Does Retinotopy Influence Cortical Folding in Primate Visual Cortex?
5
0
0
Texte intégral
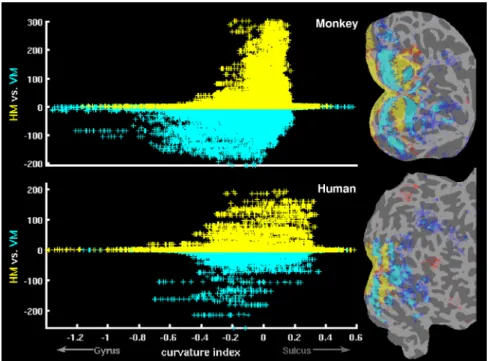
Figure

Documents relatifs