
Mots-clés : Lidar multi-temporel;Trouées;Érosion de la canopée;Croissance verticale; Croissance latérale.
0.1 Définition des trouées
La dynamique de la forêt feuillue tempérée est caractérisée par des patrons spatio-temporels d'interrelations entre la canopée et la sous-canopée (Foré et al. , 1997). Dans ces forêts, la majeure partie des changements dans le temps sont causés par les trouées de petite taille dans la canopée. C'est pour cette raison que les trouées sont étudiées depuis longtemps afin de comparer les écosystèmes forestiers partout dans le monde (McCarthy, 2001) et que cela demeure pertinent de nos jours.
Les trouées ont été définies comme des zones de hauteur faible dans les canopées causés par la mort partielle ou complète d'un ou plusieurs arbres (Pickett et White, 1985 ; Runkle, 1981 ; Runkle, 1985). Cependant, cette définition est seulement un guide et son interprétation peut donner des résultats variables (Brokaw, 1982). La majeure partie des chercheurs adoptent ce guide comme définition conceptuelle et choisissent par la suite un seuil de hauteur pour distinguer les trouées de la canopée. Même si le choix de seuil de hauteur est habituellement fait sur la base de connaissances d'experts à l'aide d'observations de terrain (Runkle, 1992), leur validité n'est pas toujours claire (Lieberman et al., 1989). Au début de ce projet, peu d'études avaient examiné les effets de choisir différents seuils de hauteur pour détecter les trouées malgré qu'il ait été démontré dans le passé que les paramètres de définition de troués peuvent avoir un impact sur la perception des caractéristiques des régimes de trouées (Barden, 1989 ; Brokaw, 1982). La pertinence des seuils de hauteur utilisés pour détecter les trouées afin d'expliquer les processus écologiques des forêts est douteuse, tout comme la répétabilité des études qui les utilisent. Il y encore peu de connaissances sur les implications de choix de seuils de hauteur
spécifique. L'adoption d'une définition objective des trouées dans la canopée est une tâche difficile en partie à cause de difficultés lors de l'échantillonnage sur le terrain, des caractéristiques spécifiques des sites et des objectifs de recherche (Runkle, 1992). Les trouées ont été définies en prenant pour acquis qu'elles pouvaient être clairement délinéées dans la canopée, ce qui n'est pas toujours pertinent (Blackburn et al., 2014). L'application du cadre conceptuel de la dynamique de trouées signifie que la complexité de la forêt est réduite à une dichotomie trouée vs non-trouées peu importe son hétérogénéité (Lieberman et al., 1989). Malgré ces bémols, le cadre conceptuel de la dynamique des trouées pourrait ne pas être une mauvaise abstraction pour l'étude de la forêt feuillue tempérée. La relation entre les trouées et la mort d'arbres est un phénomène indéniable et de nombreuses recherches ont montré son importance pour les processus de renouvellement de la forêt ( e.g. Abe et al., 1995 ; Barden, 1981 ; Brokaw, 1985; Runkle, 1985 ; Watt, 1947; Williamson, 1975; Yamamoto, 1992). Le problème avec le concept de trouées vient du fait que les trouées ne sont pas des objets discrets, mais plutôt une absence d'arbres. Une trouée est seulement un objet abstrait qui aide dans la conceptualisation de la structure de la canopée.
Les tailles minimales et maximales des trouées incluses dans une recherche sont des paramètres qui peuvent influencer la prise de données. La taill~ minimale de trouées est habituellement définie par les limitations de l'échantillonnage terrain puisque les petites trouées dans une canopée haute sont difficiles à mesurer précisément. De plus, la taille minimale de trouées basées sur une réponse fonctionnelle est difficile à déterminer et peut changer d'un endroit à l'autre (Brokaw, 1982 ; Kern et al., 2016). Cependant, les petites trouées sont souvent le type de trouées le plus commun (Barden, 1989). De plus, il y a désormais des méthodes pour déterminer les classes de taille de trouées en fonction de critères précis, tels que la distribution de lumière incidente (Zhu et al., 2015) ou la taille des couronnes des arbres dominants (Schneider et Larson, 2017). Écarter des trouées plus petites que certaines valeurs
modifie la distribution de taille des trouées et influence le calcul du temps de renouvellement des peuplements forestiers (Brokaw, 1982). Il est généralement accepté que la taille maximale d'une trouée devrait être de 200 m2 (McCarthy, 2001) à 1000 m2 (Muscolo et al., 2014). La dynamique dans les trouées plus grandes serait caractérisée par des processus de renouvellement de peuplements.
0 .2 Dynamique des trouées
Le cadre théorique décrivant le régime de perturbations naturelles en forêt feuillue tempérée, la dynamique des trouées, est caractérisé par la création continuelle d'ouvertures généralement à l'échelle d'une couronne d'arbre (Runkle, 1985). La dynamique des trouées est responsable de la structure des forêts anciennes (Franklin
et al., 2002). La dynamique des trouées a une influence importante sur le
fonctionnement et la composition des vieux peuplements, mais joue aussi un rôle structurant dans les forêts plus jeunes (Hart et Grissino-Mayer, 2009) et dans les forêts aménagées (Angers et al. , 2005). Les mécanismes de mort d'arbres peuvent
avoir des incidences et des fréquences différentes en fonction de la source des perturbations et de leur intensité (Seymour et al., 2002). Les régimes de perturbations
peuvent être caractérisés de plusieurs façons (Laska, 2001). Les perturbations exogènes à grande échelle, tels que les chablis, les épidémies d'insectes ou les tempêtes de verglas peuvent frapper soudainement de grandes zones. De tels événements, quoique habituellement rares, peuvent déraciner les arbres, casser les troncs et les branches et créer de plus grandes trouées que les perturbations endogènes (White, 1979) et ont ainsi des effets plus importants sur la structure des forêts (Lorimer, 1989). D'un autre côté, les perturbations endogènes n'affectent habituellement qu'un ou quelques arbres à la fois et peuvent résulter dans une mort d'arbres plus graduelle. La mort d' arbres graduelle est un phénomène tres fréquent dans certaines forêts tempérées nordiques (Krasny et Whitmore, 1992). Les morts d'arbres soudaines et graduelles peuvent avoir des répercussions différentes sur la dynamique des communautés forestières (Pickett et White, 1985).
Les mécanismes de dégât des arbres sont variés en forêt feuillue tempérée. Le déracinement est le mécanisme de dégât des arbres le plus violent dans les forêts feuillues tempérées (Schaetzl et al. , 1988), et celui qui a le plus d'effets sur leur composition (Nakashizuka, 1989). Les déracinements partiel ou total peuvent détruire la régénération avancée, bien que cela n'arrive pas systématiquement (McGee et al. , 2007). Contrairement aux autres mécanismes de dégât des arbres, le déracinement implique que le sol est perturbé (Schaetzl et al. , 1989). Les faiseurs de trouées dont le tronc casse peuvent tomber dans les trouées et écrasent parfois une partie de la régénération avancée. Ce mécanisme serait le plus fréquent en forêt feuillue tempérée (Barden, 1981). Les branches périphériques des couronnes d'arbres peuvent subir du dépérissement terminal par érosion mécanique (Hajek et al. , 2015 ; Hossain et Caspersen, 2012). Ce mécanisme serait même plus important comme interaction de couronne que la recherche de lumière (Hajek et al. , 2015). Un autre type de dégât peu mentionné dans la littérature est lorsque le tronc des arbres plie et reste plié (Everham et Brokaw, 1996). Ce phénomène est caractérisé par un système racinaire intact, la partie inférieure du tronc qui reste verticale alors que la partie supérieure du tronc est pliée (Gresham et al. , 1991 ). Les arbres meurent fréquemment sur pied (Busing, 2005). Après un certain temps, les chicots tombent au sol (Busing, 2005) et peuvent donc être confondus dans certaines situations avec des arbres cassés. Il est attendu que la mort sur pied crée des petites trouées (Yamamoto, 1992), bien que l'effet de la chute éventuelle de la tige sur les propriétés des trouées n'ont pas été investiguées. Les arbres au sol finissent par se décomposer et contribuent à la dynamique forestière (Franklin et al., 1987 ; Runkle, 2000).
Une des plus grandes limitations dans l'étude des arbres et des forêts est que les phénomènes qui les caractérisent se produisent généralement sur une longue période. En effet, les arbres peuvent vivre plusieurs centaines d'années et la dynamique des peuplements peut être affectée par des évènements s'étant produits sur d' encore plus grandes périodes (McCarthy, 2001 ). Le temps nécessaire pour étudier en détail la
dynamique forestière est trop long pour être facilement réalisable. Pour contrer ce problème, plusieurs stratégies sont utilisées couramment comme alternative à des études à long terme. Les plus courantes sont l'utilisation de modèles et la substitution espace-pour-temps. Les modèles sont des représentations mathématiques de la réalité qui sont utilisées pour simuler des scénarios et ainsi tirer des connaissances qui peuvent ensuite être transposées à des situations réelles, si les modèles sont bien formulés et analysés (Burnham et al., 2011 ; Johnson et Omland, 2004). La substitution espace-pour-temps est une forme d'analyse permettant d'inférer des tendances temporelles par l'étude de sites d'âge différents arrangés dans une chronoséquence (Pickett, 1989). Ce type d'analyse est particulièrement utile lorsqu'il n'y a pas encore assez de données accumulée sur un phénomène (Landres et al., 1999). Cette forme d'analyse est utilisée dans les chapitres 1 et 2 de cette thèse. 0.3 Autres types d'ouvertures de la canopée
Les recherches sur les trouées ont généralement portées sur celles créées par la mort d'au moins une partie d'arbre (e.g. branche). Cependant, les ouvertures dans la canopée ne sont pas toutes des trouées. Les chercheurs choisissent parfois d'étudier des ouvertures de la canopée dues à d'autres causes. Sur cette base, deux types d'ouvertures de la canopée peuvent être distinguées : les trouées développementales (appelé trouées dans le reste du document) et les ouvertures sans régénération (Senecal, 2011 ). La principale distinction entre les trouées et les ouvertures sans régénération tient à leur origine. Les trouées sont des ouvertures dans la canopée créées par la mort partielle ou complète d'arbres. Les ouvertures sans régénération sont des ouvertures dans la canopée créées ou maintenues par des causes autres que la mort d'arbres. Elles ont parfois été appelées trouées édaphiques lorsqu' elles étaient causées par des conditions édaphiques ou topographique (Bartemucci et al., 2002 ; Lertzman et al., 1996). Elles ont aussi parfois été appelées trouées de rebord lorsqu'elles sont causées par des sols peu profond et du roc dénudé (Kimball et al., 1995). Même si elles n' ont pas été étudiées comme des ouvertures sans régénération,
les ouvertures maintenues par la présence de végétation récalcitrante (Royo et Carson, 2006) peuvent aussi être vues comme telles puisqu' elles perturbent parfois la dynamique de trouées (Koukoulas et Blackburn, 2004). Les ouvertures sans régénération sont habituellement ignorées dans les études sur la dynamique des trouées puisqu'elles ne sont pas considérées comme des sites potentiels de régénération de la forêt. Les ouvertures sans régénération ne contribuent pas directement à la dynamique des trouées (Lertzman et al. , 1996) mais peuvent être confondues avec des trouées, particulièrement par l'usage de la télédétection (Senecal, 2011). Un autre facteur confondant est que des trouées peuvent s' ouvrir en bordure des ouvertures sans régénération. La distinction de ces deux types d'ouvertures devient alors difficile. Les petites ouvertures sans régénération peuvent aussi se refermer par comblement latéral. Les trouées et les ouvertures sans régénération peuvent ainsi être confondues. Il est donc probable que la fréquence des ouvertures sans régénération est sous-estimée et que celle des trouées comme site de régénération de la forêt est elle surestimée.
Un autre phénomène qui n'a été que peu étudié est la mort d'arbres sans création de trouées. La littérature mentionne ce phénomène parfois, bien que la fréquence de ce phénomène ne soit que rarement quantifiée (mais voir Vepakomma et al. 2010). Pourtant, la mort · d'arbres sans création de trouées a été identifiée comme un problème potentiel pour l'estimation des propriétés de la dynamique des trouées (Barden, 1989 ; Brokaw, 1982). Pour qu' il y ait de la mort d'arbres sans création de trouées, il est nécessaire qu'il y ait de la végétation préétablie sous la canopée et qµe cette végétation ne soit pas endommagée par le mécanisme de mort d'arbres. Les renseignements nécessaires à l'évaluation de l'importance de ce phénomène dans les études sur les trouées sont habituellement absents. Cela fait appel à des notions de structure puisque les forêts tempérées sont habituellement caractérisées par une structure verticale complexe (Messier et al. , 2009). Cela nécessiterait une description détaillée des éléments structuraux des forêts. Malgré son importance, la structure de
la canopée est rarement décrite en détail dans les études sur les trouées. L'utilisation de mesures de structure verticale révèle autant la possibilité de retrouver des trouées sous la canopée que de constater une absence de trouées dans tous les étages la canopée (Connell et al., 1997 ; Parker, 1995). Cela nous amène à formuler deux hypothèses pour expliquer la création de trouées lors de la mort d'arbres. La première hypothèse est que les trouées sont créées par la mort d'arbres et que certains mécanismes de dégât des arbres favorisent ou nuisent aux arbres déjà établis sous la canopée. La deuxième hypothèse est que la structure verticale des peuplements forestiers est le facteur déterminant de la présence ou non de trouées lorsqu'il y a du dégât aux arbres de la canopée. Les trouées ne seraient alors pas créées par la mort d'arbres, elles seraient plutôt révélées. Ces hypothèses sont testées dans le 3e chapitre de cette thèse afin de déterminer la plus vraisemblable.
0 .4 Effets des trouées sur la croissance des arbres
Les trouées changent la disponibilité en lumière (Canham et al. , 1990 ; Gendreau-Berthiaume et Kneeshaw, 2009) et génèrent des conditions hétérogènes de structure de la canopée, ce qui fournit des opportunités de croissance pour un large éventail d'espèces. La variabilité des caractéristiques des peuplements forestiers fait que la croissance ne répond pas toujours (Bannon et al. , 2015 ; Beckage et Clark, 2003). Plusieurs études ont montré une plus forte croissance de la végétation dans les trouées que sous couvert fermé (Canham, 1985 ; Canham et al. , 1990 ; McClure et al. , 2000 ; Runkle, 1981) et aussi suite à la coupe de jardinage (Beaudet et Messier, 2002 ; Beaudet et al. , 2004).
En réponse à l'ouverture des trouées, il se produit de la croissance verticale et de la croissance latérale qui mène éventuellement à la fermeture des trouées. La croissance latérale est la croissance des branches dans l'espace de la trouée. Cette vitesse peut varier dépendamment de plusieurs facteurs liés à l'identité des arbres, tels que leur espèce, leur âge, leur capacité à utiliser la lumière, etc. Des vitesses de croissance
latérale de 18 cm/an ont été rapportées (Runkle et Yetter, 1987), bien qu'il ait été montré que cette vitesse ralentissait avec l'augmentation de l'âge des trouées (Runkle, 1998). La croissance latérale peut mener au développement de couronnes asymétriques qui ont alors plus de chances de tomber vers les trouées (Brisson, 2001 ; Young et Hubbell, 1991). Le second mécanisme de fermeture de trouées est la croissance verticale des arbres jusqu'à ce qu'ils atteignent la canopée. Les semis et les gaules ne peuvent habituellement pas croître suffisamment pour atteindre la canopée avant qu'elle se referme par croissance latérale, bien que ce soit toutefois possible dans certaines situations (Cole et Lorimer, 2005). En général, la canopée doit s'ouvrir et se refermer plusieurs fois au-dessus d'un arbre avant qu'il soit suffisamment grand pour occuper la canopée (Canham, 1985, 1990). Cela signifie qu'il y a souvent de la régénération avancée qui s'est établie avant la perturbation (McClure et al., 2000). Les types de dégâts des faiseurs de trouées devraient donc avoir un impact sur les conditions initiales dans les trouées, ce qui devrait aussi influencer la vitesse de fermeture de la canopée.
Il est attendu que . certains arbres augmentent leur vitesse de croissance verticale lorsqu'ils détectent le risque de se faire dépasser par un compétiteur proche (Schmitt et Wulff, 1993), mais seulement dans une situation où les niveaux de lumière le permettent (Lei et Lechowicz, 1998). Les arbres qui ne sont pas en compétition pour la lumière pourraient investir des ressources dans d'autres compartiments (Givnish, 1988), alors que ceux qui sont dans l'ombre d'individus plus grands pourraient avoir une croissance verticale plus faible en raison de niveaux de lumière bas (Beaudet et Messier, 1998). De plus, la vitesse de croissance verticale change avec la hauteur des arbres (Cole et Lorimer, 2005 ; Yetter et Runkle, 1986). Les processus de croissance de la canopée dépendent de la structure de hauteur des forêts puisque les arbres poussent dans les ressources en lumière tout en tentant d'éviter la compétition (Brisson, 2001 ).
La majeure partie des petites trouées devraient se refermer par croissance latérale alors que les grandes trouées devraient rester ouvertes plus longtemps (McCarthy, 2001 ). Cependant, les processus de croissance ne sont pas purement discrets. Par exemple, à haute latitude, la végétation sous couvert de la canopée peut recevoir de la lumière de plusieurs trouées dépendamment de la position du soleil (Ban et al., 1998). De plus, les trouées peuvent s'agrandir lors de la mort d'arbres à leur bordure (Vepakomma et al., 2012). Les variations de structure de hauteur interne des trouées sont souvent le résultat d'expansion des bordures, bien que cette caractéristique n'est habituellement pas étudiée (Runkle, 1992). Les jeunes arbres de plusieurs espèces, mais pas toutes, vont croitre vers la lumière (Schamp et al., 2007). Cependant, la réaction des plantes à l'intensité et à la qualité de la lumière sont encore peu connues (Vandenbussche et al., 2005).
0.5 Forêts aménagées
La coupe de jardinage, un type de coupe partielle qui vise à préserver une structure d'âge inéquienne des peuplements, est considérée comme étant la forme d'aménagement de la forêt qui produit une structure inéquienne de d'arbres dotée d'une distribution de diamètres qui est similaire à celle trouvée en forêts feuillues tempérées (Majcen, 1994). Cependant, cette comparaison a été faite en assumant 1) que le régime de perturbations naturelles est décrit convenablement par la dynamique des trouées et 2) que l'application de la coupe de jardinage permettrait de maintenir la structure et la composition des peuplements, ce qui n'a pas été suffisamment testé. Ainsi, des études ont montré que la structure et dynamique du bois mort des forêts aménagées par coupes de jardinage étaient différentes de celle des forêts non-aménagées (Angers et al., 2005 ; Doyon et al., 2005). Outre ces différences, il a été montré que les résultats des coupes de jardinage ont été en deçà des performances escomptées, comparativement aux résultats obtenus avec les dispositifs expérimentaux originellement mis en place (Bédard et Majcen, 2003). Cela s'expliquerait par une sélection des arbres non conformes aux principes de la coupe
de jardinage (Majcen et al., 2005). Toutefois, il a été estimé que le rétablissement du
stock en bois des forêts jardinées serait envisageable dans un horizon d'un peu plus de 20 ans (Bédard et Brassard, 2002 ; Majcen et al., 2005). La coupe de jardinage
devrait donc s' accompagner d'une diminution de fréquence des arbres à faible croissance. Cette caractéristique n'a été que peu étudiée jusqu' à maintenant. Nous ne savons pas non plus comment la croissance verticale des arbres se compare entre les forêts aménagées et non-aménagées. Il a été noté que la croissance des arbres de faible diamètres était celle qui a le plus bénéficié des nouvelles conditions de lumières suite à la coupe de jardinage et que si cette croissance est maintenue la surface terrière des sites jardinés pourrait revenir a ses niveaux pré-coupes en 20 ans (Bédard et Majcen, 2003). Un rétablissement de la canopée dans un horizon de temps similaire est donc envisageable. Si ce n'est pas le cas, chaque rotation successive entrainerait une dégradation de la canopée. Dans ce cas, la coupe de jardinage ne serait vraisemblablement pas soutenable à long terme.
0.6 Lidar
J'ai utilisé les données de télédétection Lidar aéroporté comme données principales pour informer les analyses des trois chapitres de cette thèse. Un capteur Lidar aéroporté est un senseur actif généralement monté sur un avion. Le principe du Lidar est de mesurer la distance entre le senseur et les cibles au sol à l' aide d'un laser. D'autres senseurs combinés au Lidar permettent de déterminer la position et l'orientation du Lidar au moment de l'émission du laser et de son retour au senseur. Ces paramètres permettent de calculer précisement la position de la cible. De plus, les Lidars modernes peuvent généralement obtenir des mesures sous la canopée. Ces caractéristiques permettent donc au Lidar de mesurer la hauteur de la canopée des forêts et d'obtenir des renseignements sur la topographie du sol. La résolution des données Lidar dépend principalement de la densité des points de mesures effectuées lors des vols d'acquisition et des caractéristiques du senseur Lidar. Avec les capteurs récents, la densité de points est tellement grande qu'il est généralement plus facile de
transformer les renseignements obtenus par le Lidar en modèles de hauteur sous forme d'images. Pour cela, il est nécessaire de procéder à l'interpolation des images à partir des données Lidar. Cela permet de développer des modèles de hauteur réalistes et fiables sans données manquantes (Vepakomma et al., 2008). Cette façon de procéder a de nombreux avantages en terme de traitement des données ( e.g. facilité, rapidité, nécessite moins de mémoire). Peu importe les données utilisées, l'utilisation d'ordinateurs implique le développement de modèles qui sont des réprésentations imparfaites de la réalité. Cette transformation en image fait que l'unité d' analyse devient le pixel et peut ainsi cacher la précision réelle sous-jacente de l' information ayant servi à construire le modèle de hauteur de canopée.
0. 7 0 bj ectifs de l'étude
Les contributions principales de cette thèse sont de remettre en question certains aspects du cadre conceptuel de la dynamique des trouées et d' offrir une meilleure description des processus de croissance et de perte de hauteur de la canopée dans les forêts feuillues tempérées non-aménagées et aménagées par coupe de jardinage. Pour cela, trois objectifs principaux ont été abordés dans trois chapitres. Le 1er chapitre constitue le point de départ. Dans le premier chapitre, la dynamique des trouées des forêts non-aménagées et jardinées ont été analysées et comparées en tenant compte des pertes de hauteur sans création de trouées. Les limites du concept de trouées comme outil de suivi des processus ont été testées en tenant compte de la sensibilité associée au choix de seuils de hauteur utilisé pour détecter les trouées. Les 2 e et 3 e chapitres ont permis d'approfondir certains aspects qui étaient toujours flous après le 1er chapitre. Dans le 2e chapitre, les processus de croissance latérale et verticale ont été analysés plus en détail dans une chronoséquence de sites jardinés issue d'une substitution espace-pour-temps et dans des sites non-aménagés. Le rétablissement de la canopée suite à la coupe de jardinage a été analysé en comparant avec des sites non-aménagés. Dans le 3e chapitre, j'ai tenté de déterminer si la création de trouées lors de dégâts aux arbres était due à certains mécanismes de dégâts ou à la structure
verticale de la canopée. Pour réaliser cela, des zones ayant perdu de la hauteur entre 2007 et 2013 ont été visitées en forêt afin de déterminer les mécanismes ayant causé ou non des trouées. La distribution verticale de matériel végétal le long de transects a aussi été mesurée afin d'établir une référence de classes de structure de forêt similaire à celle de Parker (2000). Finalement, le support pour deux hypothèses a été calculé dans un cadre d'inférence à hypothèses multiples afin de déterminer si les trouées sont créées par des mécanismes spécifiques de dégât des arbres ou si elles sont dues à la distribution verticale des arbres.
Pour réaliser ces objectifs, des peuplements forestiers non-aménagés et aménagés par jardinage situés en Outaouais ont été étudiés. Les forêts non-aménagées étaient situés dans la réserve écologique de la Forêt-la-Blanche (45°44'N, 75°16'0) et dans la réserve faunique Papineau-Labelle (45°59'N, 75°20'0). Les peuplements aménagés étaient tous situés dans la réserve faunique Papineau-Labelle. La réserve écologique de la Forêt-la-Blanche est une aire protégée de 2 052 ha vouée à la conservation et à la recherche. La réserve faunique Papineau-Labelle occupe une superficie de 162 800 ha vouée à la récréation et à la préservation de la faune. Les aménagements forestiers et les activités récréatives (e.g. chasse, pêche, camping) y sont permis sur une partie du territoire. Les parcelles étudiées ont été choisies en fonction des objectifs de recherche et de la disponibilité des données. La rareté des sites anciens et jardinés dans la zone pour laquelle les données de télédétection étaient disponibles a donc été un facteur limitant. Les sites étaient tous caractérisés par des peuplements dominés par l'érable à sucre. Les sites jardinés que nous avons identifiés ont été aménagés en 1993, 1995, 1997, 2000, 2004, 2008 ou 2012. Le site jardiné en 2012 avait déjà subi une coupe à diamètre limite en 1985. L'historique passé des autres sites est inconnu. Il est cependant probable qu'ils ont déjà subi des coupes d'écrémage dans le passé (Angers et al. , 2005).
MANAGEMENT IMPLICATIONS OF VARYING GAP DETECTION HEIGHT THRESHOLDS AND OTHER CANOPY DYNAMICS PROCESSES IN
TEMPERATE DECIDUOUS FORESTS
This article was published in February 2018 in Forest ecology and management, volume 410
Jean-Francois Senecal, first author
Centre d'étude de la forêt, Département des sciences biologiques, Université du Québec à Montréal, C.P. 8888, Suce. Centre-Ville, Montréal, Canada, H3C 3P8. Senecal.Jean-francois.3@courrier.uqam.ca
Frédérik Doyon
Institut des Sciences de la Forêt Tempérée, Université du Québec en Outaouais, 58 rue Principale, Ripon, Canada, JOV 1 VO. Frederik.doyon@uqo.ca
Christian Messier
Centre d'étude de la forêt, Département des sciences biologiques, Université du Québec à Montréal, C.P. 8888, Suce. Centre-Ville, Montréal, Canada, H3C 3P8.; Institut des Sciences de la Forêt Tempérée, Université du Québec en Outaouais, 58 rue Principale, Ripon, Canada, JOV 1 VO. Christian.messier@uqo.ca
biomes around the world. Under the classical gap dynamics model, only canopy gaps are considered as a dynamic zone. lt is also assumed that gapless canopy change is either uncommon or unimportant for forest dynamics, even though it has rarely been quantified. In the temperate deciduous forest biome of Quebec, Canada, forest management uses selection cutting to emulate gap dynamics, but little is known about canopy dynamics in unmanaged and managed forests. Here, we investigated five canopy processes: three generating positive height change ( canopy growth, gap closure and gap filling) and two generating negative height change (newgap creation and canopy erosion) using multi-temporal LiDAR data in unmanaged and managed (by partial cutting) forests at different periods during the last 24 years. Canopy erosion, which is canopy height reduction without gap formation, was very common in all studied forests, whatever the gap height thresholds used. This canopy process was often more common than new canopy gaps. Total canopy height reduction due to canopy erosion was also comparable to that of new canopy gaps. Furthermore, we show that varying the height threshold used to detect gaps changes the perception of canopy gap dynamics. Canopy gap closure rates suggest that canopy gaps will remain more frequent in managed forests 30 years after cuts compared to unmanaged forests. At the same time, canopy cover of managed forests became more continuous than that of unmanaged forests. Our results show that it will take at least 30 years for natural canopy dynamics to recover following cuts at the currently used harvest intensities. We also show that measurements of canopy dynamics processes are sensitive to canopy gap detection height thresholds. This sensitivity is clearly limiting the reproducibility and the comparability of studies in gap ecology.
they provide environmental conditions that favour establishment and growth of many tree species (Canham, 1988b; Kem et al., 2012) due to increased light and below-ground resources (Beckage et Clark, 2003 ; Caspersen et Saprunoff, 2005). Although canopy gaps have been studied for decades (Jones, 194 5 ; Runkle, 2012 ; Watt, 194 7), the concept has always been somewhat unclear since gaps do not exist exactly as physical objects; canopy gaps are abstract constructs that are used to study canopy structure or canopy structure change, and multiple methods have been designed to describe what a gap is.
The classical conceptual definition that has been most commonly used is probably Brokaw' s «treefall gap» that describes a gap as the projection of a hole in the forest canopy extending through all levels to a height threshold of two meters above ground (Brokaw 1982). Other definitions have been proposed over the years, such as the "expanded gap" (Runkle 1982). In the majority of previous gap studies, the gap detection method and its threshold parameters were determined based on research objectives and expert knowledge of the forest dynamics of the study sites. However, the choice of specific height threshold parameters, like height of the vegetation in the canopy openings or minimum size of the opening, influences the results and limits repeatability of these studies (Barden 1989; Brokaw 1982; Lobo and Dalling 2014). These subjective choices have limited the ability to compare results across studies, particularly those relating to upscaled landscape patterns or processes, like the gap turnover rate (Després et al. 2017). Even repeated analyses of gap dynamics such as those using multi-temporal LiDAR data are not immune to these subjective choices, although they ensure that canopy openings result from a decrease in canopy height. Gap studies are based on a gap vs. non-gap dichotomy as part of the detection protocol, which reduces canopy structure complexity to two classes (Lieberman et al.
1989). However, as mentioned by Lieberman et al. (1989), "the forest canopy is not a swiss cheese". It has been pointed out that most methods/thresholds used may have led to the study of a limited set of forest canopy height reduction events (Nadkarni et al. 2008). It is, in fact, possible to observe tree death without gap creation (Brokaw 1982; Connell et al. 1997; Young and Hubbell 1991), and yet this pattern has not been carefully recognized and investigated as a canopy dynamics phenomenon. Most studies have focused on the role of gaps in forest dynamics where the forest is seen as a group of spatially localized stems under a canopy, instead of looking at it as a collection of leaves forming a 3D structure from above. To our knowledge, no study has quantified the occurrence of tree death without gap creation and its effect on canopy dynamics.
Understanding canopy dynamics and its associated effects on tree communities is particularly relevant with the increased use of the concept of emulating natural disturbance, even though it has been misused in some cases (Kuuluvainen 2009; Landres et al. 1999; Seymour et al. 2002). Emulating natural disturbance has been the comerstone of the ecosystem management paradigm (Gauthier 2009). Many forest ecologists have recommended the use of partial eut silvicultural systems, like selection cutting or irregular shelterwood, for mimicking the gap disturbance regime (Majcen 1994; Raymond et al. 2009; Seymour et al. 2002). However, it has been suggested that managing temperate deciduous forests with selection cutting might not be an adequate way to emulate the natural disturbance regime of this biome as the spatio-temporal patterns of trees and shrub structures in selection eut forests differ from those in unmanaged old-growth forests (Angers et al. 2005; Doyon et al. 2005), and because partial cuts often do not create the microsites necessary for seedling establishment (Bolton and D' Amato 2011). The objective of this paper was to describe and compare the canopy processes of managed (by partial-cutting) and unmanaged forests using multi-temporal airbome LiDAR data. We hypothesized that canopy dynamics will be different between managed and unmanaged forests. We
expected that the positive and negative growth processes would balance each other out in unmanaged forests while the growth processes would be positive for managed forests after partial cutting. We also expected less gap creation but greater gap closure in managed forests until a certain period after the eut when canopy processes start to become similar in both managed and unmanaged forests. Finally, we wanted to verify the sensitivity of observed patterns in canopy processes to the gap detection height threshold.
1. 3 Study area and Methods 1.3.1 Study area
The study area was located in the south of the province of Que bec, Canada. Nine sites subjected to selection cuts and one unmanaged site were located in the Papineau-Labelle wildlife reserve ( 45°59' N, 75°20' W). Two other unmanaged sites were located nearby in the ecological reserve of the Forêt-la-Blanche (45°44' N, 75°16' W). The partially eut stands had been subjected to selection cuts at different times over the last 25 years (1993 , 1995, 1998, 2000, 2004 and 2008). One additional site was subjected to diameter limit cuts, a type of partial eut to cull the bigger trees, in 1985, followed by a selection eut in 2012. The history of human-caused disturbances for the other managed sites is unknown. However, it is likely that there was some form of high grading performed there in the past (Angers et al. 2005). The study area used in the analysis is comprised of 827 ha and 352 ha of partially eut and unmanaged stands, respectively (Table 1.1). Climatic conditions are very similar in all sites. The average annual temperature in the study area varies from 2.5 to 5 °C, with a growing season (daily average over 5.6 °C) of 180-190 days (Robitaille et Saucier, 1998). Mean annual precipitation ranges from 900 to 1000 mm with a third falling as snow. The dominant tree species in the area are sugar maple (Acer saccharum Marsh.) often in combination with American beech (Pagus grandifolia Ehrh.). Other notable tree species are basswood (Tilia americana L.), Eastern hemlock (Tsuga canadensis (L.)
Carrière), ironwood (Ostrya virginiana (Mill.) K. Koch), red maple (Acer rubrum L.), striped maple (Acer pensylvanicum L.), white ash (Fraxinus americana L.), balsam fir (Abies balsamea), white elm (Ulmus americana L.) and yellow birch (Betula alleghaniensis Britt.). All study sites are mature uneven-aged forests. To the best of our knowledge, the unmanaged forests have not suffered noticeable stand-level catastrophic or anthropogenic disturbances in the last 200 years. However, the study area was affected by the North American ice storm of 1998, receiving between 40 and 100 mm of freezing rain, causing light to severe damage to forest stands in the study area (Chabot, 1998). Furthermore, beech bark disease (Houston, 1994) has been present in the study area for several years, causing mortality and defects on large beeches. These events might have led to a higher than normal level of advanced regeneration, although normality is highly subjective when speaking of mature or old-growth forests (Pesklevits et al., 2011).
1.3.2 LiDAR data processing
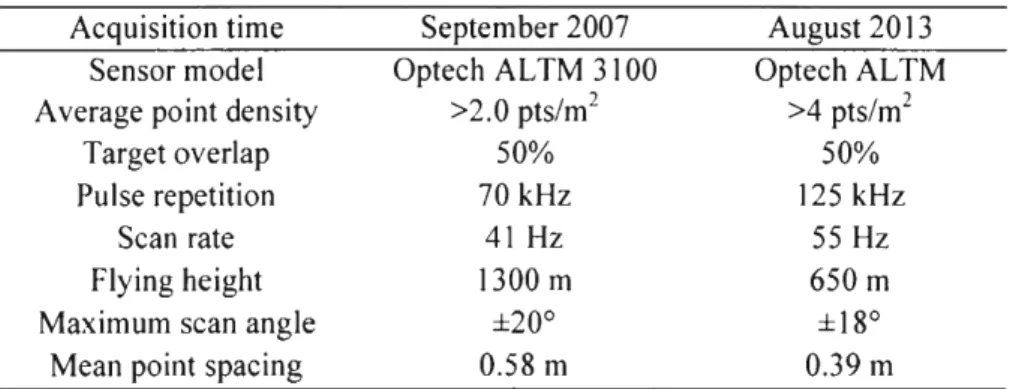
Two LiDAR datasets were used in this study. The first airbome discrete-retum LiDAR dataset was acquired in September 2007 using an Optech AL TM 3100 instrument operating at an average altitude of 1300 m (Table 1.1 ). Average point density was over two retums per m2, maximum half-scan angle was 20°, scan rate was 41 Hz, and line spacing was 750 m (for a targeted 50% overlap between strips). The second airbome discrete retum LiDAR dataset was acquired in August 2013 using an Optech ALTM Gemini instrument operating at an average altitude of 650 m. Average point density was over four retums per m2, maximum half-scan angle was
18°, scan rate was 55 Hz, and line spacing was 261 m (for a targeted 50% overlap between strips). Vepakomma et al. (2011) showed that two points per m2 were sufficient to identify canopy gaps. Preprocessing of the LiDAR data was accomplished with the Terrascan software package (Terrasolid, Leppavaara, Finland). This process included data cleaning, bird hit removal, and classification of LiDAR points as ground surface hits using the morphological properties of the point cloud.
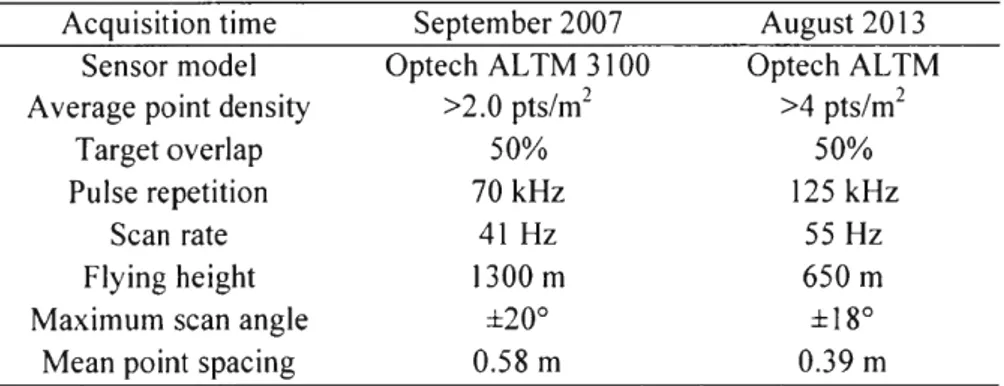
Table 1.1 LiDAR acquisition parameters Acquisition time
Sensor model Average point density
Target overlap Pulse repetition
Scan rate Flying height Maximum scan angle
Mean point spacing
September 2007 Optech ALTM 3100 >2.0 pts/m2 50% 70 kHz 41 Hz 1300 m ±20° 0.58 m August 2013 Optech ALTM >4 pts/m2 50% 125 kHz 55 Hz 650 m ± 18° 0.39 m
The canopy height models were generated following the guidelines of V epakomma et al. (2008). The multi-temporal LiDAR data was first co-registered to ensure that there were no significant systematic planimetric errors. This was assessed by producing height models with the first return Lidar points of 2007 and 2013. Height above sea level at locations known to be devoid of vegetation were compared in the two datasets to ensure that height shift was minimal. Furthermore, we also assessed horizontal shift, which is a problem caused by errors from the Lidar sensor that cause Lidar returns being registered in an incorrect location. On a height change model, this problem can be detected as several tree crowns appearing to have all lost height on a side having grown laterally on the opposite side. As there was no systematic shift of any kind detected, we proceeded with height models production. A digital terrain model was created at a 50 cm resolution from the combination of points classified as ground retums in the two LiDAR datasets. The lowest value was assigned to the pixels in which ground points fell. Pixels with missing values were interpolated by inverse distance weighting using the combined ground points and a power of two for the weighted average. The resolution of the. height models was determined based on average point spacing of the LiDAR datasets as suggested by Vepakomma et al. (2008). The digital surface models of 2007 and 2013, representing ground elevation above sea level, were produced using the same procedure, but with the highest height of LiDAR retums in each pixel instead of the lowest. The canopy height models
(eHM) of 2007 and 2013, high-resolution raster representations of the height of vegetation above the ground, were then obtained by subtracting the digital terrain model from the corresponding digital surface models. A height change image was produced by subtracting the 2007 eHM from the 2013 eHM. Positive values were interpreted as canopy height growth and negative values as canopy height loss.
The study areas were delimited within the 2x2 km square tiles to obtain relatively homogeneous and comparable characteristics, such as species composition, site history, topography and a minimum continuous area of 42 ha. The bodies of water were removed. We classified the sites into three management categories: the unmanaged forest sites (UMF), the sites partially eut between 1993 and 2004 as the old selection eut sites (OSC), and the sites partially eut in 2008 and 2012 as the recent selection eut sites (RSe). We analyzed ose separately from the RSe because cuts were performed in the latter sites between the 2007 and 2013 LiDAR data acquisitions. Thus, any height loss observed in the RSe is predominantly due to cuts, while those in the ose are not. Properties of these sites are presented in table 1.2, irrespective of the gap detection height threshold.
1.3.3 eanopy dynamics processes
We produced maps of the following five canopy dynamics processes: new gap formation, canopy erosion, filled gap, existing gap, and canopy growth (Vaughn et
al., 2015). Newly formed canopy gaps are the result of a height reduction event which
lead to heights below the threshold used to detect gaps. eanopy erosion is also a canopy height reduction event but resulting height above the gap detection height threshold. We defined canopy erosion as areas where crown losses result in a height reduction for which the posterior height is still above the height threshold used to detect a canopy gap. eanopy erosion happens when sub-canopy vegetation is present after the event that has led to canopy height reduction (Vepakomma et al., 2010). Filled gaps are due to the growth processes that lead to canopy closure, either through
lateral growth of branches or vertical growth of tree tops (Runkle et Yetter, 1987). Existing gap is a growth process that happens within gaps that don't close over the study period (Vepakomma et al. , 2011 ). Finally, canopy growth is the height growth process that happens outside gaps.
Canopy erosion and new canopy gaps were detected within height reduction areas (HRA). HRA were defined as the spatial clusters of at least 1 m of height reduction. The 1 m height change cutline was chosen arbitrarily as a way to reduce false positive errors that might arise from LiDAR measurement uncertainties. It was chosen visually and from knowledge of the LiDAR sensor imprecision. Within the HRA, individual pixels with a canopy height in 2013 at - or below - the gap detection height threshold (GDHT) were classified as new canopy gaps, and those above were classified as canopy erosion. The 2007 and 2013 CHM pixels that were both below the GDHT were classified as existing gaps, whereas those below that GDHT in 2007 but not in 2013 were classified as filled gaps. The remaining pixels were classified as canopy growth. We used GDHT varying from 1 to 10 m, with a 1 m increment to show the variability of our results to this subjective parameter (e.g. Lobo et Dalling, 2014). Analyses were performed separately for each GDHT. Treatments to produce canopy height models and detection of gaps were carried out in SAGA-GIS 2.2.3 (Conrad et al., 2015).
We investigated canopy dynamics processes by analyzing their occurrence and the respective canopy volume change they generate. We defined canopy volume as the volume between the top of the canopy to the ground and can be pictured as estimating the Gaussian volume of the 3D form shaped by a cloth draping the canopy. We then computed the relative canopy volume change as the volume difference between both volumes of the 2007 and 2013 CHMs over the volume of the 2007 CHM. This was computed for each canopy dynamics pattern at all GDHTs. Overall canopy volume change was then obtained by subtracting the sum of canopy volumes from negative canopy dynamics processes (gap creation, canopy erosion) from the sum of positive
processes ( canopy growth, gap closure, gap filling). A gap extinction rate was computed as the proportion of 2007 gap pixels that became non-gap over the study period divided by the time between the two flights (6 years) for the 10 GDHT.
1.4 Results
1. 4 .1 Mean canopy height
The mean canopy height in 2007 ranged between 14.0 m and 18.3 m, and in 2013 between 11.9 m and 16.8 m (Table 1.2), although all research sites had canopies detected at heights above 28 m. The 2007 mean canopy height of UMF sites was higher than that of managed sites, except for the site partially eut in 2008 (Table 1.2), which had a height distribution more typical of a single layer continuous canopy structure (Figure 1.1 ). This can also be seen in the lower standard deviation around the 2007 canopy height for that site compared to UMF or other managed forests. Mean canopy height increased from 2007 to 2013 on all sites, except for the two RSe sites (Table 1.2). Between 2007 and 2013, the greatest increase in canopy height occurred in the ose site in 1997 and was 25.0 cm/year, generating 10.6% of canopy volume increase in 6 years. In the UMF, the canopy was globally growing 11.7 cm/year, making it still the highest canopy in 2013. Standard deviation around the 2013 mean canopy height was higher in the RSe sites and most ose sites than in the UMF (Table 1.2). Standard deviations around the 2013 mean canopy height in the 1993 and 1995 ose sites lower than in the UMF sites indicate that canopy cover was more continuous, even though mean canopy height was lower. On the other hand, standard deviations around mean canopy height remained the same between 2007 and 2013 on the UMF sites.
Figure 1.1 shows that canopy height distribution changes from a strong bimodal distribution with a typical mode at low height (mostly below 2 m) following partial
Table 1.2 Mean canopy height (± 1 standard deviation), area and frequency of height reduction for the investigated study sites in 2007 and 2013. The frequency of negative canopy height change is the proportion of all the pixels that experienced a decrease in height of 1 meter or more between 2007 and 2013. RSe refers to recent selection cuts-stands that were eut between 2007 and 2013. ose means old selection cuts-stands that were eut the given year, before 2007. UMF stands for unmanaged forests-forests that have never been eut.
Site type Total Mean Mean Canopy Change in Frequency
and area canopy canopy volume mean of height
associated (ha) height in height in change canopy reduction
year of 2007 (m) 2013 (m) height
Eartial eut {cm/rear)
UMF 352 16.3±6.3 16.8±6.3 +3.9% 9.3±82.3 14.3% ose 1993 290 15.2±6.2 16.3±6.0 +6.7% 17.2±74.0 11.3% ose 1995 78 15.1±6.9 16.3±6.1 +8.3% 20.8±73.5 6.7% ose 1997 59 14.2±7.3 15.7±6.6 +10.6% 25.0±83.0 8.3% ose 2000 135 14.3±7.1 15.1±6.3 +5.6% 13 .3±82.3 11.8% ose 2004 160 14.0±7.5 14.5±7.3 +3.6% 8.3±93.0 15.5% RSe 2008 58 18.3±5.5 14.3±8.1 -22.0% -67.0±138 41.0% RSe 2012 47 14.2±5.5 11.9±6.6 -16.2% -38.3±114 34·.9%
cutting (RSe sites) to a unimodal distribution with a strong mode in the overstory canopy (OSe sites and UMF site). A similar low height mode can be observed in the ose sites (left sicle of the graph (2007). In 2013 (right sicle of the graph) for the ose sites, this low height mode is still perceptible but tends to shrink as the years since the eut increases. This is due to a low height growth of the vegetation that gradually reduced in frequency until a balance is reached, similar to the one o bserved in the 1993 ose site and in the UMF site.
We observed that, on the ose and UMF sites, height loss was caused by vegetation at heights at the highest mode of the 2007 height distributions, into the upper parts of the canopy, and to a lesser degree from intermediate heights (Figure 1.1 ). However, height loss ended up fairly homogeneously distributed throughout the lower,