Intra- and intercrop diversification in cereal cropping and effect on pest control
179
0
0
Texte intégral
Figure

+7


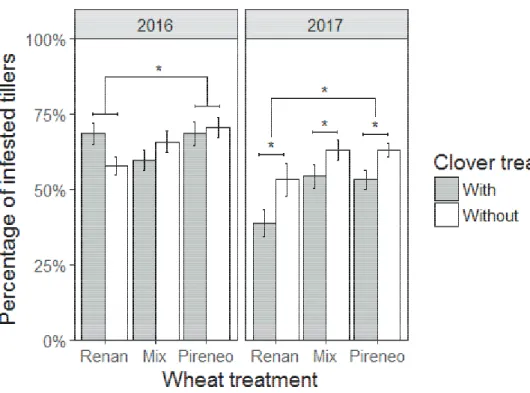

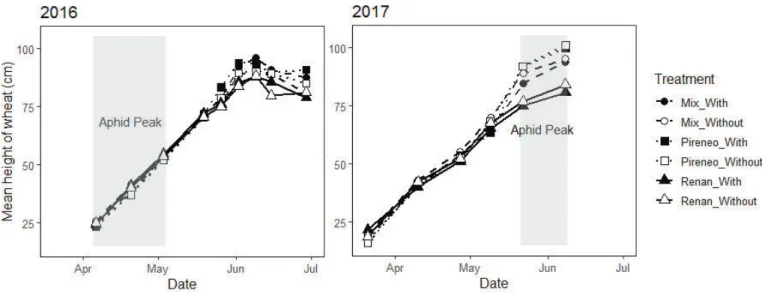
Documents relatifs