Single medium microbial fuel cell: Stainless steel and graphite electrode materials select bacterial communities resulting in opposite electrocatalytic activities
10
0
0
Texte intégral
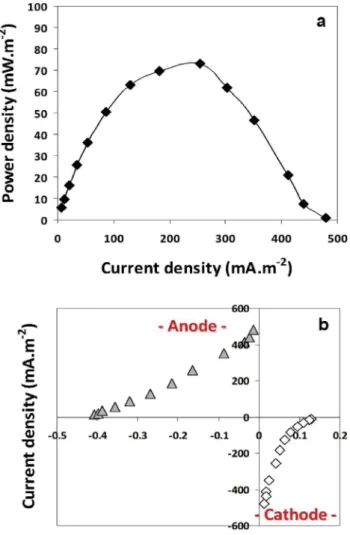
Figure
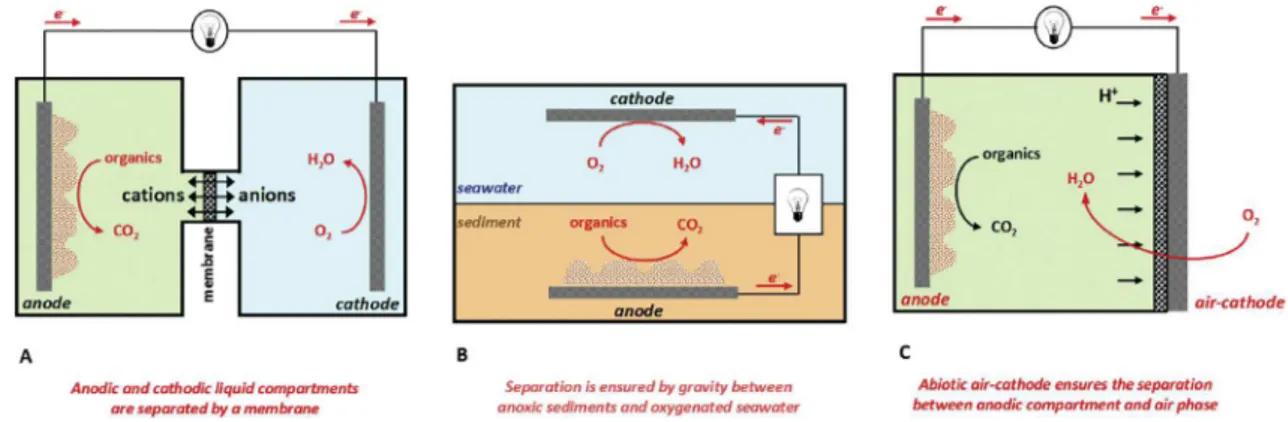
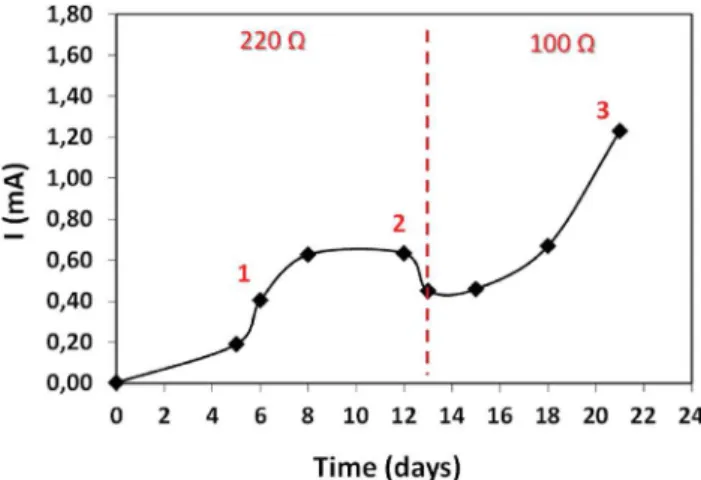
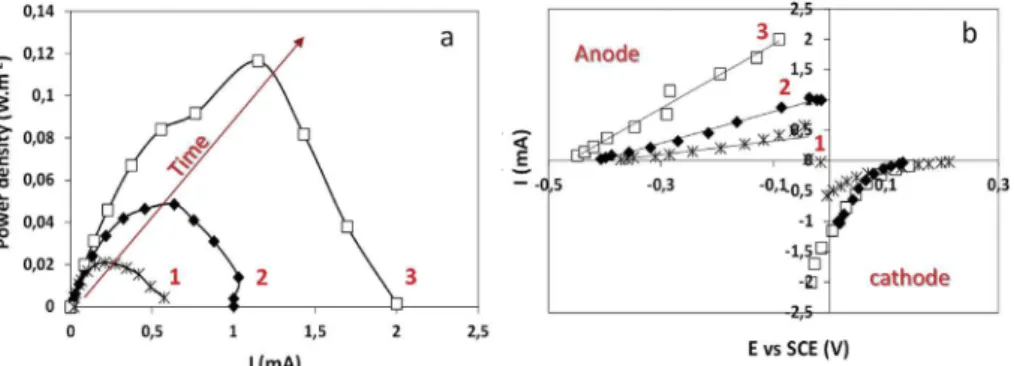
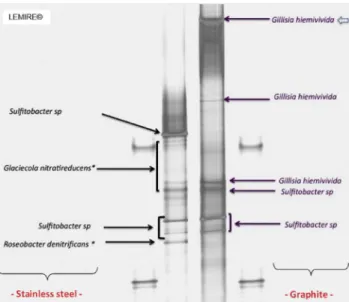
+3

Documents relatifs
