Purification et caractérisation de signaux biologiques à l'état de traces
Texte intégral
Figure
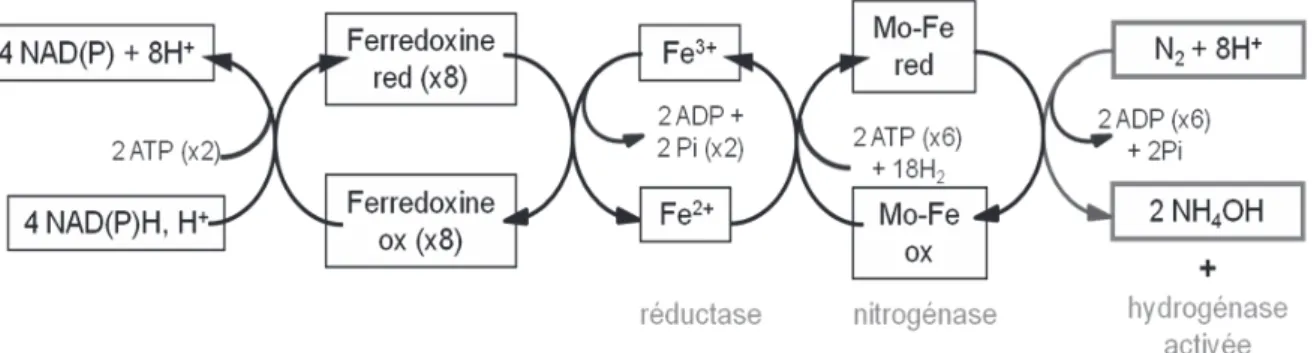


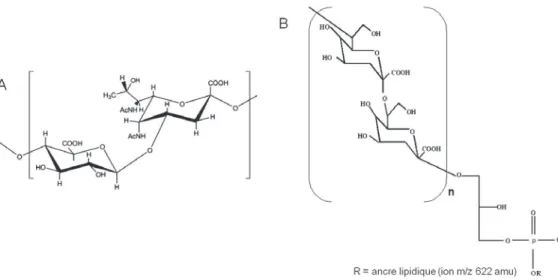
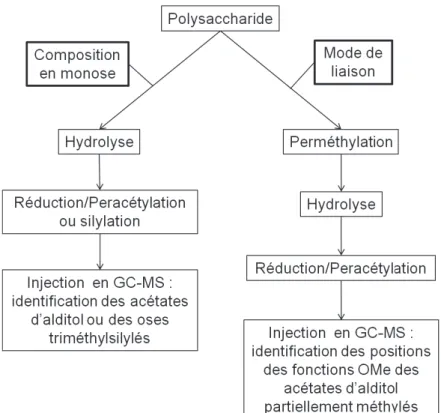


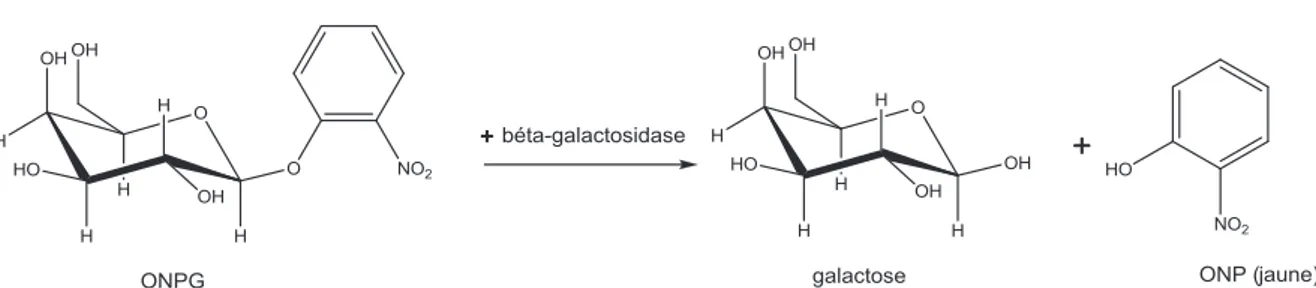


Outline
Documents relatifs
Les contenus digestifs révèlent que les espèces récoltées dans l’herbier de posidonies montrent une préférence pour les algues épiphytes, mais ingèrent aussi
Ces résultats ont été confirmé par les teneurs phénoliques et flavonoïdes de l’extrait méthanolique du M .vulgare qui sont faible par rapport au celles des
Pour tester cette méthode dans le cadre du problème de la classification des signaux de la base SISED, il faut maintenant établir un moyen d’attribuer une classe à un signal
A partir des exemples de notre corpus, le parti pris initial de proposer une description sémantique de ces phénomènes vise à montrer que le locuteur recherche, pour chaque défigement
On voit bien ainsi que les distributions de Pareto (et plus généralement les asymptotiquement paretiennes de P. Levy) sont, dans l’étude des
Le nombre de travaux publiés sur les lectines a vu une grande croissance principalement grâce à l’abondance des lectines dans tous les organismes vivants, accompagné d’une
Avant de démoduler les signaux, nous allons linéariser les détecteurs de puissance, proposer un circuit de compensation des tensions continues qui apparaissent à la sortie
Développement et transfert technologique de procédés chromatographiques destinés à la préparation de composés biologiques à usage thérapeutique : exemples de la purification
