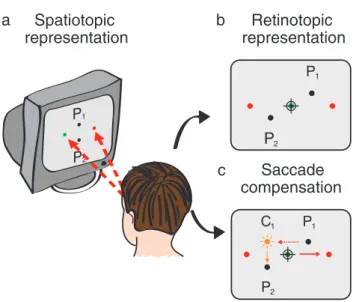
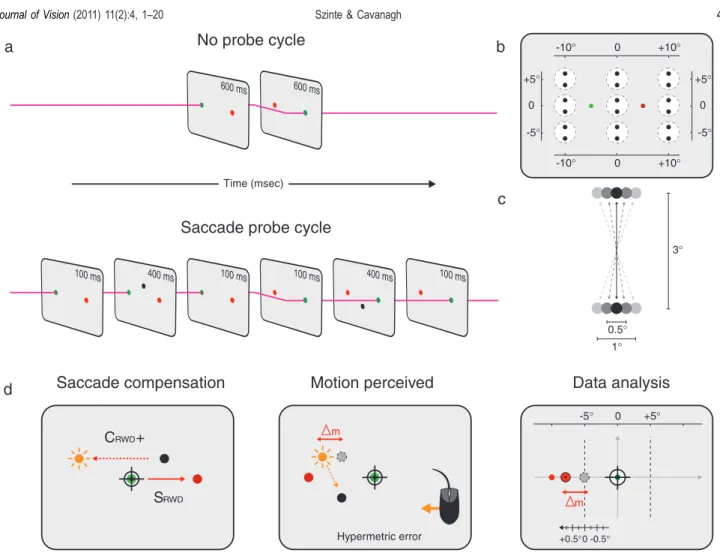
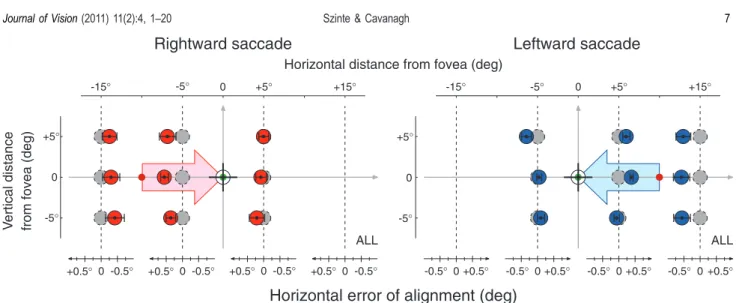
The recovery of target locations in space across movements of eyes and head
194
0
0
Texte intégral
Figure


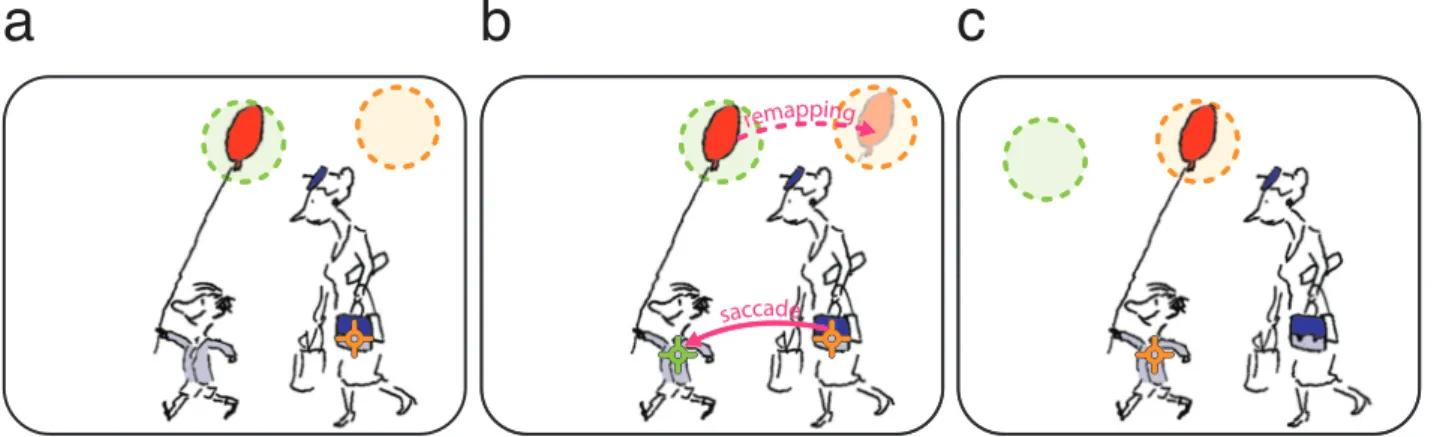

+7
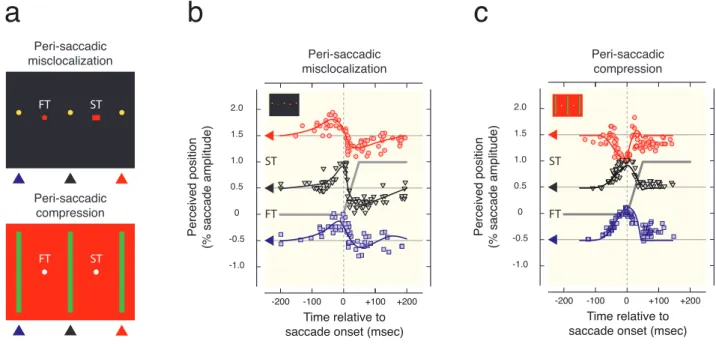
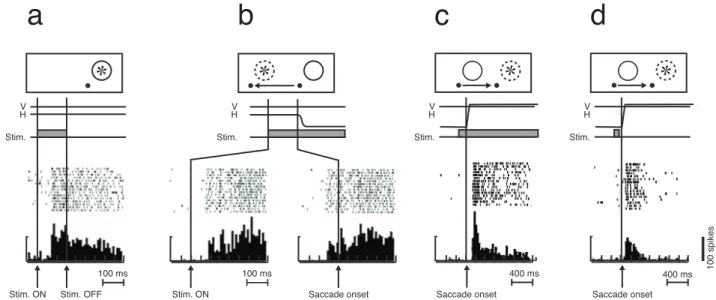
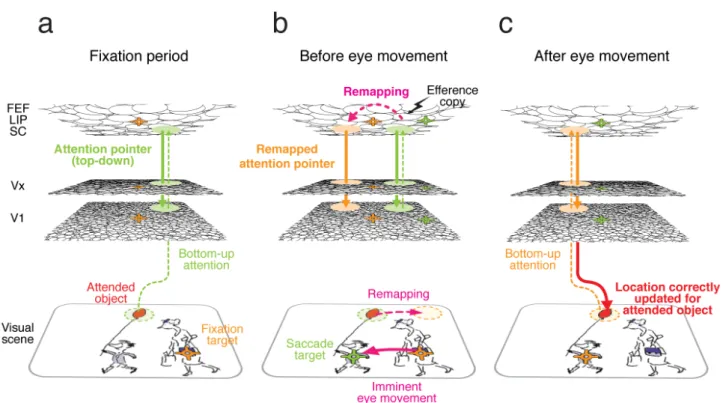
Documents relatifs