Winning versus losing during gambling and its neural correlates
Texte intégral
Figure
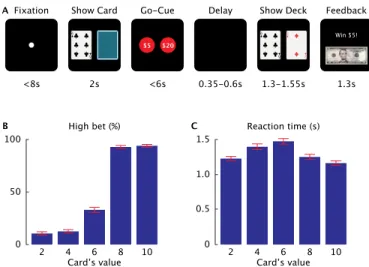

Documents relatifs
The self-reference effectis associated with increased brain activation changes during encoding, suggesting that the beneficial effect of Self on memory may occur at encoding
Participants received individually adjusted electrical pain stimuli in a high-distraction condition (one-back task) and in a low-distraction condition (simple letter response
A brief summary is given of recent results on the use of noise in the optimal training of neural networks for the retrieval of static patterns and on competition between the..
This leads to a new connection between ANN training and kernel methods: in the infinite-width limit, an ANN can be described in the function space directly by the limit of the NTK,
A l’équateur, le déséquilibre local du gradient de pression radial et l’accélération centrifuge cause un écoulement radial allant du voisinage de la sphère
A series of experiments are designed to verify the anxiolytic effect of ethyl acetate extract and laetispicine in the light/dark test, elevated plus maze test and
At intermediate interactions in quench dynamics, we see this same behavior but the critical momentum at which this Cherenkov behavior kicks in requires the
Le corps des pectinaires est allongé en cône inverse , et régulier comme le tube qu’il habite. 11 est extrêmement re marquable par les deux peignes roides à paillettes dorées
