UNIVERSITE DE SHERBROOKE
ETUDE DES EFFETS
NEUROCOMPORTEMENTAUX DE LA
NEUROTENSINE ET DE PEPTIDES APPARENTES
DANS UN MODELE ANIMAL DE LA MALADIE DE
PARKINSON
par
ROBERT RIVEST
Mémoire présenté à la Faculté de Médecine en vue de
l'obtention du grade de maître ès Sciences (M. Sc)
Permission bas been granted
to the National Library of
Canada to microf i lm this
thesis and to lend or sell
copies of the film.
The author (copyright owner)
has reserved other
publication rights, and
neither the thesis nor
extensive extracts from it
may be printed or otherwise
reproduced without his/her
written permission.
ISBN
L'autorisation a été accordée
à
la Bibliothèque nationale
du Canada de microfilmer
cette thèse et de prêter ou
de vendre des exemplaires du
film.
L'auteur (titulaire du droit
d'auteur) se réserve les
autres droits de publication:
ni la thèse ni de longs
extraits de celle-ci ne
doivent être imprimés ou
autrement reproduits sans son
autorisation écrite.
REMERCIEMENTS
Je désire rémercier sincèrement le Dr. François B. Jolicoeur pour m'avoir accepté dans son laboratoire. J'apprécie grandement l'extrême marque de confiance dont vous avez toujours témoigné envers moi, de même que le support scientifique exceptionnel lorsque j'en avais besoin ou simplement pour le plaisir de discuter. Grâce à votre générosité, j'ai eu à plusieurs reprises, l'occasion de côtoyer "le monde scientifique• et j'y ai pris goût.
Je suis également très reconnaissant envers Andréa Drumheller pour sa
participation directe et très intéressée au projet. Quelle gentillesse et générosité autant dans notre travail professionnel qu'à l'extérieur. Aussi, comme mes textes Anglais étaient bien écris, après qu'Andréa les eut repris.
À Daniel Gaudin ainsi qu'à Daniel Ménard, je leur doit l'excellent soutient et l'agréable entente qu'il y a au laboratoire.
Je remercie sincèrement le Dr. Dominico Régoli et le Dr. Paul Bédard pour avoir accepté de corriger cette thèse et ainsi, contribué à améliorer la qualité de cet ouvrage.
Je tiens également à souligner la colaboration du Dr. Serge St-Pierre qui nous a donné les peptides utilisés durant tout mes travaux.
TABLE DES MATIERES
page REMERCIEMENTS ... ... ... ... .... ... ... ... .... .... ... ... ... .... .
TABLE DES MATIERES... ... iii
LISTE DES ILLUSTRATIONS... ... vi
LISTE DES TABLEAUX... ... ... ... ... x
RESUME. ... .. ... xiii
SOMMAIRE... ... ... ... xiv
INTRODUCTION .... ... ... ... ... ... ... ... ... ... ... . 1- GENERALITES SUR LA NEUROTENSINE ... . 2-INTERACTIONS NEUROTENSINE-DOPAMINE... ... ... ... ... 10
3- LA MALADIE DE PARKINSON... 16
3.1- Les neuropeptides et la maladie de Parkinson... 20
3.2- La neurotensine et la maladie de parkinson... 22
4- LES MODELES ANIMAUX DE LA MALADIE DE PARKINSON... 23
MATERIEL ET METHODES ... .... ... ... 28
1- GENERALITES... ... ... ... ... ... .... ... 28
2- INJECTION DE LA 6-0HDA... 28
3- CANULATION DES RATS ET DES SOURIS. ... 30
3.1- Canulation intra-cérébrale... 30
3.2- Canulation IC... ... 32
3.3- Canulation IT... ... 32
4- INJECTION DES PEPTIDES ET DES DROGUES... 34 5- MESURES NEUROCOMPORTEMENTALES... ... ... 36 5.1- L'activité motrice... 36 5.2- La catalepsie... 38 5.3- La rigidité musculaire... 38 5.4- Les tremblements... 39
5.5- Les tremblements chez les souris... 39
6- HISTOLOGIE... 40
6.1 - Perfusion des animaux à la formaldéhyde... 40
6.2- Isolement et coupe des cerveaux au microtome... 40
6.3- Coloration des coupes de cerveau au crésyl violet... 41
7- ANALYSES NEUROCHIMIQUES... 42
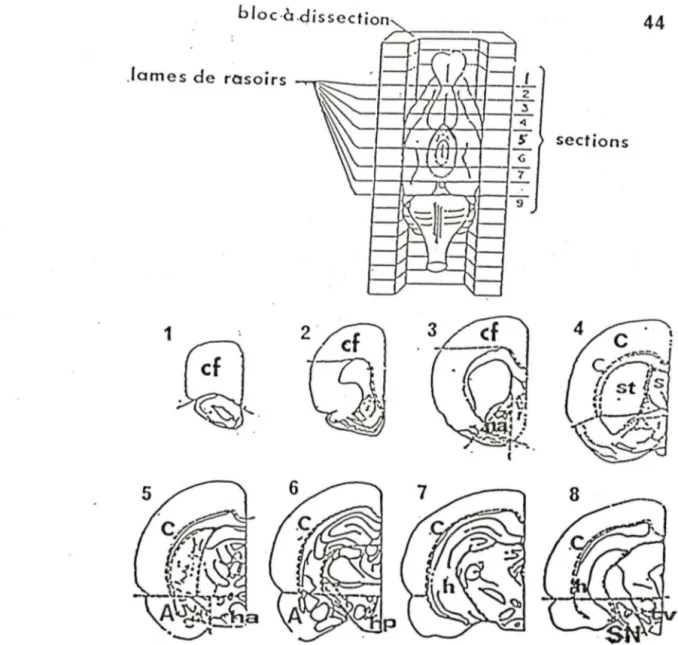
7.1- Dissections des régions cérébrales... 43
7.2- Homogénéisations des régions cérébrales... 45
7 .3- Détections électrochimiques des composés DA, DOPAC HVA, NE, 5-HT et 5-HIAA... 45
8- STATISTIQUES... 46
RESULTATS... 48
1- ETUDE DU MODELE ANIMAL DE LA MP... 48
1.1- Mesures neurocomportementales des effets de la 6-0HDA... 48
1.2- Mesures neurochimiques des effets induits par la 6-0HDA... 48
1.3- Etudes des effets neurocomportementaux de divers agents antiparkinsonniens... 51
2- EFFETS . DE LA NT ET DE PEPTIDES APPARENTES... 59
2.1- Effets de l'injection ICV de la NT... 59
V
2.2- Etude de structure-activité... 65
2.3- Etude des sites d'action de la NT... 74
DISCUSSION... 93
1- ETUDE DU MODELE ANIMAL DE LA MP... 93
2- ETUDE DES EFFETS DE LA NT ET DE PEPTIDES APPARENTES... 101
3- ETUDE DES SITES D'ACTION DE LA NT... 10 8 4- GENERALITES SUR LES EFFETS DE LA NT ... 110
CONCLUSION ... , ... : 112
BIBLIOGRAPHIE... ... 114
APPENDICE A- Liste des produits utilisés... 151
LISTE DES ILLUSTRATIONS
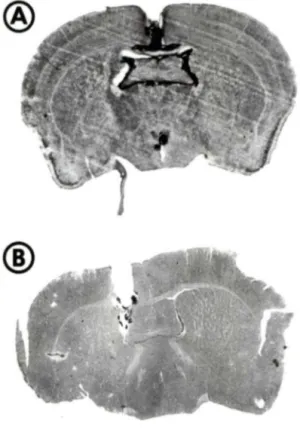
page Figure 1: Photographies de coupes histologiques de cerveau de
souris: évaluation des coordonnées stéréotaxiques du
ventricule latéral gauche... 3 5
Figure 2: Représentations photographiques des comportements
analysés chez les rats traités à la 6-0HDA dans le FVM... 3 7 Figure 3: Représentation schématique de la méthode de dissection
des cerveaux ... 44
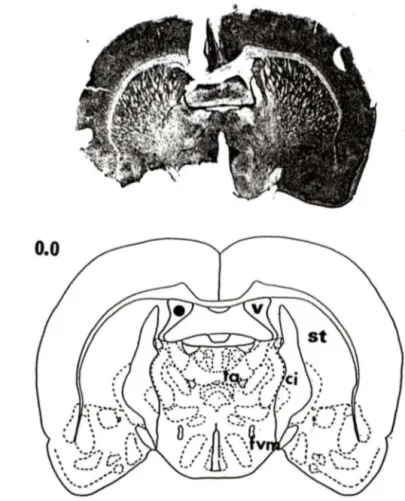
Figure 4: Vérification histologique d'une injection dans le FVM ... 49
Figure 5: Cinétique des effets de différentes doses d'apomorphine sc sur l'activité motrice chez des rats traités à la 6-0HDA
dans le FVM ... 56
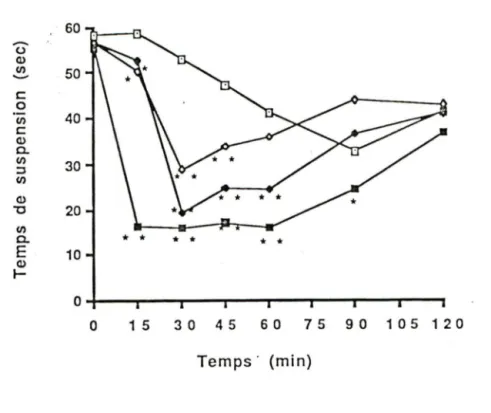
Figure 6: Cinétique des effets de différentes doses d'apomorphine sc sur le temps de suspension chez des rats traités à la 6-0HDA
dans le FVM ... 5 7
Figure 7: Cinétique des effets de différentes doses d'apomorphine sc sur
Figure 8: Effets dose-réponses de l'apomorphine sc sur les tremblements
induits chez des rats traités à la 6-0HDA dans le FVM ... 60
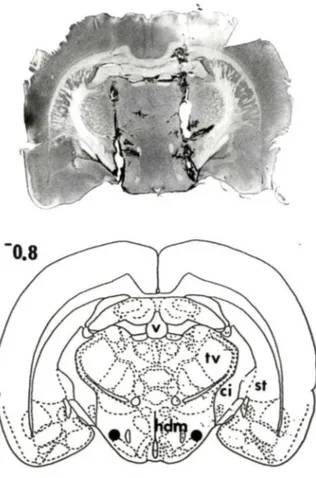
Figure 9: Vérification histologique d'une injection ICV ... 61
Figure 10: Cinétique des effets de l'administration ICV de différentes doses de NT sur le temps de suspension chez des rats
traités à la 6-0HDA dans le FVM ... ... 63
Figure 11 : Cinétique des effets de l'administration de différentes doses de NT sur la rigidité de la queue chez des rats
traités à la 6-0HDA dans le FVM ... 64 Figure 12: Effets dose-réponses de l'injection ICV de NT sur les
tremblements induits chez des rats traités à la
6-0HDA dans le FVM ... ... 66
Figure 13: Cinétique des effets de l'administration ICV de différentes doses de [D-tyr11 ]-NT sur le temps de suspension chez des
rats traités à la 6-0HDA dans le FVM ... 70
Figure 14: Cinétique des effets de l'administration ICV de différentes doses de [D-tyr 11 j-NT sur la rigidité de la queue chez des
rats traités à la 6-0HDA dans le FVM... 71 vii
Figure 15: Effets dose-réponses de l'injection ICV de [D-tyr 11 )- NT sur les tremblements induits chez des rats traités à la
6-0HDA dans le FVM... 72
Figure 16: Cinétique des effets de l'administration ICV de 30 µg de NT ou de [D-Tyr 11 )-NT sur le temps de suspension
chez des rats traités à la 6-0HDA dans le FVM ... 7 6
Figure 17: Vérification histologique d'une injection effectuée dans
le ST- ant {A) et le ST-post (8) ... 77
Figure 18: Vérification histologique d'une injection effectuée dans
le globus pallidus ... 7 8
Figure 19: Vérification histologique d'une injection effectuée dans
la SN {A) et le thalamus médio-ventral (8)... 79
Figure 20: Cinétique des effets de l'administration IC de différentes doses de NT sur le temps de suspension chez des
rats traités à la 6-0HDA dans le FVM ... 8 6 Figure 21: Cinétique des effets de l'administration IC de différentes
doses de NT sur la rigidité de la queue chez des rats
traités à la 6-0HDA dans le FVM... 8 7 viii
Figure 22: Effets dose-réponses de l'injection IC de NT
sur les tremblements induits chez des rats traités à la
6-0HDA dans le FVM ... 88
Figure 23: Cinétique des effets de l'administration IT de différentes doses de NT sur le temps de suspension chez des
rats traités à la 6-0HDA dans le FVM... 8 9
Figure 24: Cinétique des effets de l'administration IT de différentes doses de NT sur la rigidité de la queue chez des rats
traités à la 6-0HDA dans le FVM ... 90
Figure 25: Effets dose-réponses de l'injection IT de NT
sur les tremblements induits chez des rats traités à la
6-0HDA dans le FVM... ... 91 ix
LISTE DES TABLEAUX
page Tableau 1: Coordonnées stéréotaxiques des régions cérébrales
canul ée s ... 31
Tableau Il: Liste des solvants et des temps utilisés durant les colorations
histologiques au crésyl violet... ... 42
Tableau Ill: Sommaire des effets neurocomportementaux induits
par l'administration de 6-0HDA dans le FVM ... 50
Tableau IV: Concentrations régionales en DA, DOPAC et HVA,
exprimées en ng/mg de tissu ... 52
Tableau V: Concentrations régionales en NE, 5-HT et en 5-HIAA
exprimées en ng/mg de tissu ... 53
Tableau VI: Sommaire des effets neurocomportementaux de divers traitements pharmacologiques sur les rats lésés à
la 6-0HDA dans le FVM ... 54
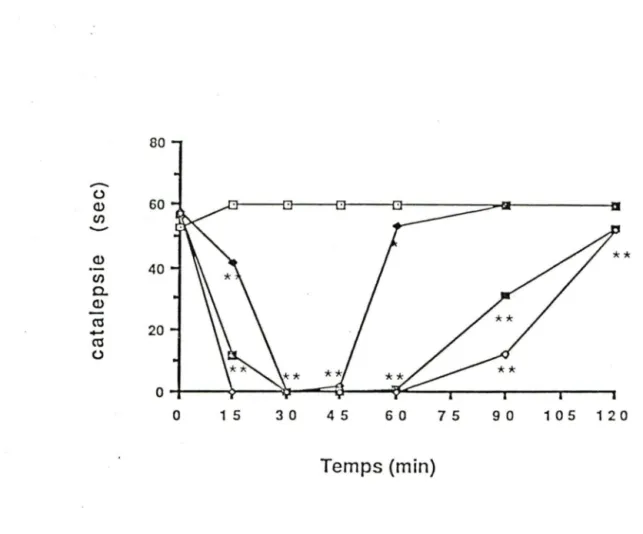
Tableau VII: Effets de l'administration ICV de 120.0 µg de NT sur la catalepsie et l'akinésie induit par un traitement à la
Tableau VIII: Effets de l'injection ICV de NT et de divers analogues et fragments sur le temps de suspension chez des rats traités à
la 6-0HDA dans le FVM... 6 7
Tableau IX: Effets de l'injection ICV de NT et de divers analogues et fragments sur la rigidité de la queue chez des rats traités à
la 6-0HDA dans le FVM... 69
Tableau X: Effets en pourcentage par rapport au temps=O de diverses doses de NT et de !D-tyr11 ]-NT administrés par la voie ICV sur le temps de suspension chez des rats traités à
la 6-0HDA dans le FVM ... 73
Tableau XI: Comparaison des doses seuils (µg) des effets de la NT et de l'analogue !D-tyr 11 ]-NT sur la rigidité de la queue et les tremblements induits par la 6-0HDA administrée
dans la FVM ... 75
Tableau XII: Effets de l'injection intra-régionale de NT
sur le temps de suspension chez des rats traités à
la 6-0HDA dans le FVM... 80
Tableau XIII: Effets de l'injection intra-régionale de NT
sur la rigidité de la queue chez des rats traités à
la 6-0HDA dans le FVM ... 82 xi
xii
Tableau XIV: Effets de l'injection intra-cérébrale de NT
sur le temps de suspension chez des rats traités à
la 6-0HDA dans le FVM ... 83
Tableau XV: Effets de l'injection intra-cérébrale de NT
sur la rigidité de la queue chez des rats traités à
RESUME
ETUDE DES EFFETS NEUROCOMPORTEMENTAUX DE LA NEUROTENSINE ET DE PEPTIDES APPARENTES DANS UN MODELE ANIMAL DE LA MALADIE DE PARKINSON
par
ROBERT RIVESTLes symptômes cardinaux de la maladie de Parkinson (MP) sont l'hypokinésie, la rigidité musculaire et les tremblements. Ils seraient principalement causés par à une diminution importante de la dopamine (DA) dans le striatum, faisant suite à une dégénérescence précipitée des corps cellulaires dopaminergiques au niveau de la substance noire (SN). Or, il appert que de fortes densités de récepteur à la neurotensine (NT) sont localisées dans la SN et que chez des patients atteints de la MP, une baisse marquée des récepteurs neurotensinergiques
y
est trouvée. Due à ces évidences expérimentales, il est possible que la NT puisse être impliquée dans l'étiologie eUou la symptomatologie de cette maladie.Au cours de ce projet de recherche effectué avec des rats, un modèle animal de la MP a été développé dans le but d'étudier les effets de la NT sur les symptômes reliés à cette maladie. Des rats ont été canulés dans le faisceau ventro-médian et ont reçu 4µ1 de 6-hydroxydopamine (6-0HDA) à une concentration de 6.5 µg/µI. Deux jours après le traitement, nous avons procédé à des mesures neurocomportementales de catalepsie, d'activité motrice, de rigidité musculaire (test de suspension et le test de la rigidité de la queue) et d'observations de tremblements. Tout ces symptômes furent fortement augmentés par le traitement sauf l'activité motrice qui a été diminuée très significativement. Les effets neurochimiques induits par le traitement dans diverses régions du système nerveux central ont été évalués au moyen d'analyses par chromatographie liquide à haute pression couplée à la détection électrochimique(HPLC-DE). Nous avons trouvé des baisses très marquées de la DA et de son métabolite, l'acide 3,4-dihydroxyphénylacétique et de même que de la norépinéphrine dans le ST, le noyau accumbens, l'amygdale et !'hypothalamus alors que l'acide homovanillique fut diminué dans les trois premières régions. Aucun changement ne fut observé sur les concentrations en sérotonine et en 5-hydroxyindolacétique dans les régions cérébrales étudiées. L'apomorphine et le L-dopa ont renversé les effets induits par la lésion alors que les deux anticholinergiques, artane et le benztropine ont produit une légère hausse de l'activité motrice. L'administration intra-cérébro-ventriculaire de 30 µg de NT a réduit significativement la rigidité musculaire et la présence de tremblements. Le (D-Tyr 11 J-NT s'est révélé environ 16 fois plus puissant que la NT dans le test de suspension et environ 33 fois plus efficace sur la diminution de l'apparition des tremblements. Par contre, le fragment NT 1•1
o
et l'analogue (Ala 11 J-NT ont été inactifs pour atténuer les deux symptômes. L'étude des sites d'action de la NT a montré que l'administration intra-cérébrale du peptide permet d'obtenir des effets variables selon les régions. Par contre, il semble qu'en général, l'injection de la NT dans les cavités telles que les ventricules latéraux, la cisterne magna et la thèque de la moelle épinière procure des effets plus marqués que lorsque le peptide est administré directement dans le tissu cérébral. En résumé, ces résultats suggèrent qu'un modèle animal de la MP a été obtenu à partir d'injection de 6-0HDA dans le faisceau ventro-médian. La NT semble être un peptide efficace pour réduire les symptômes induits par la neurotoxine. De plus, notre étude de structure activité nous indique que les effets du peptide ne seraient pas uniquement causés par l'administration de peptides quelconques puisque la NT 1 -1o
et l'analogue [Ala 11 ]-NT furent inactifs. Aussi, la tyrosine en position 11 possède un rôle important dans l'action du peptide.SOMMAIRE
L'objectif de ce projet de recherche a été : 1 ·- d'établir un modèle neurocomportemental de la maladie de Parkinson avec des rats; 2·- d'étudier les effets de la neurotensine et de quelques peptides apparentés dans ce modèle; 3 • - d'entreprendre les premières études des sites d'action du peptide.
Nous avons procédé au moyen de lésions neurochimiques induites au niveau du faisceau ventre-médian. La neurotoxine utilisée a été de la 6-hydroxydopamine, qui fut administrée bilatéralement selon les conditions suivantes; 26µg/4µ 1 durant un temps d'infusion de 4 minutes. Lorsque la préparation des animaux était faite en prévision de l'étude des effets intra-cérébraux des peptides, les rats ont été canulés durant la même chirurgie dans une des régions suivantes: ventricule latéral gauche, striatum postérieur, striatum antérieur, thalamus, substance noire, 3ième ventricule, 4ième ventricule, cisterna magna et dans la thèque de la moelle épinière. Entre la deuxième et la quatrième journée après la chirurg ie, nous avons procédé à des mesures neurocomportementales afin d'étudier les effets de diverses drogues et peptides, sur les comportements induits par la 6-hyd roxydopam ine . .
Les principaux symptômes de la maladie de Parkinson sont l'hypokinésie, la rigidité musculaire et les tremblements. L'état hypokinétique des animaux fut évalué par les tests de la catalepsie et de la mesure de l'activité motrice spontanée. Deux tests ont servi à évaluer la rigidité musculaire. En premier lieu, le test de suspension a consisté à mesurer le temps durant lequel les animaux peuvent demeurer suspendus par les pattes antérieures à une barre. La deuxième méthode consistait a soulever la queue d'environ 5 cm à partir de 2 cm de son extrémité avec une barre de 0.5 cm de diamètre, et à enregistrer le temps qu'elle tient sur la barre. Enfin, nous avons également procédé à l'observation de l'apparition des tremblements, qui étaient apparents lorsque l'on soulevait, par la queue, le train arrière des
XV animaux de quelques centimètres. Nous avons aussi étudié les changements neurochimiques de quelques amines et .métabolites, induit par le traitement neurotoxique, au moyen d'analyses par éhromatographie liquide à haute pression couplée à la détection électrochimique.
Les résultats obtenus ont été les suivants:
1 • - Nous avons étudié les effets neurocomportementaux et neurochimiques causés par l'administration de la 6-hydroxydopamine au niveau du faisceau ventre-médian. En comparaison au groupe témoin qui a reçu de la saline dans la même région, les animaux du groupe lésé ont montré des différences très marquées dans tout les tests comportementaux. Le traitement induit, a- une forte catalepsie , b- de la rigidité musculaire observée par les tests de suspension et de la queue, c- fait apparaitre des tremblements et d- diminu de manière très évidente l'activité motrice des animaux. Selon nos analyses neurochimiques, des baisses très importantes de la concentration (ng/mg tissu) en dopamine, acide 3,4-dihydroxyphénylacétique, acide homovanillique et en norépinéphrine ont été induites par la lésion dans les régions du striatum, du noyau accumbens, de !'hypothalamus et de l'amygdale. L'acide homovanillique fut diminué dans les trois premières régions. Aucune différence ne fut observée dans la substance noire. De plus, aucun changement ne fut observé sur les concentrations en sérotonine et en 5-hydroxyindolacétique dans les régions cérébrales étudiées.
2· - Des drogues à propriété dopaminergique telles que l'apomorphine et le L-dopa ont été efficaces pour renverser les effets comportementaux induits par la 6-hydroxydopamine. Par contre, l'artane et la benztropine, deux anticholinergiques, n'ont produit aucun effet, mis à part de légères hausses de l'activité motrice des rats.
xvi
3· - Dans notre modèle animal, l'administration de 30 µ g de neurotensine réduit significativement les symptômes de rigidité musculaire et atténue l'apparition des tremblements. La puissance et la durée des effets dépendent de la dose administrée mais durent généralement environ 60 minutes. Il semble que le peptide soit plus puissant à induire ses effets lorsqu'il est injecté dans les cavités du système nerveux central telles que les ventricules latéraux, la cisterna magna et la thèque de la moelle épinière plutôt que lorsqu'il est donné directement dans les régions cérébrales. L'analogue [D-Tyr 11 ]-NT s'est révélé plus puissant que la neurotensine pour atténuer les effets due à la 6-hydroxydo pam ine.
En résumé, un modèle neurocomportemental de la maladie de Parkinson peut-être obtenu suite à l'administration bilatérale de 6-hydroxydopamine au niveau du faisceau ventro-médian. Il en résulte des effets d'hypokinésie, de rigidité musculaire et de tremblements. Ce traitement se traduit par une diminution d'environ 80 % en dopamine, acide 3,4-dihydroxyphénylacétique et en acide homovanillique dans le striatum. L'administration intra-cérébrale de neurotensine atténue significativement les symptômes neurocomportementaux induits par la neurotoxine. Ces résultats viennent appuyer l'hypothèse que certains neuropeptides, en outre la neurotensine, pourraient être impliqués dans l'étiologie et/ou la symptomatologie de la maladie de Parkinson. De plus, ce projet apporte des évidences que des peptides apparentés à la neurotensine pourraient être développés en vue de leur utilisation éventuelle dans le traitement de cette maladie.
1
INTRODUCTION
1. GENERALITES SUR LA NEUROTENSINE
La découverte de la neurotensine (NT) est survenue de manière fortuite lors de la purification de la substance P à partir d'hypothalamus bovin par CARRAWAY et LEEMAN, ( 1973). Ceux-ci observèrent qu'un effluent de la colonne chromatographique distinct de celui de la substance P, produisait de l'hypotension, de la vasodilatation et de la cyanose lorsqu'injecté par voie intra-veineuse chez le rat. Le peptide fut ainsi nommé à cause de sa présence dans le système nerveux et à son effet hypotenseur. Plus tard, la NT a été également isolée à partir d'intestins de boeuf et humain (CARRAWAY et coll., 1978; HAMMER et coll., 1980). Selon CARRAWAY et LEEMAN (1975) il est composé de 13 acides aminés dont la séquence est la suivante : p-Glu-Leu-Tyr-G lu-Asn-Lys-Pro-Arg-Arg-Pro-Tyr-lle-Leu-OH. L'identification de la structure du tridécapeptide a rendu possible sa synthèse sur support solide (CARRAWAY et LEEMAN ,1975; FOLKERS et coll., 1976; RIVIER et coll., 1977; ST-PIERRE et coll., 1981). Par la suite, la production de NT synthétique a perm is d'entreprendre des études immunologiques, pharmacologiques et physiologiques. Une revue de la littérature des effets de la NT permet d'affirmer que le peptide possède des caractéristiques communes aux neurotransmetteurs eUou aux neuromodulateurs. Afin de mieux expliciter cette affirmation, je présente une revue générale des études de distribution, libération, métabolisme, de ces récepteurs et des principaux effets neurophysiologique s et neurocomportementaux de la NT.
La présence de NT immunoréactive (NT-1) dans le cerveau est un critère essentiel pour classer le peptide parmi les neurotransmetteurs. Toute les espèces étudiées ont démontré de l'immunoréactivité à la NT (CARRAWAY et LEEMAN, 1976; KOBAYASHI et coll., 1977;
2
UHL et coll., 1977; KATAOKA et coll., 1979; MANBERG et coll., 1982; BISSETTE et coll., 1982). Selon CARRAWAY et LEEMAN (1976), 10% de la NT-1 est présente dans le cerveau de rat et 90% dans le reste du corps dont 85% au niveau du système digestif. La NT est donc classée parmi les peptides apparentés au système nerveux et digestif. Dans le cerveau, !'hypothalamus et le tronc cérébral contiennent les plus grandes concentrations de NT-1 avec 70% du contenu total (CARRAWAY et LEEMAN, 1975). D'après la même étude, le thalamus, la glande hypophyse et le cervelet contiennent 11 , 1 et 0.8% respectivement. Toujours chez le rat, EMSON et coll. (1982) ont publié une étude détaillée de la distribution de NT-1. De façon similaire aux autres neuropeptides connus, la NT se distribue de manière hétérogène dans tout le cerveau. Les plus grandes concentrations sont localisées dans les noyaux de !'hypothalamus et le noyau strié. Des taux appréciables sont également trouvées dans les régions du système limbique, incluant le noyau central de l'amygdale, le septum, le noyau accumbens · et les tubercules olfactifs. Aussi, des taux considérables de NT-1 furent trouvés dans le tronc cérébral au niveau des noyaux habénulé, thalamus médian, noyau raphé dorsal, locus coeruleus, de la région périaquéductale grise, la substance noire (SN), les noyaux du faisceau solitaire et du faisceau du nerf trijumeau ainsi que dans la moelle épinière au niveau de la corne dorsale cervicale, lombaire et thoracique. Enfin, les plus petites concentrations sont trouvées dans le cervelet et le cortex. Cette distribution de NT-1 est semblable à celle trouvée dans le cerveau humain par MANBERG et coll. (1 982); EMSON et coll. (1985) ; MICHEL et coll. (1986) et SAKAMOTO et coll. (1987). Cependant, il est à noter que de grandes concentrations de NT-1 ont été trouvées dans la SN du cerveau humain comparativement aux concentrations rapportées dans cette région chez les autres espèces animales étudiées (MAN BERG et coll. , 1982; EMSON et coll., 1985). Dans la cellule nerveuse, de la NT-1 a été trouvée au niveau des corps cellulaires, des axones et dans les terminaisons axonales(UHL et coll. , 1979).
3
La première voie nerveuse neurotensinergique a été identifiée par UHL et SNYDER (1979). Elle origine du noyau central de l'amygdale et se termine dans le noyau de la strie terminale. Depuis cette étude, d'autres fibres neurotensinergiques ont été trouvées impliquant les régions cérébrales suivantes, de l'origine vers (->) la terminaison : hippocampe-cortex antérieur cingulaire (ROBERTS et coll., 1981), formation grise centrale et noyau solitaire -> noyau raphé (BEITZ, 1982); région du tegmentum ventral -> noyau accumbens (KALIVAS et JENNER, 1982); noyau central de l'amygdale -> noyau parabrachial (MOGA et GRAY, 1985); striatum (ST) -> noyau accumbens (SUGIMOTO et MIZUNO, 1987).
Puisque de la NT-1 est présente dans le cerveau des mammifères, il était prévisible que des récepteurs spécifiques au peptide puissent y être également trouvés. La liaison de la NT sur .les membranes neuronales possède les caractéristiques de haute affinité, saturabilité, réversibilité et spécificité (UHL et SNYDER, 1977; UHL et coll ., 1977; LAZARUS et coll., 1977; KITABGI et coll., 1977). Alors que différents laboratoires ont trouvé que la NT se lie à une seule classe de récepteur (GOEDERT et coll., 1984; NAKAGAMA et coll., 1984; VILLER, 1985), le groupe de MAZELLA et coll. (1983) ont trouvé deux sites de liaison pour le peptide. L'étude des classes de récepteur à la NT fut par la suite reprise par SCHOTTE et coll. (1986, 1987). Selon ces études, les deux sites de liaison sont saturables, stéréospécifiques et ont des affinités pour le peptide dans l'ordre du nanomolaire. Par contre, le site numéro 1 montre une distribution homogène dans tout le cerveau, il est saturable à des concentrations (15nM) qui sont plus élevées que le deuxième site (4nM) et n'a pas encore été trouvé chez des espèces autres que les rats et les souris. Pour ces raisons, ce site de liaison est considéré comme un "accepteur· ou un site de reconnaissance à la NT sans fonction physiologique particulière. Le deuxième site possède toutes les caractéristiques pour être classé comme un véritable récepteur puisque le
4
peptide s'y lie avec une haute affinité, à un faible seuil de saturation et présente une distribution hétérogène dans le cerveau. Enfin, un seul site de liaison de la NT a été trouvé dans les membranes de cerveau humain (KANBA et coll., 1986).
La distribution de la liaison de la NT dans le cerveau a été étudiée par les méthodes de liaisons classiques (LAZARUS, 1977; KAN BA et coll., 1986) et aussi de manière plus détaillée au moyen de la technique de l'autoradiographie (YOUNG et KUHAR 1981; OUIRION et coll., 1982; SARRIEAU et coll., 1985). Comme il a été observé pour la NT-1, les récepteurs du peptide sont distribués partout dans le cerveau, à des concentrations variables selon les régions. Selon YOUNG et KUHAR (1981); QUIRION et coll. (1982), la distribution des récepteurs de la NT peut-être comparée pour plusieurs régions du cerveau, à la distribution de NT-1. Cependant, une analyse détaillée de la comparaison entre la localisation des récepteurs et de la NT-1 révèle qu'il y a de grandes disparités (HERKENHAM, 1987). Ainsi, plusieurs régions contenant des fortes densités de récepteur ont très peu d'immunoréactivité au peptide (SN , cortex frontal , septum dorsolatéral, cervelet) alors que l'inverse survient dans le noyau central de l'amygdale, le septum ventrolatéral, le striatum médian et le noyau accumbens. Dans le ST de chat, GOEDERT et coll. (1984) ont trouvé une corrélation négative entre la densité de la NT-1 et la liaison à son récepteur. Il semble donc qu'il y a des disparités entre la localisation des récepteurs à la NT et la NT-1. Selon GOEDERT et coll. (1984) ; HERKENHAM (1987) , les peptides auraient possiblement la propriété d'agir à distance par transport trans-synaptique et nous pouvons supposer qu'il en va de même pour la NT.
Les mécanismes exacts de la synthèse de la NT ne sont pas très bien connus. Cependant, nous savons que des ARNm spécifiques pour des neuropeptides ont été trouvés (SUTCLIFFE et MILNER, 1984; LARHAMMAR et coll., 1987). Une étude récente de PARKER et coll. (1986) a apporté des évidences qu'il y aurait de l'ARNm spécifique pour
5
la NT. Donc, puisqu'il s'agit d'un peptide et parce que de l'ARNm a été trouvé , il parait concevable que la synthèse de la NT s'effectue par le biais de processus communs aux protéines (VINCENT et SIMONET, 1986). Selon ce mécanisme, la synthèse peut-être divisée en deux grandes étapes. La première est similaire pour toutes les protéines cellulaires. Elle concerne la transduction de l'ADN en ARNm et la traduction de l'ARNm en protéine au niveau des ribisimes. Il survient alors un processus de ségrégation des séquences protéiques qui sont coupées lors de la synthèse au niveau du réticulum endoplasmique. Selon les même auteurs, il semble qu'il puisse y avoir une maturation extracellulaire durant laquelle les pro-peptides sont coupés en fragments actifs correspondant aux neuropeptides.
L'application de NT sur des préparations in vitro ou in vivo affecte l'excitabilité neuronale des cellules (MILETIC et RANDIC, 1979; ANDRADE ET AGHAJANIAN, 1981; HENRY, 1982; STANZIONE et ZIEGLGÀNSBERGER, 1983; SHI et coll., 1987). Des effets neuroexcitateurs ( ANDRADE et AGHAJNIAN , 1981; HENRY, 1982; SHI et coll., 1987) et neuroinhibiteurs (MILETIC et RANDIC, 1979; STANZIONE et ZIEGLGÀNSBERGER, 1983) ont été rapportés selon les régions étudiées. Parfois, les deux effets ont été trouvés dans la même région (MILETIC et RANDIC, 1979; HENRY, 1982). De plus, des études effectuées par IVERSEN et coll. (1978) ainsi que MAEDA et FROHAN (1981), ont démontré que de la NT-1 pouvait être libérée à partir de tranches d'hypothalamus de rat, par une dépolarisation neuronale au potassium et que cette dépolarisation était dépendant du calcium.
Selon CHECLER et coll. (1983), MCDERMOTT et coll. (1982, 1986), la NT peut être métabolisée à partir de fractions membranaires, par des protéases qui se trouvent autant dans la fraction particulaire que soluble, cette dernière étant plus active que la première. Les sites de coupures enzymatiques les plus évidents se situent aux liens peptidiques
6
suivants: Arg8-Arg 9 , Pro 1o_Tyr11 et Tyr 11_lie12. Les enzymes impliquées seraient une métalloendopeptidase qui agirait au niveau de la liaison Arg8-A rg 9 alors que les liens Pro10_Tyr11 et Tyr11_11e12 seraient coupés par une enképhalinase. Il en résulte les fragments peptidiques N-trerminaux NT1_8, NT1_10 et NT1_11 alors que seulement le fragment C-terminal NT 9_ 13 a été détecté. Ce fragment est par ailleurs rapidement dégradé ce qui permet de libérer la tyrosine en position 11. La tyrosine provient également de la dégradation des autres fragments C-terminaux tel que la NT 11_ 13. Le fragment NT 1_ 10 est rapidement métabolisé en NT 1_8 par l'enzyme de conversion de !'angiotensine. Selon CHECLER et coll.(1986) l'enzyme qui est en grande partie responsable de la coupure au niveau du lien Pro 1O-Tyr11 serait une métalloendopeptidase distincte des enzymes déjà connues. Comme pour les autres peptides, il semble qu'il n'y a pas une enzyme spécifique à la dégradation de la NT. Ainsi, d'après TURNER et coll. (1985) il y aurait un nombre limité de peptidase pour un grand nombre de peptides. Le rôles de ses peptidases serait dépendant de leur localisation subcellulaire et cellulaire plutôt que de leur spécificité à un peptide en particulier. Enfin, mentionnons que des études du métabolisme in vivo de la NT après injection intra-veineuse chez des rats (ARONIN et coll., 1982) et chez l'homme (LEE et coll., 1984) ont confirmé les sites de coupures enzymatiques déjà obtenus in vitro. Les fragments obtenus NT 1 _8 et NT 1_1 1, se sont révélés biologiquements inactifs dans les essais biologiques. En résumé, il semble donc que le mécanisme de bioinactivation de la NT en fragments N-terminaux biologiquements inactifs s'effectu par des enzymes non-spécifiques au peptide lui-même mais reconnaissant des séquences peptidiques particulières. Les fragments C-terminaux semblent être rapidement dégradés en entier.7
neurotransmetteur, il est essentiel qu'il induise des effets neurophysiologiques après avoir été administré in vivo. Tout d'abord, NEMEROFF et coll. (1977), ont démontré indirectement que le peptide ne passe pas la barrière hémato-encéphalique puisque l'administration périphérique de la NT ne reproduisait pas les effets physiologiques obtenus par une injection dans le cerveau. Le premier changement neurophysiologique observé après administration centrale de NT fut la baisse de température corporelle chez des rats et des souris (BISSETTE et coll., 1976; NEMEROFF et coll., 1977; CLINESCHMIDT et MCGUFFIN, 1977; JOLICOEUR et coll., 1980). Selon JOLICOEUR et coll. (1980) cet effet est dépendant de la dose et dure environ deux heures. Par la suite, il fut démontré que non seulement la puissance mais également la durée des effets dépendent de la quantité administrée (JOLICOEUR et coll., 1984). Selon cette étude, l'administration ICV d'une faible dose (0.47 µg) de NT chez le rat a provoqué une diminution de la température corporelle de 1.4·c
et ce, pendant une période de 30 minutes. Par contre, l'administration d'une dose plus élevée (60 µ g) a produit une baisse de la température corporelle de 4.2'C pour une période de 150 minutes. L'injection intra-cérébrale de NT a été utilisée afin d'identifier les régions où le peptide serait plus susceptible d'agir. MARTIN et coll. (1981) ont administré 2.5 µ g de NT dans 135 régions du cerveau chez des rats. Ils ont obtenu des réductions significatives de la température corporelle dans 63 des régions étudiées. À la même période, KALIVAS et coll. (1982) ont également étudié les effets de l'injection intra-cérébrale de 2.5 µg de NT sur la température corporelle du rat. Ils ont analysé 223 régions avec lesquelles ils ont obtenu des effets variables selon les sites d'administration. Brièvement, les sites d'injection qui ont donné les plus fortes baisses de température ont été les suivants; la région préoptique médiale, !'hypothalamus antérieur et dorsomédial, la région périaquéductale grise, le thalamus ventral et la région du tegmentum ventral (MARTIN et coll., 1982; KALIVAS et coll., 1983).8
Selon les tests employés, la NT induit une baisse du tonus musculaire. Ceci fut rapporté par OSHBAR et coll. (1979), chez des souris qui étaient injectées avec 30µg de NT par la voie intra-cisternale (IC). Selon JOLICOEUR et coll. (1981), des diminutions du tonus musculaire sont obtenues chez des rats soumis au test de la traction après une injection ICV de 1.8µg ou plus. Par contre, la même étude a montré qu'aucun effet n'est obtenu avec le test de suspension et cela jusqu'à des doses de 120µ g injectées par la même voie d'administration.Des diminutions de l'activité motrice spontanée induites par la NT ont également été rapportées (NEMEROFF et coll., 1976; JOLICOEUR et coll., 1981). Selon cette dernière étude effectuée chez le rat, la dose seuil était de 1.8 µ g alors que les effets les plus puissants furent obtenus avec 7.5 µg du peptide. Au delà de cette dose, l'amplitude des effets a semblé diminuer avec l'élévation de la dose. Cela suggère que l'effet induit par la NT est exprimé par une courbe dose-réponse biphasique. Les études effectuées par VAN WIMERSMA et coll.(1982) ont montré que c'est l'activité motrice générale, incluant les stéréotypies, qui est diminuée par le peptide. Chez la souris, des injections IC de 25 pg de NT suffisent pour réduire l'activité motrice (CLINESCHMIDT et coll. ,1979). Lorsque le peptide est administré ICV, MEISENBERG et coll. (1985) ont évalué la dose seuil à 25 ng. L'étude détaillée des effets de la NT sur l'activité motrice suite à des injections intra-cérébrales a été effectuée par KALIVAS et coll. {1982, 1984). Les résultats ont montré que de l'hypoactivité ou de l'hyperactivité peuvent être obtenues selon les sites d'administration. Des augmentations de l'activité motrice sont produites principalement lorsque la NT est injectée au niveau ventromédial du mésencéphale (KALIVAS, 1984).
L'influence de la NT sur la réactivité des animaux soumis à des stimuli nociceptifs a été également étudiée. Il a été rapporté que le peptide induit des effets antinociceptifs chez la souris (CLINESCHMIDT et coll. ,1977; NEMEROFF et coll., 1979; CLINESCHMIDT et coll. ,
9
1979; SHIBATA et coll.,1987). Chez les rats, la NT ne semble pas altérer la réactivité des animaux lorsqu'elle est injectée dans les ventricules latéraux (COWAN et coll. , 1982; KALIVAS et coll., 1982). Cependant, des effets antinociceptifs ont été rapportés avec des injections de peptide IC. Il semble que la NT soit efficace pour produire de l'analgésie lorsqu'elle est administrée au niveau des régions reconnues pour être impliquées dans la transmission des sensations nociceptives telles que la substance grise périaqueducale, la région rostroventromédiale du bulbe(FANG et coll., 1987), le plancher du quatrième ventricule et la région pontine de la formation réticulée (KALIVAS et coll., 1982). La relation entre la présence de NT-1 dans la région périaquéductale grise et les effets analgésiques du peptide a été soulevée récemment par SHIPLEY et coll. (1987). Les mécanismes précis des effets antinociceptifs de la NT demeurent inconnus à l'heure actuelle. Cependant, BEHBEHANI et coll. (1984; 1987) ont proposé que la NT pourrait avoir une action excitatrice sur les neurones de la région périaqueducale grise ce qui pourrait cause l'activation des systèmes inhibiteurs de la douleur médiés par le noyau raphé.
Des effets périphériques de la NT sont aussi obtenus suite à son injection centrale. Selon OSUMI et coll. (1978), l'injection ICV de NT (1 à 10 µg) réduit le taux basal des sécrétions gastriques. Suite à cet effet, ORLANDO et coll. (1985); NEMEROFF et coll. (1986), ont étudié le rôle de la NT ICV sur l'apparition d'ulcères gastriques chez des rats placés en situation de stress intense. Selon RAY et coll. (1987), des injections de NT dans le noyau central de l'amygdale pourraient également ralentir la formation d'ulcères gastriques induits par l'immobilisation forcée des rats. L'influence de la NT sur la pression artérielle a donné des résultats contradictoires. Alors que les groupes de SUMNERS et coll. ( 1982) et SH IDO et coll. ( 1985) ont observé un effet hypertenseur chez des rats non anesthésiés, RIOUX et coll. (1981) et OUIRION et coll. (1981) ont trouvé un effet hypotenseur suite à l'injection ICV du peptide. Cette disparité dans les résultats est peut-être due aux
1 0
différences expérimentales puisque OUIRION et coll. ( 1981) ont utilisé des animaux anesthésiés .
En résumé, il semble que la NT est synthétisée et libérée à partir de neurones; le peptide se lie à des récepteurs qui lui sont spécifiques, il est métabolisé et induit des effets électrophysiologiques, neurophysiologiques, neurochimiques et neurocomportementaux. De plus, il existe des voies neurotensinergiques dans le cerveau. Toutes ces évidences suggèrent que le peptide peut être considéré comme un neurotransmetteur ou un neu rom odu 1 a te u r.
2. INTERACTIONS NEUROTENSINE-DOPAMINE
Depuis plusieurs années, il apparait de plus en plus évident que si les neurotransmetteurs semblent posséder un profil d'action neuropharmacologique unique à chacun (LAJTHA, 1985), en aucun cas des fonctions physiologiques ou des états pathologiques ont pu être attribués à l'action spécifique d'un seul neurotransmetteur. Il est plus que probable que les réponses physiologiques ou pathologiques résultent plutôt des innombrables intéractions entre de multiples neurotransmetteurs. La NT ne faisant pas exception, des intéractions du peptide ont été rapportées avec la norépinéphrine (NE) (FUXE et coll., 1984; DRUMHELLER et coll., 1986, !'acétylcholine (GRIFFTHS et coll., 1986; JOLICOEUR et coll., 1987; FREY et coll., 1988), le TRH (NEMEROFF et coll., 1978), mais surtout avec la dopamine (DA) (NEMEROFF et coll., 1982, 1984, 1985, 1986; PRANGE JR et coll., 1982; QU IRION ,1983; GOEDERT, 1984; JOLICOEUR et coll., 1985).
Les premières observations qui ont conduits à l'hypothèse des intéractions NT-DA proviennent d'une étude de BISSETTE et coll. (1976) au cours de laquelle ils ont raporté l'effet du peptide sur la température corporelle (hypothermie) suite à son injection dans le système nerveux. En premier lieu, les mêmes auteurs avaient suggéré que cet effet n'était
1 1
pas altéré par des agonistes cholinergiques, noradrénergiques ou dopaminergiques. Cela supposait que le peptide n'agissait pas de concert avec les autres neurotransmetteurs pour produire son action. Par contre, ils ont par la suite observé qu'une baisse de l'activité dopaminergique induite par injection de 6-0HDA, augmentait l'hypothermie induite par la NT (NEMEROFF et coll., 1978, 1980). Durant la même période, NEMEROFF et coll. (1977) ont montré que la NT augmentait, comme le font plusieurs neuroleptiques, l'effet sédatif des barbituriques et de l'alcool. Enfin, de manière similaire aux neuroleptiques, il a été démontré que l'injection centrale du peptide induisait une baisse de l'activité motrice spontanée (NEMEROFF et coll., 1977; CLINESCHMIDT, 1979; JOLICOEUR et coll., 1981, 1982; WIMERSMA et col., 1984; CADOR et coll., 1985; MEISENBERG et SIMMONS, 1985) et une diminution du tonus musculaire (OSHBAR et coll.,1979; JOLICOEUR et coll., 1981). Puisque les quatre effets neurocomportementaux de la NT ci-haut mentionnés sont comparables à ceux produits par des neuroleptiques, il fut donc suggéré que le peptide pourrait avoir les propriétés antidopaminergiques endogènes (NEMEROFF, 1980). L'intérêt manifesté à l'égard de la NT depuis la fin des années 1970 n'est certe pas étranger à ces propriétés antidopaminergiques. Cela a conduit plusieurs laboratoire à étudier plus à fond cette intéraction NT-DA.
Tout d'abord, les premières études portant sur la distribution de la NT-1 avaient déjà montré, sans en noter l'intérêt, que le peptide était présent en concentration importante dans plusieurs régions des systèmes dopaminergiques mésolimbiques et nigro-striés (CARRAWAY et LEEMAN, 1976; UHL et SNYDER 1977; KOBAYSHI et coll., 1977). Par contre, les rapports qui ont suivi n'ont pas omis de spécifier cet aspect (UHL et SNYDER 1979; EMSON et coll., 1982, 1985; MICHEL et coll., 1986; SAKAMOTO et coll., 1987; MAI et coll ., 1987). La localisation des récepteurs de la NT analysée par autoradiographie a également montré qu'une intéraction NT-DA devrait être considérée. En utilisant des
12
tranches de cerveau de rat, YOUNG et . coll. (1981) ainsi que OUIRION et coll. (1982) ont trouvé de très fortes densités de récepteurs à la NT au niveau de la SN et du tegmentum ventral d'où origine les importants faisceaux dopaminergiques mésolimbiques et nigrostriés. Plus récemment, SARRIEAU et coll. (1985), KANBA et coll (1986), MOYSE et coll. (1987) ont présenté la distribution des récepteurs de la NT dans le cerveau humain. En plus de trouver de fortes concentrations de récepteurs du peptide dans la SN et le tegmentum ventral, ils ont aussi observé de fortes densités de liaisons dans le noyau accumbens, putamen et de l'amygdale. De plus, les récepteurs de la NT pourraient être situés sur les neurones dopaminergiques puisqu'une injection de 6-0HDA (6.5µg/2µ1) dans la SN induit une perte de 90% des récepteurs de la NT (PALACIOS et coll., 1981). Au niveau du ST, GOEDERT et coll. (1984) ont évalué qu'approximativement 30% des récepteurs neurotensinergiques étaient situés sur des terminaisons dopaminergiques. Enfin, QUIRION et coll.(1985) ont montré que les récepteurs à la NT étaient surtout localisés sur des cellules dopaminergiques pré-synaptiques du faisceau nigro-strié et post-synaptiques de la voie mésolimbique. Il appert donc que la distribution de la NT-1 et des ses sites de liaison viennent appuyer l'hypothèse d'une intéraction NT-DA.
Des évidences de Co-localisation entre la NT et la DA sont encore rares. Selon IBATA et coll. (1983, 1984) et HÔKFELT et coll.(1984), la plupart des cellules immunoréactives à la NT du noyau arqué contiennent aussi de la DA. D'après HÔKFELT et coll. (1984), il y aurait de la co-localisation NT-DA au niveau de la région A 12 du faisceau tubéro-infundibulaire. S'il y a encore que très peu d'exemples de coexistence entre la NT et la DA, de l'avis de EMSON et coll. (1985) cela serait du au fait que la co-localisation pourrait être confinée à des régions très discrètes. Quoi qu'il en soit, les quelques preuves qui existent déjà viennent renforcer l'hypothèse de l'intéraction NT-DA.
13
spontanée des rats, le peptide devrait . également diminuer l'hyperactivité comportementale induite par des agonistes dopaminergiques. Ceci fut rapporté par NEMEROFF et coll. (1980) lorsqu'ils ont trouvé une atténuation significative de l'hyperactivité locomotrice induite par l'amphétamine, le méthylphénidate ou la cocaïne suite à l'injection IC de NT. Plusieurs autres études ont confirmé l'effet inhibiteur de la NT sur l'hyperactivité comportementale induite par des agonistes dopaminergiques (ERVIN et col. , 1981; JOLICOEUR et coll., 1981 , 1983, 1985; NEMEROFF et coll., 1983; COSTALL et .coll. , 1984; KALIVAS et coll., 1984; SKOOG et coll., 1986; GAUDIN et coll., 1987).
L'étude des effets neurochimiques de la NT a également produit des évidences que le peptide altère les fonctions dopaminergiques. Ce fut tout d'abord GARCIA-SERVILLA et coll. 1978, puis WIDERLOV et coll.1982, REECHES et coll. 1983, NEMEROFF et coll. 1983 qui ont démontré que des injections intra-cérébrales de NT induisaient des augmentations dose-dépendantes des deux principaux métabolites de la DA, soit l'acide homovanillique (HVA) et l'acide 3,4 ~ dihydroxyphénylacétique (DOPAC) au niveau de
quelques terminaisons dopaminergiques comme le noyau accumbens, les tubercules olfactifs et le ST. Nous avons récemment démontré que les rapports DOPAC/DA ou HVA/DA, mesurés par chromatographie liquide à haute pression couplée à la détection électrochimique (HPLC-DE), sont augmentés dans plusieurs régions du cerveau (ST, accumbens, amygdala, tegmentum ventral), suite à des injections ICV de 7.Sµg et de 30µg de NT (DRUMHELLER et coll., 1987). Cela suggère que la NT induit une augmentation du métabolisme de la DA dans ces régions. Dernièrement, une étude in vivo des effets de la NT sur la concentration en DA dans l'espace extra-cellulaire du noyau accumbens, effectuée par dialyse intra-cérébrale couplée à la détection électrochimique, a montré une augmentation dose-dépendante de la DA après injection ICV du peptide (RUGGERI et coll., 1987). Le même résultat a été obtenu par BLAHA et coll. ( 1987) alors que ces derniers ont
14
mesuré les amines par voltammétrie. Utilisant la même technique, FORD et MARSDEN, (1987) ont rapporté que l'injection de 1 µg de NT dans la région du tegmentum ventral induisait une libération de DA dans le noyau accumbens ipsilatéral. Cet effet d'activation du système mésolimbique suite à l'injection de NT dans le tegmentum ventral est connu depuis les travaux de KALIVAS et coll.(1983) et a été confirmé par CADOR et coll. (1986). Ces auteurs ont observé qu'une augmentation de l'activité exploratrice des animaux était obtenue après injection de NT dans le tegmentum ventral (KALIVAS et coll., 1983, 1985; CADOR et coll., 1986). Les mécanismes proposés pour expliquer ces effets inusités ont été que le peptide pourrait agir directement sur la cellule dopaminergique pour diminuer le seuil de dépolarisation et ainsi faciliter l'activation neuronale, ou encore qu'il bloquerait l'effet de la DA sur les récepteurs dendritiques ou axoniques et, de cette manière, induirait une augmentation de l'activité neuronale (KALIVAS et coll., 1983).Les études électrophysiologiques ont confirmé les effets excitateurs de la NT dans cette région de même que dans la SN (ANGRADE et coll., 1981 ). Des résultats semblables ont été obtenus par SHI et coll. (1987) avec en plus, des évidences que la NT inhibe directement l'effet de la DA pré-injectée dans la SN.
Des augmentations de l'activité dopaminergiques suite à l'injection intra-hypothalamique de la NT ont aussi été suggérées par HAWKINS et coll. (1986) lorsqu'ils ont démontré que l'effet aphagique du peptide était augmenté avec l'administration IP d'agonistes dopaminergiques. Bien que cela demeure très hypothétique, les auteurs ont proposé que la capacité de la NT à induire de l'aphagie pourrait être médiée par son abilité à augmenter l'activité dopaminergique. Enfin, GLIMCHER et coll. (1984, 1987), ont observé que la NT induisait un effet de renforcement positif dans des tests d'auto-injection.
15
chroniques des neuroleptiques sur les concentrations en NT-1 dans le cerveau. Toutes les études ont rapporté des augmentations en NT-1 dans le noyau accumbens et le ST après administration de neuroleptiques (GOVONI et coll., 1980; GOEDERT et coll., 1985; DE CEBALLOS et coll., 1986; FREY et coll., 1986). Aussi, UHL et KUHAR (1984), ont observé que l'administration chronique de neuroleptiques induisait des augmentations de la densité des récepteurs de la NT dans la SN.Dans le but de mieux caractériser cette cc-modulation NT-DA, AGNATI et coll., (1983, 1985), ont étudié, dans un premier temps, les effets de la NT sur la liaison de l'agoniste dopaminergique, 3H-n-propylnorapomorophine, sur les membranes mésolimbiques provenant du noyau accumbens et des tubercules olfactifs et, dans un deuxième temps, les effets de la DA sur la liaison de la NT sur ces mêmes tissus. Leurs résultats ont révélé qu'il semble exister une cc-modulation récepteurs-récepteurs puisque la NT réduit l'affinité de la liaison de l'agoniste dopaminergique alors que la DA diminue l'affinité de la liaison de la NT tout en augmentant le nombre de récepteurs (Bmax). Le même effet de la NT sur l'affinité de la liaison dopaminergique D-1 et D-2 fut récemment démontré par ISHIBA et coll. (1987), ainsi que par VON EULER et FUXE, (1987). Selon AGNATI et coll. (1985), il semble que l'activation dopaminergique induise une augmentation de la transmission neurotensinergique en élevant le nombre de sites de liaison de la NT, ce qui pourrait par la suite, produire une baisse de l'affinité de la liaison sur les récepteurs dopaminergiques. Des évidences appuyant cette cc-modulation récepteur-récepteur ont été publiées dernièrement par FUXE et coll. ( 1986). Cette étude montre que lorsque des récepteurs dopaminergiques du ST ont été rendus supersensibles suite à une lésion de la SN par injection de 6-0HDA, cela induisait une diminution dans l'affinité de la liaison de la NT dans cette région (FUXE et coll., 1986).
1 6
nerveux. Bien que cela nécessite encore d'être étudié, il semble qu'il ne s'agisse pas d'une simple modulation de la NT sur les systèmes dopaminergiques mais plutôt, d'une co-modulation entre le peptide et !'amine.
3. LA MALADIE DE PARKINSON
C'est en 1817 que le médecin Anglais James Parkinson décriva les symptômes de la "paralysie agitée', qui fut par la suite appelée en 1862, "maladie de Parkinson (MP), d'après l'idée du médecin français, Jean Martin Charcot. Lorsque comparé aux autres désordres neurologiques, nous pouvons affirmer que la symptomatologie et à la neuropathologie de la MP, est aujourd'hui relativement bien connue. On reconnait à la MP, trois symptômes cardinaux: soit l'hypokinésie, la rigidité musculaire et les tremblements (SCHULTZ, 1984). Le terme ' hypokinésie' réfère à un groupe de symptômes caractérisés par; 1 ' - la diminution de la capacité d'initier des mouvements et dans certains cas, l'immobilité quasi totale, soit l'akinésie; 2'- une diminution de la vitesse des mouvements, spécialement lors de l'accomplissement de mouvements répétitifs volontaires, soit la bradykinésie; 3' -la perte de !'habilité à modifier ou arrêter un mouvement qui a été initié. L'hypertonie musculaire est généralisée. Elle affecte les muscles extenseurs et fléchisseurs mais une prédominance proximale est observée. Les tremblements sont rhythmiques et alternatifs entre un groupe de muscles et ses antagonistes. Ce sont les muscles distaux qui sont principalement affectés. Il est plutôt courant qu'ils se manifestent de manière asymétrique, c'est-à-dire, qu'un coté du corps est plus affecté que l'autre. La fréquence des tremblements est d'environ 3 à 5/seconde. Ils disparaissent durant le sommeil et peuvent augmenter jusqu'à 1 O à 12/sec avec le stress et l'anxiété qui peuvent provenir d'une simple commande d'initier un mouvement volontaire. Une fois que le mouvement est commencé, les tremblements
1 7
vont généralement cesser. C'est pourquoi ils sont généralement appelés tremblements de repos. Cependant, il peut survenir qu'un patient puisse présenter des tremblements de type postural ou d'action (survenant lors d'une action). Selon HADAR et coll. (1987), les trois types de tremblements (de repos, postural et d'action) seraient tous dus à une même neuropathologie de sorte que nous pouvons penser qu'un seul type de tremblements caractérise la MP.La MP est associée à certaines altérations morphologiques cérébrales bien spécifiques chez tous les patients. D'autres changements peuvent être apparents mais ceux-ci sont généralement variables selon l'histoire des individus, la cause et la progression de la maladie. Ainsi, chez tout les individus atteints de la MP, nous retrouvons une dégénérescence accélérée et une dépigmentation des corps cellulaires de la SN avec formation d'inclusions cytoplasmiques hyalines appelées corps de Lewy (JELLINGER, 1986). Les corps cellulaires qui originent de la SN projètent en majeur partie leurs axones vers le ST, formant l'important faisceau dopaminergique nigro-strié. Or, selon CARLSSON, (1959); et BERTLER et ROSENGREN , (1959), le ST contient environ 80% de toute la DA du cerveau. Chez tous les patients atteints de la MP et cela chez tous les types de la maladie, nous retrouvons une diminution importante de la concentration en DA au niveau du ST (BERNHEIMER et coll., 1973). Une corrélation positive a été trouvée entre les changements en DA dans cette région du cerveau et l'importance des pertes cellulaires dans la SN (BERNHEIMER et coll., 1973). D'autres altérations biochimiques associées aux systèmes dopaminergiques ont également été identifiées. Il y a en outre, une diminution des enzymes, tyrosine hydroxylase et dopa décarboxylase (LLOYD et coll ., 1975), de même qu'une diminution des métabolites HVA et DOPAC (BERNHEIMER et coll., 1973). Selon ces mêmes auteurs, aucun symptôme n'est visuellement apparent avec des diminutions en DA de moins de 80% de la concentration normalement contenue dans le ST. En effet, il
18
semble que des mécanismes de compensation permettent d'atténuer les effets de la diminution en DA et cela jusqu'à un seuil qui peut varier entre 70 et 80% . Les mécanismes de compensation pourraient être dus à une augmentation de l'activité des cellules intactes et à la supersensibilité des récepteurs de la DA au ST, cela permettant d'atténuer les effets de baisses aussi marquées du neurotransmetteur. Par contre, lorsque les concentrations en DA sont de moins de 20% des taux normaux, les mécanismes de compensentions ne suffisent plus et les symptômes de la MP apparaissent (BERNHEIMER et coll., 1973). Le ST est composé des noyaux caudé et putamen. Or, selon HORNYKIEWICZ et KISH, (1986), c'est cette dernière partie qui serait la plus affectée et impliquée dans la MP.Il est également très fréquent d'observer une dégénérescence des corps cellulaires noradrénergiques du locus coeruléus de même que de la région du tegmentum ventral (UHL et coll.,1983; JELLINGER et KISH, 1986). Les concentrations en DA et en NE sont variablement réduites dans plusieurs régions du cerveau incluant, le ST, le noyau accumbens et quelques noyaux de !'hypothalamus. L'atteinte de la région du tegmentum ventral permet en outre, d'expliquer les diminutions en DA dans des régions cérébrales comme le noyaux accumbens, les tubercules olfactifs et de quelques noyaux de l'amygdale (FARLEY et coll., 1977). D'autres altérations ont été rapportées dans la littérature mais il n'en demeure pas moins que la perte marquée en DA au niveau du ST constitue l'altération biochimique la plus évidente de la MP (HORNYKIEWICZ et KISH, 1986).
Le traitement des patients atteints de la MP a été directement basé sur les connaissances acquises de la neurochimie de la maladie. Cela constitue en ce sens, le premier exemple "réussi" selon cette approche, dans le traitement d'un désordre neurodégénératif chronique. L'observation que la DA est fortement diminuée dans la MP (BARBEAU, 1960), a conduit au développement de l'utilisation de son précurseur naturel, soit le L-dopa, afin de tenter de rétablir les niveaux en DA chez les patients (BARBEAU et
1 9
coll., 1961 ). La DA elle-même n'est pas efficace puisqu'elle ne traverse pas la barrière hémo-encéphalique (BARBEAU et coll ., 1961; MARX, 1979). Dans le but d'augmenter l'efficacité du précurseur, des inhibiteurs de la L-dopa décarboxylase (benzérazide et carbidopa) peuvent être administrés conjointement avec le L-dopa. Ces inhibiteurs d'enzymes ont l'intéressante propriété de ne pas traverser la barrière hémo-encéphalique, ce qui permet d'éviter la transformation du L-dopa en DA en périphérie et ainsi favoriser l'action du précurseur au cerveau (RIEDERER et coll., 1984). L'utilisaton du déprényl, un inhibiteur de la monoamine oxydase B responsable en majeur partie de la dégradation de la DA dans le cerveau, a aussi permis d'optimiser les actions du L-dopa. En réduisant le catabolisme de la DA, le déprényl permet de diminuer la dose de L-dopa et atténue ainsi ses effets "on and off" chez le patient (BIRKMAYER, 1978). Parmi d'autres types de traitements utilisés mentionnons l'amantadine, qui stimule la libération de la DA à partir des terminaisons (SCHWAB et coll., 1972), et la bromocriptine qui est un agoniste dopaminergique utilisé généralement avec le L-dopa (KELLER et DAPRADA, 1979; LIEBERMAN et GOLSTEIN, 1985). Durant les débuts de la maladie, des agents anticholinergiques tel que le trihéxyphénydil et la benztropine peuvent être utilisés afin de renverser l'hyperactivité cholinergique au ST. Puisque la DA est un inhibiteur de l'activité cholinergique dans le ST, sa diminution se traduit par une augmentation de l'activité de l'acétylcholine dans cette région. Cependant, la diminution des taux en DA progresse constamment au cours de l'évolution de la maladie et devient tellement importante que les capacités de régulariser le rapport DA/acétylcholine par des anticholinergiques s'atténuent ( RIEDERER et coll., 1984 ). Parce qu'il semble que plusieurs neurotransmetteurs sont altérés dans la MP, il est possible que le trairement idéal devra agir sur plus d'un système à la fois (RIEDERER et coll., 1984).Si la symptomatologie et la pathologie de la MP sont bien connues, par contre, l'éthiologie n'est pas encore bien comprise chez 75 à 90% des patients (JELLINGER,
20
1976). Parmi ce groupe, 60 à 75% de_s patients représentent la forme 'idiopathique' (donc de causes inconnues) de la MP et environ 15% proviennent des autres types de neurodégénérescences. Il existe différentes classes à la maladie qui sont basées sur les causes qui ont provoqué l'apparition de celle-ci. Ainsi, 6 à 8% des patients ont des atteintes suite à un désordre cérébrovasculaire , le parkinson post-encéphalopathique est responsable d'environ 3% des cas et un faible pourcentage est associé à des traumatismes crâniens, des effets neurotoxiques de drogues ou à des agents environnementaux. Selon HOEHN et YAHR, (1967); TERÂVÂINEN et coll. (1986), l'appararition de la maladie survient autant chez les hommes que les femmes. Aussi, la majorité des individus ont entre 55 et 58 ans. L'incidence de la maladie serait de 20 :100 000 habitants alors que la population totale atteinte serait de 200:100 000 habitants (RAJPUT et coll., 1984; KURTZKE, 1984). Bien qu'une prédisposition génétique ait été trouvée, principalement chez les patients idiopathiques à début précoce (BARBEAU et ROY, 1984; ELISA ALONSO et coll., 1984), il semble qu'en général, l'histoire familiale n'a que peu d'importance dans l'apparition de la maladie chez un individu (WARD et coll., 1983; TERÂVÂINEN et coll., 1986).
3.1 Les neuropeptides et la maladie de Parkinson
La découverte de nombreux neuropetides dans le système nerveux central (SNC) et les évidences de plus en plus certaines de leur rôle de neuromodulateur ou neurotransmetteur ont conduit à évaluer leur implication possible dans les désordres neurologiques (BEAL et MARTIN, 1986; GERSTENBRABD et POEWE, 1986). Selon BEAL et MARTIN, (1986), l'étude des altérations des neuropeptides dans les maladies neurologiques pourrait aider à mieux comprendre la spécificité des systèmes atteints ou résistants durant la maladie. De plus, cela pourrait contribuer à mieux distinguer les différentes formes de la MP. Bien que peu d'évidences directes impliquant les
21
neuropeptides dans l'étiologie, la symptomatologie ou la pathologie des troubles neurologiques ont été trouvées, il y a par contre assez de preuves montrant qu'ils ont des niveaux anormaux dans les tissus pour justifier les travaux entrepris pour tenter de comprendre leur fonction dynamique et de déterminer s'ils sont directement impliqués dans ces maladies (BEAL et MARTIN, 1986). De plus, puisque la plupart des peptides induisent des effets relativement prolongés, l'intérêt qu'ils suscitent en vue de leur utilisation future en thérapie sërait justifié (MARSDEN, 1986).Puisque l'étiologie de la MP est encore inconnue, l'analyse de l'implication possible des peptides dans cette maladie fut des plus étudiée (BISSETTE et coll., 1985; AG IO et JAVOY-AGID, 1985; BEAL et MARTIN, 1986; GERSTENBRAND et POEWE, 1986; AG IO et coll., 1986). Selon ces auteurs, l'étude de cerveaux de patients atteints de la MP a montré des diminutions en [Met]enk, [Leu]enk, substance P, cholecystokinine-8 , NT, somatostatine et bombésine. Aussi, UHL et coll., (1986) ont trouvé que les récepteurs spécifiques à la NT, la somatostatine et aux opiacés de type kappa, sont très fortement diminués au niveau de la SN de patients atteints de la MP. Leur étude suggère que les neuropeptides pourraient jouer un rôle important comme neuromodulateur de l'activité neuronique des cellules de la SN.
Il fut plutôt inusité que le premier peptide à avoir été expérimenté en clinique contre les désordres extrapyramidaux ait été le tripeptide MIF (Pro-Leu-NH2), qui n'avait pas encore été trouvé dans le cerveau humain. Chez le rat, ce peptide s'est avéré un stimulateur des effets du dopa et un antagoniste des propriétés excitatrices de l'oxotrémorine, qui est un agoniste cholinergique (PLOTN IKOFF, 1975). Il fut donc intéressant d'expérimenter ce peptide contre la MP. Les résultats des essais cliniques ont été relativement satisfaisant et rapportés par différents laboratoires (KASTIN et BARBEAU, 1972 ; FISHER et coll ., 1974;