Interactions des ions métalliques avec des formes tronquées du peptide amyloïde-bêta liées à la maladie d'Alzheimer
214
0
0
Texte intégral
Figure
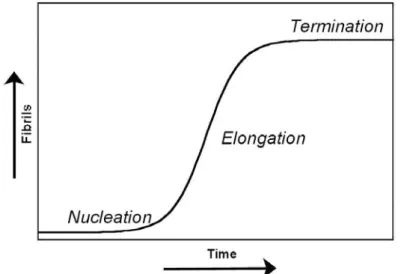
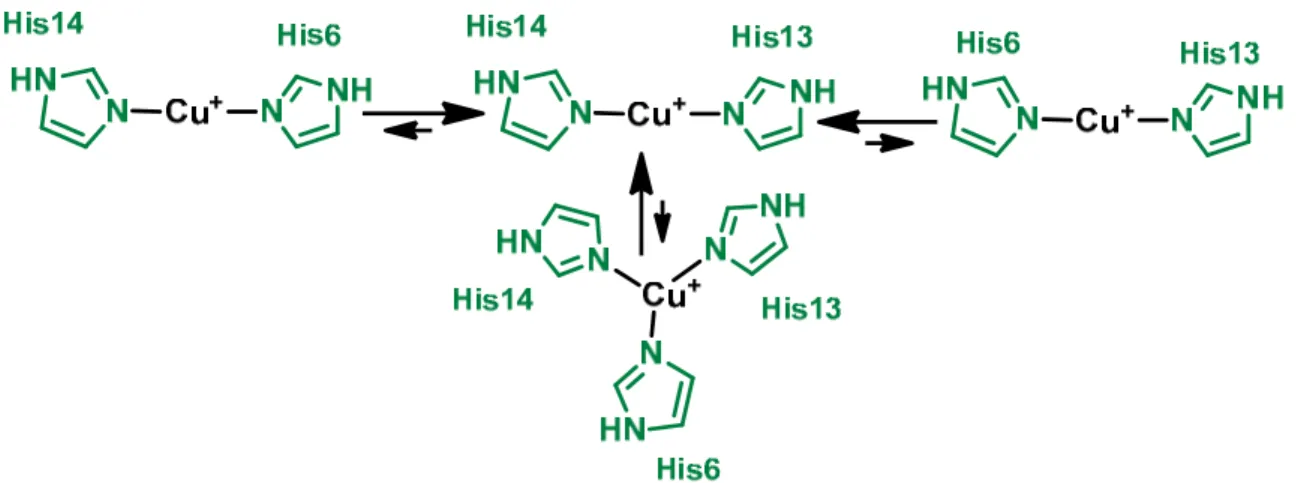

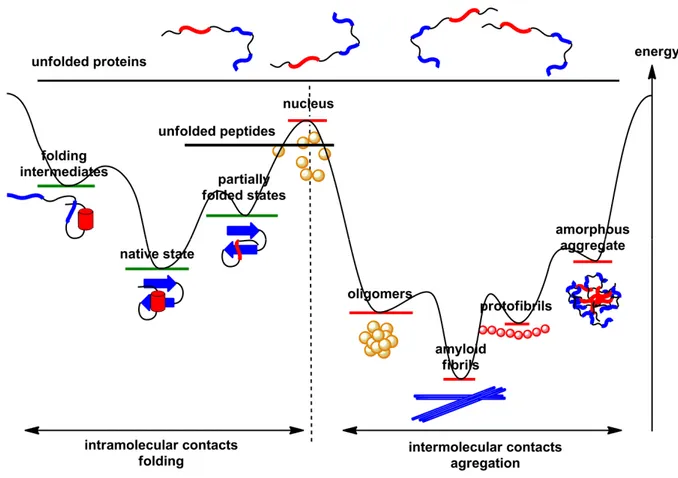
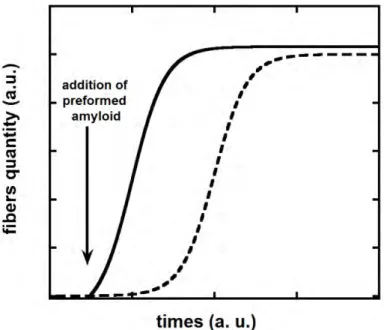
+7
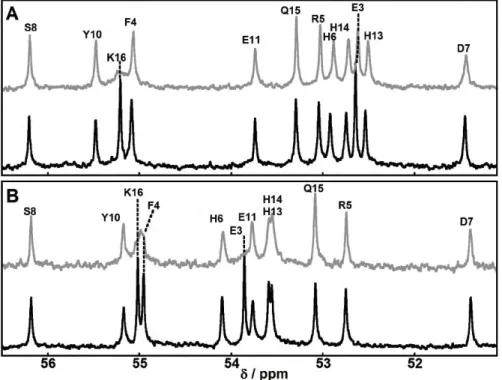
![Figure 2. Effect of pH on the CD spectra of [Cu II (peptide)] complexes, from pH approximately 3 (blue line) to pH approximately 12 (red line)](https://thumb-eu.123doks.com/thumbv2/123doknet/2088764.7347/70.911.200.715.95.643/figure-effect-spectra-peptide-complexes-approximately-approximately-line.webp)
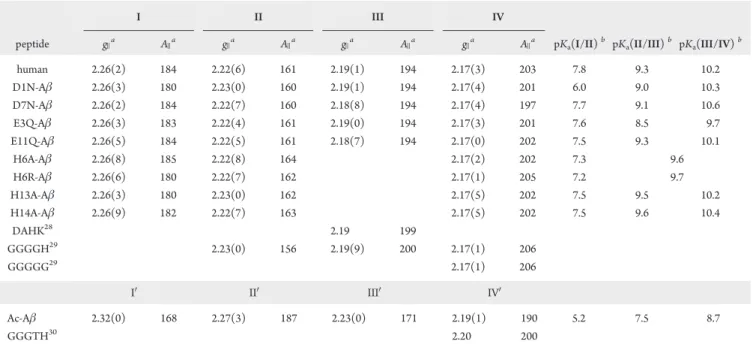
![Figure 3 in the case of the [Cu II (Aβ16)] complex, this could also](https://thumb-eu.123doks.com/thumbv2/123doknet/2088764.7347/71.911.473.826.106.609/figure-case-cu-ii-av-complex.webp)

Documents relatifs