Structure terminale des chromosomes : le « capuchon télomérique »
Texte intégral
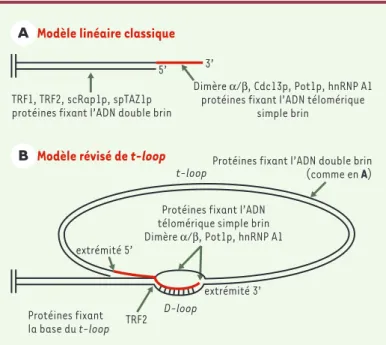
Figure
![Figure 1. Structure primaire des télomères. A. Séquences télomériques des chromosomes de dif- dif-férents organismes [5, 17]](https://thumb-eu.123doks.com/thumbv2/123doknet/12478499.338421/2.892.341.812.523.992/structure-primaire-télomères-séquences-télomériques-chromosomes-férents-organismes.webp)

Documents relatifs
L’ADN ou acide désoxyribonucléique qui compose le patrimoine génétique de tous les organismes vivants est fragile, et les cassures double brin, c’est-à-dire la rupture du
ملو ،ةنٌزخلل ةصاخلا تاباسحلا نمض نم ةٌلاملا نٌناوقب قلعتملا نوناقلا ًف اهركذ .ةٌونسلا ةٌلاملا نٌناوق اهل رشت ةٌمومعلا تاسسؤملا اهردصت ًتلا مهسلأا
A sharp turbidity peak of short duration (ca. 60 – 120 minutes) occurred during flood rise at the Upper site and shortly preceded maximum discharge at the Lower site (data shown for
L’UNIVERSITÉ CLAUDE BERNARD- LYON1 pour l’obtention du DIPLOME DE DOCTORAT soutenue publiquement le 24 septembre 2014 par Lydia FAUQUE TITRE Mécanismes Moléculaires de la
L’analyse chromosomique de cer- taines tumeurs des tissus mous, plus particulièrement des liposarcomes du sous-type histologique bien diffé- rencié ou encore de la tumeur
Criblage par PCR et FISH Microcellule Fusion cellulaire Cellules A9 de souris Fusion de microcellule Cellules ES de souris Cellules ES contenant le chromosome humain Injection à
L’ évolution du pH au cours de la fermentation est comparable à celle observée pour les deux autres milieux : une chute concomitante à la consommation de la source azotée
Cette analyse a été divisée en deux parties, une partie in vitro consistant en un clonage et une expression hétérologue des domaines A des NRPS pour l’analyse de leur activité, et
