Paléoécologie du Quaternaire
dans la région de Wainwright en Alberta
Mémoire
Charles Clavet
Maîtrise en sciences géographiques
Maître en sciences géographiques (M.Sc.Géogr.)
Québec, Canada
Paléoécologie du Quaternaire
dans la région de Wainwright en Alberta
Mémoire
Charles Clavet
Sous la direction de :
Martin Lavoie, directeur de recherche
Michel Parent, codirecteur de recherche
iii
Résumé
En 2012, la Commission géologique du Canada a procédé à une série de forages dans les dépôts quaternaires de la région de Wainwright dans le centre-est de l’Alberta dans le cadre d'un projet en hydrogéologie. Les forages, qui atteignent plus de 60 m de profondeur, montrent la présence de niveaux organiques intercalés dans les unités stratigraphiques minérales, dont certains contiennent des morceaux de bois qui suggèrent la présence passée in situ d'une végétation ligneuse. La position stratigraphique des sables contenant des niveaux organiques sous le till régional (Wisconsinien supérieur) laisse penser qu'ils correspondraient à des épisodes interstadiaires, voire même de l'interglaciaire Sangamonien, des datations 14C ayant livré des âges non-finis
(>43 500 ans A.A.). L’objectif principal de la présente étude était de documenter la composition du couvert végétal associé aux niveaux organiques à l’aide de l’analyse pollinique et macrofossile. Plusieurs des niveaux se sont avérés pauvres ou même stériles en grains de pollen et en macrorestes végétaux. Les assemblages polliniques sont le plus souvent dominés par des espèces herbacées (p. ex.: Cyperaceae, Artemisia sp.) et des fougères. Chez les arbres, l’épinette est l’espèce la mieux représentée mais elle montre des pourcentages polliniques relativement faibles. Les assemblages macrofossiles sont notamment caractérisés par la présence de matière ligneuse, de l’épinette et de plantes aquatiques. Plusieurs échantillons contiennent aussi des charbons de bois qui témoignent de l’incidence de feux. Les résultats des analyses paléoécologiques suggèrent que le couvert végétal contemporain du dépôt des niveaux organiques était à caractère boréal ouvert. Ces niveaux auraient donc été mis en place au cours d’un intervalle climatique plus froid que l’actuel, présumément un interstade wisconsinien plutôt qu’au cours du dernier interglaciaire (Sangamonien).
iv
Table des matières
Résumé ... iii
Liste des tableaux ... v
Liste des figures ... vi
Remerciements ... vii
1. Introduction ... 1
1.1 Objectif et hypothèse ... 6
2. Région d’étude ... 8
3. Méthodologie ... 10
3.1 Travaux sur le terrain... 10
3.2 Travaux en laboratoire ... 11 4. Résultats ... 14 4.1 Analyse pollinique ... 17 4.2 Analyses macrofossiles ... 21 5. Discussion ... 25 6. Conclusion ... 29 7. Références ... 30 Annexe 1 ... 33 Annexe 2 ... 34 Annexe 3 ... 36
v
Liste des tableaux
Tableau 1 : Datations 14C des forages Catt-28, E1-30 et F2-27. ... 16 Tableau 2 : Niveaux ayant fait l’objet d’une analyse pollinique dans les forages F2-27 et C3-31. 17 Tableau 3: Niveaux ayant fait l'objet d'une analyse macrofossile dans les forages F2-27, C3-31 et E1-30. ... 21
vi
Liste des figures
Figure 1: Localisation de quelques exemples de sites où des dépôts sédimentaires datant de l’interglaciaire Sangamonien ont été trouvés au Canada et aux États-Unis: 1) East Bay, Castle Bay et Lower Middle River, Nouvelle-Écosse; 2) East Milford, Nouvelle-Écosse; 3) Hillsborough, Nouveau-Brunswick; 4) Trois-Rivières, Québec; 5) Toronto, Ontario; 6) Innerkip, Ontario; 7) Moose River Crossing, Ontario; 8) Central Illinois; 9) Bird, Manitoba; 10) Minnedosa, Manitoba; 11) Fort Qu’Appelle, Saskatchewan; 12) Saskatoon, Saskatchewan; 13) Montana plains,
Montana; 14) Medicine Hat, Alberta; 15) Bindloss, Alberta; 16) Watino, Alberta; 17) Westwold, Britannique; 18) Quesnel Forks, Britannique; 19) Quesnel, Colombie-Britannique; 20) Rat River, Territoires du Ouest; 21) Lower Carp Lake, Territoires du Nord-Ouest; 22) Old Crow River Locality 44, Yukon; 23) Herschel Island, Yukon; 24-25) Banks Island, Territoires du Nord-Ouest; 26) Boothia Peninsula, Nunavut; 27) Clyde Foreland, île de Baffin, Nunavut. Carte réalisée à partir de St-Onge (1987), Harrington (1990) et Fréchette & de Vernal
(2013). ... 2
Figure 2: Sites où des restes de vertébrés datant du Wisconsinien moyen et du début de l'Holocène ont été trouvés en Alberta. Carte réalisée à partir de Burns (1996). ... 3
Figure 3: Localisation de la base militaire "The Western Area Training Centre" en Alberta (figure réalisée à partir d’une carte de la Commission géologique du Canada). ... 8
Figure 4: Localisation des forages (étoiles rouges) réalisés par la Commission géologique du Canada en 2012 dans la région de Wainwright. Le forage C3-GW-26 n’apparait pas sur la carte car le trépan s’est brisé lors des travaux d’échantillonnage et les sédiments sous-jacents au till ne purent donc être récoltés. (Figure réalisée à partir d’une carte de la Commission géologique du Canada). ... 10
Figure 5: Coupes stratigraphiques des différents forages faits à Wainwright, Alberta. ... 14
Figure 6: Granulométrie du forage F2-27. ... 16
Figure 7: Spectres polliniques des quatre échantillons du forage F2-27. ... 19
Figure 8: Dénombrement des pièces macrofossiles extraites de quatre échantillons du forage F2-27. ... 22
vii
Remerciements
Un remerciement spécial à tous ceux et celles qui ont collaboré à la réalisation de mon mémoire de maîtrise : Michel Parent (chercheur à la Commission géologique du Canada, division de Québec) qui fut l’instigateur de ce mémoire, Martin Lavoie (directeur, professeur au Département de géographie et chercheur au Centre d’études nordiques de l’Université Laval), Najat Bhiry (membre du comité, professeure au Département de géographie et directrice et chercheure au Centre d’études nordiques), Myosotis Bourgon Desroches et Élisabeth Robert (professionnelles de recherche au Centre d’études nordiques), Daniel Landry (archéologue, Université de Montréal) et Lyna Lapointe Elmrabti (étudiante à la maîtrise au Département de géographie de l’Université de Montréal).
1
1. Introduction
Le Quaternaire, qui représente les 2,7 derniers millions d’années, est divisé en deux grandes périodes : le Pléistocène (2,7 Ma – 11 700 ans A.A. [avant aujourd’hui]) et l’Holocène (˂11 700 ans A.A.). Le Pléistocène fut caractérisé par des périodes glaciaires entrecoupées d’interglaciaires. La dernière glaciation, celle du Wisconsinien, s’est amorcée il y a environ 80 000 ans suite à l’interglaciaire Sangamonien (130 000 - 80 000 ans A.A.; stade isotopique 5) et a atteint son apogée il y a environ 20 000 ans (Dyke & Prest, 1987). La glaciation wisconsinienne est divisée en trois principales périodes : le Wisconsinien inférieur (80 000 - 65 000 ans A.A.; stade isotopique 4), le Wisconsinien moyen (65 000 - 23 000 ans A.A.; stade isotopique 3) et le Wisconsinien supérieur (23 000 - 11 700 ans A.A.; stade isotopique 2). Pendant le Wisconsinien, des périodes de recul des glaciers ont eu lieu (interstades). Les conditions climatiques pendant certains de ces interstades furent assez clémentes pour permettre une reprise de la dynamique végétale, tel fut le cas lors du Wisconsinien moyen. Comparativement à un interglaciaire, les conditions climatiques d’un interstade sont plus froides et la végétation est de type plus boréale. Les conditions plus chaudes d’un interglaciaire permettent, dans les régions tempérées, le développement d’un couvert végétal similaire à l’actuel et la présence d’espèces plus thermophiles (Clet & Occhietti, 1995).
Des dépôts sédimentaires d’âges sangamonien et interstadiaire ont été trouvés à plusieurs endroits au Canada, autant dans les régions tempérées que dans les régions boréales et nordiques (Figures 1 et 2). Sur le plan stratigraphique, ils sont généralement situés entre des couches de tills. Ces dépôts consistent, selon le cas, en des graviers, des sables, des silts, des argiles d’origine fluviale ou lacustre et parfois mêmes des niveaux de matière organique pouvant contenir divers types de fossiles végétaux ou animaux.
2
Figure 1: Localisation de quelques exemples de sites où des dépôts sédimentaires datant de l’interglaciaire Sangamonien ont été trouvés au Canada et aux États-Unis: 1) East Bay, Castle Bay et Lower Middle River, Nouvelle-Écosse; 2) East Milford, Nouvelle-Nouvelle-Écosse; 3) Hillsborough, Nouveau-Brunswick; 4) Trois-Rivières, Québec; 5) Toronto, Ontario; 6) Innerkip, Ontario; 7) Moose River Crossing, Ontario; 8) Central Illinois; 9) Bird, Manitoba; 10) Minnedosa, Manitoba; 11) Fort Qu’Appelle, Saskatchewan; 12) Saskatoon, Saskatchewan; 13) Montana plains, Montana; 14) Medicine Hat, Alberta; 15) Bindloss, Alberta; 16) Watino, Alberta; 17) Westwold, Colombie-Britannique; 18) Quesnel Forks, Colombie-Britannique; 19) Quesnel, Colombie-Britannique; 20) Rat River, Territoires du Nord-Ouest; 21) Lower Carp Lake, Territoires du Nord-Nord-Ouest; 22) Old Crow River Locality 44, Yukon; 23) Herschel Island, Yukon; 24-25) Banks Island, Territoires du Nord-Ouest; 26) Boothia Peninsula, Nunavut; 27) Clyde Foreland, île de Baffin, Nunavut. Carte réalisée à partir de St-Onge (1987), Harrington (1990) et Fréchette & de Vernal (2013).
Dans l’ouest canadien, des fossiles de vertébrés, datant possiblement du Sangamonien, ont été découverts dans la région de Medicine Hat en Alberta (Figure 1, site 14) par Stalker et Churcher (1982) dans une unité stratigraphique nommée « Unit XIII » qui se trouve entre deux couches de till. Des datations U/Th d’ossements ont livré des âges de 74 000 ± 5000 et 76 000 ± 5000 ans A.A. Ces résultats suggèrent que les fossiles en question correspondent à la fin du Sangamonien ou au début du Wisconsinien.
Il fut aussi possible de reconstituer les conditions environnementales qui prédominaient au Sangamonien à l’aide de restes de vertébrés. Par exemple, sur 30 espèces de mammifères identifiées en Saskatchewan et en Alberta, environ 40 % d’entre-elles sont des espèces habituellement associées à des environnements de prairies ouvertes, 17 % à des
3
forêts-parcs, 14 % à des environnements boisés, 7 % à des forêts conifériennes ou à la toundra forestière, alors que les autres (22 %) sont plutôt caractéristiques des prairies alpines (Harington, 1978). Ces régions étaient donc, au Sangamonien, fort probablement caractérisées en partie par de vastes étendues de prairies ouvertes sans doute similaires à celles qui prévalent aujourd’hui. Des fossiles de chiens de prairies (Cynomys niobrarius
churcherii) découverts à Hand Hills au nord-est de Calgary (Figure 2) ont, pour leur part,
été datés entre 33 000 et 22 000 ans. Cette espèce suggère des conditions relativement sèches et de faibles écarts saisonniers de température (Young et al., 1999).
Figure 2: Sites où des restes de vertébrés datant du Wisconsinien moyen et du début de l'Holocène ont été trouvés en Alberta. Carte réalisée à partir de Burns (1996).
4
Le site « January Cave » dans le sud-ouest de l’Alberta au piedmont des montagnes Rocheuses (Figure 2) est un lieu important en ce qui concerne nos connaissances des conditions environnementales durant la seconde moitié du Wisconsinien moyen (33 000 - 23 000 ans 14C A.A.) dans l’Ouest canadien (McAndrews, 1984). La composition du
couvert végétal a été déduite de l’analyse pollinique de 190 cm de sédiments calcareux contenant un peu de matière organique. Les sédiments contenaient très peu de grains de pollen arboréens et arbustifs (épinette, pin, sapin, bouleau, saule) et une représentation pollinique élevée de certaines plantes herbacées (Caryophyllaceae, Compositeae, Poaceae, Cyperaceae). Les restes d’animaux trouvés sont ceux de petits mammifères comme le pika et la marmotte (espèces d’environnements alpins), l’écureuil fouisseur colombien et le rat des bois (espèces associées aux prairies subalpines), le campagnol à queue ainsi que la souris sylvestre (espèces associées aux milieux subalpins). En résumé, ces éléments suggèrent pour cette époque un climat plutôt froid et sec, et un couvert végétal sans doute très ouvert et similaire à celui d’une toundra arbustive (Burns, 1991). Ces interprétations vont de pair avec celles déduites pour la région de Hand Hills décrites précédemment.
Une autre découverte datant du Wisconsinien moyen en Alberta concerne une couche de matière organique de 80 centimètres d’épaisseur dont la composition est similaire à de la tourbe dans la région de Birch Mountains, au nord-ouest de Fort McMurray (Paulen et al., 2005; Figure 2). Cette couche se situe entre deux unités de till et comporte des macrorestes végétaux comme du bois et des aiguilles de pin (Pinus sp.), de l’écorce, de la mousse, des charbons de bois et des graines de cypéracées. Deux datations effectuées à partir de matière organique en vrac ont livré des âges de 32 690 ± 340 (bois de pin) et ˃50 000 ans 14C A.A. À ce jour, ce site est celui le plus au nord-est de l’Alberta qui
montre une séquence datant du Wisconsinien moyen. Des macrorestes d’épinette (Picea sp.) datant de 35 000 à 43 000 ans 14C A.A. ont aussi été découverts dans la région
d’Edmonton (Figure 2; Burns, 1996).
Une étude récente a été réalisée en Saskatchewan sur des paléosols dunaires carbonisés (Bélanger et al., 2014). Les datations de charbons de bois dans les paléosols ont livré des âges variant entre 26 000 et 50 000 ans A.A. L’identification botanique des charbons a montré la présence locale du pin, du sapin (Abies sp.), de l’épinette, du mélèze
5
(Larix sp.) et du genévrier (Juniperus sp.). Ces résultats suggèrent que des arbres étaient présents dans un contexte périglaciaire, dans le centre du Canada, à cette époque. Ceci expliquerait pourquoi, selon les auteurs de l’étude, que le processus d’afforestation postglaciaire dans cette région se serait produit relativement rapidement et ce, sans épisode préalable de toundra, car les arbres devaient talonner le glacier en retrait. En contrepartie, il est surprenant de constater que les paléosols carbonisés ne se trouvent pas sous des dépôts glaciaires; il est ainsi difficile d’affirmer avec certitude qu’ils datent bel et bien du Wisconsinien moyen. Les dates obtenues sont peut-être dues à la contamination de charbons très anciens par des acides humiques ou fulviques provenant de la pédogénèse holocène. Si tel est le cas, il serait plus probable que ces charbons soient d’âge holocène plutôt que Wisconsinien moyen.
Plusieurs études ont aussi été menées ailleurs au Canada concernant les conditions environnementales (végétation, climat) du Sangamonien et des interstades du Wisconsinien moyen à l’aide de l’analyse pollinique et macrofossile. Plusieurs d’entre-elles suggèrent que le climat du Sangamonien était relativement similaire, voire même plus chaud, qu’aujourd’hui (Klassen, 1989; Schweger, 1989; Anderson et al., 1990; Mott, 1990; Clet & Occhietti, 1995; Fréchette et al., 2008; Fréchette & de Vernal, 2013). Par exemple, au Cap-Breton, la température moyenne annuelle aurait été de 6 à 7°C plus élevée qu’aujourd’hui et l’ensoleillement supérieur de 15 à 20 % (Fréchette & de Vernal, 2013). Au Wisconsinien moyen, entre 50 000 et 40 000 ans 14C BP dans la région des Grands-Lacs, il y aurait eu
formation d’une forêt coniférienne de type « boréal » composée entre autres de pins et d’épinettes (Bajc et al., 2015). En région atlantique, il y aurait eu durant le Wisconsinien moyen au moins trois épisodes de forêt boréale en alternance avec des épisodes de végétation de toundra (Fréchette & de Vernal, 2013).
Les études réalisées à ce jour en Alberta concernant les dépôts interglaciaires et interstadiaires se sont attardées davantage à la stratigraphie des dépôts sédimentaires et aux restes de vertébrés plutôt qu’à la composition du couvert végétal, sans doute en raison du fait que les dépôts étudiés étaient peu propices à la préservation des grains de pollen et des macrorestes végétaux. L’état de préservation des grains de pollen, des spores et des pièces macrofossiles dépend notamment du type de sédiments et des conditions de saturation en
6
eau du milieu. L’analyse pollinique permet de dresser un aperçu du couvert végétal contemporain du dépôt du sédiment analysé, le plus souvent à une échelle spatiale régionale (Lavoie, 2001). Pour sa part, l’analyse macrofossile livre des informations sur la composition in situ du cortège floristique car les pièces macrofossiles ne sont en général pas dispersées sur de grandes distances et se déposent près des plantes émettrices (Bhiry & Filion, 2001). Ces pièces peuvent être du bois, des charbons de bois, des graines, des feuilles, etc. L’analyse pollinique et l’analyse macrofossile sont deux techniques paléoécologiques complémentaires qui fournissent une image de la composition et de la structure du couvert végétal passé, image qui doit être interprétée. Par exemple, ce sont surtout les espèces anémophiles qui sont bien représentées dans les assemblages polliniques en raison de leur plus grande productivité pollinique et de leur plus grand pouvoir de dispersion.
En 2012, le Ministère de la Défense nationale du Canada a mandaté l’Institut national de la recherche scientifique (INRS) et la Commission géologique du Canda (CGC) pour réaliser une série de forages de plusieurs dizaines de mètres de profondeur dans la région de Wainwright, au sud-est d’Edmonton en Alberta, dans le cadre d’un projet portant sur l’hydrogéologie régionale. Le mandat était de déterminer l’architecture stratigraphique des formations quaternaires sous-jacentes et d’évaluer l’impact potentiel de celle-ci sur les conditions hydrogéologiques sur le territoire de la base militaire “The Western Area
Training Centre” et des secteurs adjacents. Certains forages ont révélé la présence de
niveaux organiques, à priori d’âge interglaciaire ou interstadiaire. Ces forages constituaient ainsi une occasion unique d’augmenter nos connaissances sur la nature du couvert végétal qui prévalait à ces époques dans l’ouest canadien.
1.1 Objectif et hypothèse
L’objectif de ce projet était d’obtenir des informations relatives à la composition du couvert végétal qui prévalait lors de la mise en place des dépôts organiques trouvés dans les forages réalisés dans la région de Wainwright, dépôts à priori d’âge pré-Wisconsinien inférieur, c’est-à-dire interglaciaire (Sangamonien) ou interstadiaire (Wisconsinien moyen
7
ou supérieur). L’hypothèse qui a été testée est que le couvert végétal de l’interglaciaire Sangamonien était similaire à l’actuel (forêt-parc dominée par le peuplier faux-tremble), alors que celui des interstades correspondait à une forêt de type boréal. Afin de répondre à l’objectif du mémoire et de tester cette hypothèse, l’analyse pollinique et l’analyse macrofossile ont été utilisées.
8
2. Région d’étude
La région d’étude se situe à environ 200 km au sud-est d’Edmonton en Alberta. Les forages qui ont été effectués se trouvent sur le territoire de la base militaire fédérale “The
Western Area Training Centre” (WATC), qui couvre une superficie de 614 km2 au
sud-ouest de la ville de Wainwright (Figure 3).
Figure 3: Localisation de la base militaire "The Western Area Training Centre" en Alberta (figure réalisée à partir d’une carte de la Commission géologique du Canada).
Dans cette région, le paysage est légèrement vallonné. Le relief le plus prononcé consiste en la vallée encaissée de la rivière Battle. Les principaux cours d’eau sont la rivière Battle, qui traverse la partie ouest de la région, et le ruisseau Ribstone qui traverse la partie
9
sud-est. La région est caractérisée par la présence d’une vallée préglaciaire orientée est-ouest qui a été enfouie sous 140 mètres de dépôts préglaciaires et glaciaires. Les dépôts de surface comprennent principalement des sédiments glaciaires, des sédiments sablo-graveleux fluvio-glaciaires, des sédiments sableux et silteux glacio-lacustres et du sable d’origine éolienne, dont des dunes (Martel et al., 2013). La température moyenne annuelle oscille entre 0,5 et 2,5 oC. La température moyenne estivale varie de 13 à 16 oC alors
qu’elle se situe entre -14,5 et -12,5 oC en hiver. Les précipitations moyennes annuelles sont
peu élevées, atteignant de 200 à 300 mm selon les endroits (Ricketts et al., 1999; Charlier et al., 2009).
Le couvert végétal de la région comprend plus de 60 000 hectares de forêt-parcs dominées par le peuplier faux-tremble (Populus tremuloides) et, dans une moindre mesure, par le peuplier baumier (Populus balsamifera). Aussi appelée « tremblaie-parc », il s’agit d’une formation végétale relativement ouverte comprenant aussi des peuplements d’épinette blanche (Picea glauca) et de sapin baumier (Abies balsamea). Le pin gris (Pinus
banksiana) s’observe sur les stations les plus sèches, alors que le saule (Salix spp.),
l’épinette noire (Picea mariana) et le mélèze sont présents sur celles moins bien drainées. Les espaces non forestiers sont occupés par des arbustes et des plantes herbacées (p. ex :
Carex spp., Artemisia spp.). La tremblaie-parc est une écorégion qui fait partie du biome
« prairies, savanes et brousses tempérées » (Ecological Stratification Working Group, 1995). Ce biome forme un arc depuis la frontière du Manitoba et du Dakota du Nord aux États-Unis jusqu’au centre de l’Alberta. D’après les diagrammes polliniques de deux sites au sud-est d’Edmonton, la formation végétale de la tremblaie-parc se serait formée il y a environ 3000 ans avant nos jours (Vance et al., 1983). Sur la base de données polliniques à haute résolution temporelle, Campbell et al. (1994) ont suggéré que le peuplier aurait toutefois beaucoup augmenté dans le paysage depuis la fin du 19e siècle suite à l’importante
10
3. Méthodologie
3.1 Travaux sur le terrain
Huit forages ont été réalisés par la Commission géologique du Canada en 2012. Il s’agit des forages G2-GW-25, C3-GW-31, C3-GW-26, F2-GW-27, CATT-GW-28, C1-GW-29, E1-GW-30 et 3A-GW-13 (Figure 4).
Figure 4: Localisation des forages (étoiles rouges) réalisés par la Commission géologique du Canada en 2012 dans la région de Wainwright. Le forage C3-GW-26 n’apparait pas sur la carte car le trépan s’est brisé lors des travaux d’échantillonnage et les sédiments sous-jacents au till ne purent donc être récoltés. (Figure réalisée à partir d’une carte de la Commission géologique du Canada).
Les sites de forage ont été choisis le long d’un profil sismique du sous-sol. La méthode qui a été utilisée pour l’échantillonnage est celle du forage rotasonique. Elle offre l’avantage de forer rapidement et de produire des échantillons en continu. La foreuse
11
fournit des carottes à des intervalles de 152 cm dans des manchons en plastique. Quatre forages (G2-GW-25, C3-GW-31, Catt-GW-28 et F2-GW-27) ont atteint une profondeur de 61 mètres. Les autres ont atteint des profondeurs de 31 m (C1-GW-29), 21 m (C3-GW-26) et 46 m (3A-GW-13 et E1-GW-30). Les échantillons ont été immédiatement observés et décrits sur le terrain par le personnel de la Commission géologique du Canada. Des sous-échantillons ont ensuite été prélevés à des intervalles réguliers de 30, 76 et 122 cm pour chaque intervalle de 152 cm, de même qu’à l’interface entre les unités stratigraphiques. Les sous-échantillons furent ensuite entreposés à la base militaire à Wainwright et à la Commission géologique du Canada à Québec.
3.2 Travaux en laboratoire
Analyses granulométriques
Des analyses granulométriques ont été faites sur des échantillons du forage F2-27 en raison de ses unités stratigraphiques et des données paléoécologiques qu’il était susceptible de fournir puisqu’il s’agissait du forage possédant le plus grand nombre de niveaux organiques. Les analyses ont été réalisées pour 29 niveaux à l’aide d’un granulomètre laser (modèle Horiba LA-950V2) dans les laboratoires de l’INRS à Québec. L’analyse granulométrique permet de déterminer quantitativement la distribution des particules d’un échantillon par classes de diamètres et donc de fournir des renseignements sur le type d’environnement sédimentaire qui prévalait au moment du dépôt de l’échantillon.
Analyses du contenu en matière organique
Des analyses du contenu en matière organique ont été effectuées pour les forages F2-27 (18 échantillons), C3-31 (18 échantillons) et E1-30 (7 échantillons). Les volumes analysés variaient selon la quantité de sédiments disponibles. Ces analyses ont été faites au Laboratoire de géomorphologie et de sédimentologie du Département de géographie de l’Université Laval. La méthode utilisée a été celle de la perte au feu à 550 oC (Heiri et al.,
12 Analyses polliniques et macrofossiles
Les analyses polliniques ont été effectuées pour les forages F2-27 et C3-31 (Figure 5). Elles ont porté sur les niveaux pour lesquels les grains de pollen étaient susceptibles d’être bien conservés (p. ex.: silt, argile). Pour le forage F2-27, 12 échantillons d’un volume sédimentaire de 2 cm3 ont d’abord été traités chimiquement à l’aide du protocole
usuel de Faegri et Iverson (1989) qui comporte notamment le recours à l’acide fluorhydrique (HF) pour éliminer la matière minérale (Annexe 1). Cette méthode a permis de montrer la présence de grains de pollen dans certains niveaux, mais il était par contre impossible de procéder à leur identification et dénombrement car la quantité de débris organiques ayant résisté aux traitements chimiques était encore trop importante pour permettre de bien voir les grains. De ce fait, une deuxième méthode a été utilisée. Celle-ci consiste à séparer les grains de pollen et les spores de la matrice organique et minérale à l’aide d’un liquide dense, soit le polytungstate de sodium. Cette méthode a été développée par M. Daniel Landry au Laboratoire Jacques-Rousseau de l’Université de Montréal (Annexe 2). Ainsi, les mêmes 12 niveaux du forage F2-27 ont été traités à partir cette fois d’un volume sédimentaire de 3 cm3. En ce qui concerne le forage C3-31, 12 échantillons
ont été traités avec la méthode usuelle. Dans la mesure du possible, une somme pollinique minimale de 300 grains de plantes vasculaires terricoles fut comptée pour chacun des échantillons. Les résultats sont exprimés sous la forme de pourcentages polliniques. La représentation en pourcentage des taxons invasculaires a été calculée à partir de la somme pollinique de base. La concentration pollinique des échantillons (grains/cm3) a été calculée
grâce à l’ajout de pastilles de spores de Lycopodium lors du traitement chimique.
L’analyse macrofossile a été réalisée pour 26, 28 et 30 échantillons prélevés respectivement des forages F2-27, C3-31 et E1-30 (Figure 5). Les niveaux ont été choisis par rapport à leur position stratigraphique et aux endroits où il était le plus plausible de trouver de façon simultanée du pollen et des macrorestes végétaux (dans le cas des forages F2-27 et C3-31). La méthode qui fut employée en laboratoire pour extraire les pièces macrofossiles est présentée à l’Annexe 3 (Bhiry & Filion, 2001). En fonction de la quantité de sédiment disponible, le volume analysé pour chacun des échantillons variait entre 4 et 43 cm3. Les résultats de l’analyse macrofossile du forage F2-27 sont présentés sous la
13
forme de dénombrements (nombre de pièces par volume sédimentaire analysé). Pour les forages C3-31, E1-30 et quelques niveaux du F2-27, l’analyse macrofossile ne fut réalisée que de façon qualitative en raison du mauvais état de préservation des pièces.
Datations radiochronologiques
Trois échantillons des forages Catt-28, E1-30 et F2-27 (un par forage; Figure 5) ont fait l’objet de datations 14C par spectrométrie de masse par accélérateur (SMA) au
laboratoire Beta Analytic (Floride). Les dates ont été obtenues à partir de macrorestes végétaux. Pour ce qui est du forage Catt-28, il s’agissait d’un morceau de bois prélevé à la profondeur 47,4 mètres. Pour le forage E1-30, la datation a été faite sur un morceau de bois prélevé à la profondeur 30,9 mètres. Enfin, pour ce qui est de forage F2-27, la datation a été faite sur de la matière végétale (non déterminée) à 28 mètres de profondeur.
14
4. Résultats
Stratigraphie des forages et analyses granulométriques
Dans les forages, on trouve des niveaux constitués de till, de silt, d’argile, de sable et de matière organique (Figure 5). Plus précisément, il y a du sable éolien, du sable alluvial, du sable et silt lacustre et de l’argile silteuse lacustre. Il y a aussi présence de sédiments de plaines d’épandage et des sédiments glaciolacustres proximaux et distaux. Certains des niveaux contiennent des fragments de bois de taille macroscopique, essentiellement dans les unités constituées de sable alluvial.
15
Figure 5: (suite).
Les résultats des analyses granulométriques menées sur des échantillons du forage F2-27 sont illustrés à la Figure 6 et représentent le coefficient d’uniformité (Cu). Les particules dont la taille se situe entre 0,06 et 2 mm sont associées à du sable alors que celles plus petites, entre 0,002 et 0,06 mm, à des silts. Sur les 29 échantillons analysés, 19 se situent entre 0,06 et 2 mm et 10 entre 0,002 et 0,06 mm. Les unités sous les couches de tills sont en grande partie des dépôts de sables fins (14 échantillons) et quelques niveaux sont silto-sableux (9 échantillons). Puisque ces dépôts alluviaux sont plutôt fins, ils se sont probablement mis en place dans un cours d’eau dont le courant était relativement calme. Il n’est par contre pas possible de faire ressortir une tendance entre les différents niveaux analysés car l’écart de profondeur entre les échantillons est trop important.
16
Figure 6: Granulométrie du forage F2-27.
Datations radiochronologiques
Les trois datations 14C réalisées sur les forages Catt-28, E1-30 et F2-27 ont toutes
donné des âges non-finis, soit supérieurs à 43 500 ans 14C BP (Tableau 1).
Tableau 1 : Datations 14C des forages Catt-28, E1-30 et F2-27.
Échantillon
(profondeur) de laboratoire Numéro Matériel daté δ
13C/12C Âge
(années 14C BP)
Catt-28 (47,4 m) Beta-375086 Bois -27.2 ‰ > 43 500 BP
E1-30 (30,9 m) Beta-375087 Bois -23.9 ‰ > 43 500 BP
17
4.1 Analyse pollinique
Les analyses polliniques du forage F2-27 ont toutes été effectuées sur l’unité constituée de sable alluvial entre les profondeurs 25 et 61 m, qui est aussi celle pour laquelle des fragments de bois étaient visibles (Figure 6). Dans le cas du forage C3-31, les analyses ont été réalisées dans les unités constituées de sédiments glaciolacustres distaux (8 à 10 m et 24 à 32 m), de sédiments glaciolacustres proximaux (37 à 39 m), de sable et silt lacustre (45 à 53 m) et d’argile silteuse lacustre (53 à 61 m) (Figure 5). Les échantillons se sont avérés pour la plupart très pauvres ou stériles en grains de pollen et en spores. Sur les 12 niveaux analysés du forage F2-27, seulement quatre (profondeur 25 m, 25,9 m, 28 m et 47,5 m) contenaient du pollen et ce, malgré le fait qu’ils provenaient tous de la même unité (Tableau 2; Figure 8) constituée de sable avec de la matière organique. Les grains de pollen étaient fragmentés et très corrodés, de sorte que plusieurs n’ont pu être identifiés, même au niveau du genre ou de la famille. En ce qui concerne le forage C3-31, les 12 échantillons analysés se sont soit avérés complètement stériles, soit qu’ils ne contenaient que quelques grains en quantité insuffisante pour dresser un spectre pollinique statistiquement fiable, peu importe la composition de l’unité stratigraphique (Tableau 2).
Tableau 2 : Niveaux ayant fait l’objet d’une analyse pollinique dans les forages F2-27 et C3-31.
Forage F2-27 Forage C3-31
Profondeur (m) Pollen/spore Profondeur (m) Pollen/spore
25 √ 5,8 Stérile 25,9 √ 32,3 Stérile 28 √ 35,4 Stérile 31,3 Stérile 36,3 Stérile 34,3 Stérile 38,4 Stérile 36,9 Stérile 45,4 Stérile 39,9 Stérile 46,9 Stérile 43 Stérile 48,5 Stérile 47,5 √ 50,6 Stérile 49 Stérile 52,1 Stérile 52,4 Stérile 53 Stérile 60,2 Stérile 55,2 Stérile
18 Forage F2-27
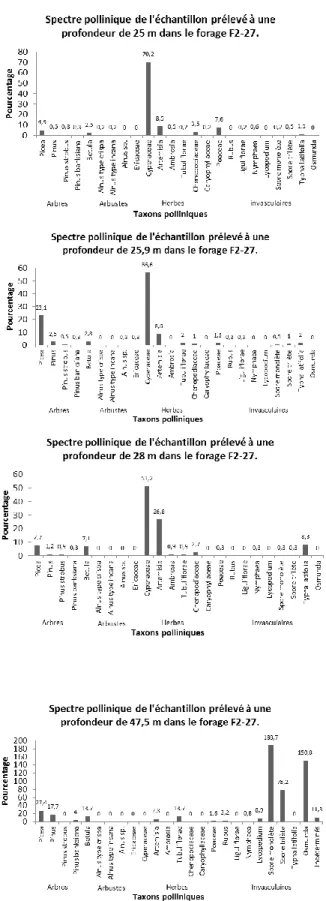
Seuls quatre échantillons du forage F2-27 contenaient du pollen en quantité statistiquement suffisante, dont trois consécutifs dans la partie supérieure de l’unité constituée de sable alluvial (Figure 6). Les sommes polliniques varient entre 124 et 634 grains, le nombre de taxons polliniques de 14 à 18, et les concentrations polliniques de 3150 à 77 830 grains/cm3.
L’échantillon le plus profond à 47,5 mètres de profondeur provient d’un niveau constitué de sable organique (6,2 % de matière organique). La somme pollinique n’est que de 124 grains et la concentration pollinique de 3150 grains/cm3, soit la plus faible des
quatre échantillons (Figure 9). Chez les plantes vasculaires, le spectre pollinique est dominé par le pollen arboréen (63 %). Les taxons polliniques arborescents dominants sont Picea (27 %), Pinus (18 %) et Betula (14 %). Chez les pins, c’est le pin gris (Pinus banksiana) qui a été identifié (4 %). Aucun grain de pollen de plante arbustive n’a été trouvé. La représentation totale des plantes herbacées est de 26 %. Les principaux taxons sont les Tubulifloreae (14 %), Artemisia (7,3 %), Rubus (3,2 %) et les Poaceae (1,6 %). Les spores des plantes invasculaires, notamment les fougères (spores monolètes et trilètes, 267 %), les osmondes (151 %) et les lycopodes (9 %) sont abondantes. La proportion des grains qui n’ont pu être identifiés (grains indéterminés) en raison de leur mauvais état de préservation est aussi très élevée (plus de 11 %).
19
20
L’échantillon à 28 mètres de profondeur est constitué de sable organique (3,0 % de matière organique). La somme pollinique est de 336 grains et la concentration pollinique de 77 832 grains/cm3 (Figure 9). Le spectre pollinique est dominé par les plantes herbacées
(83 %), essentiellement les Cyperaceae (51 %) et Artemisia (27 %). C’est d’ailleurs dans ce niveau que la représentation pollinique d’Artemisia est la plus élevée. Le pollen arboréen atteint 17 %. Les pourcentages polliniques de Picea et de Betula y sont similaires (7,7 et 7,1 % respectivement). Encore une fois, aucun grain de pollen de plante arbustive ne fut trouvé. Une particularité de ce spectre concerne les nombreux grains de pollen de Typha
latifolia (8,3%).
L’échantillon analysé à 25,9 mètres de profondeur est constitué de sable organique (3,4 % de matière organique). La somme pollinique est de 394 grains et la concentration pollinique de 8 360 grains/cm3 (Figure 9). La représentation totale des taxons polliniques
herbacés atteint 70 %. Ils sont représentés essentiellement par les Cyperaceae (57 %),
Artemisia (8,6 %), les Tubulifloreae (2 %), les Poaceae (1,5 %) et les Chenopodiaceae
(1 %). La représentation des arbres est plus élevée que dans l’échantillon précédent, atteignant 29 % : il s’agit surtout de Picea (23 %), Pinus (3,3 %) et Betula (2,8 %).
L’échantillon à 25 mètres de profondeur, soit au sommet de l’unité, est constitué de sable organique (3,9 % de matière organique). La somme pollinique est de 634 grains et la concentration pollinique de 62 000 grains/cm3 (Figure 9). Les herbacées composent plus de
90 % de l’assemblage pollinique, notamment en raison de l’abondance des Cyperaceae (70 %). Les autres principaux taxons polliniques herbacés sont, par ordre d’importance,
Artemisia (8,5 %), les Poaceae (7,6 %) et les Chenopodiaceae (3,5 %). Le pollen arboréen
(9 %) se compose principalement de Picea (4,9 %), Betula (2,5 %) et Pinus (1,6%). Comme dans les trois autres échantillons, pratiquement aucun pollen de plante arbustive ne fut trouvé.
21
4.2 Analyses macrofossiles
Des analyses macrofossiles ont été réalisées pour les forages F2-27 (26 niveaux), C3-31 (28 niveaux) et E1-30 (30 niveaux) (Tableau 3). Dans le cas des forages F2-27 et C3-31, les niveaux analysés pour leur contenu en macrorestes provenaient des mêmes unités stratigraphiques que celles analysées pour le pollen (Figures 5 et 6). En ce qui concerne le forage E1-30, les unités analysées sont composées de sédiments glaciolacustres et de sable alluvial (Figure 6). Plusieurs échantillons se sont avérés stériles. Au total, 12 niveaux du forage F2-27, 16 niveaux du forage C3-31 et 12 niveaux forage E1-30 contenaient des pièces macrofossiles (Tableau 3). Certains niveaux contenaient des pièces macrofossiles comme des morceaux de bois, des coquillages (surtout des gastéropodes), des fragments de matières végétales (feuilles, racines, rhizomes, etc.) et des restes d’insectes. Par contre, dans la plupart des cas, la petite taille et l’état de décomposition très avancé des pièces n’a pas permis de les identifier.
Tableau 3: Niveaux ayant fait l'objet d'une analyse macrofossile dans les forages F2-27, C3-31 et E1-30.
Forage F2-27 Forage C3-31 Forage E1-30
Profondeur (m) Quantité (cm3) Macrorestes Profondeur (m) Quantité (cm3) Macrorestes Profondeur (m) Quantité (cm3) Macrorestes 25 24 √ 5,8 25 √ 18,6 30 Stérile 25,3 20 √ 6,9 11 Stérile 20,1 30 Stérile 25,9 15 √ 7,9 4 Stérile 23 30 Stérile
26,2 14 Stérile 9,8 12 Stérile 23,6 20 Stérile
27,1 22 √ 27,1 12 Stérile 24,5 16 Stérile
27,6 14 Stérile 28,7 4 Stérile 26 16 Stérile
28 8 √ 30,2 24 Stérile 27 42 Stérile
31,2 20 Stérile 31,7 4 Stérile 28,5 42 Stérile
34,3 17 Stérile 32,8 36 Stérile 30,2 30 Stérile
36,9 4 Stérile 33,5 26 √ 30,8 30 Stérile 37,8 11 Stérile 34,7 42 √ 30,9 56 √ 38,1 10 √ 35,4 22 √ 31,1 42 √ 38,4 18 √ 35,8 34 √ 33,2 41 Stérile 39,9 10 Stérile 36,3 31 √ 36 20 Stérile 41 12 √ 36,9 26 √ 37,6 14 √ 41,5 23 Stérile 37,8 30 √ 38,2 30 √ 43 14 Stérile 38,4 25 √ 38,6 20 Stérile 46,7 16 Stérile 45,4 30 √ 39 16 Stérile 46,9 10 Stérile 46,9 23 √ 39,5 29 √ 47,5 20 √ 48,5 20 √ 40,1 42 √ 49 13 Stérile 50,6 26 √ 40,5 1.5 √ 52,1 10 Stérile 51,5 4 Stérile 40,7 20 √ 52,4 20 √ 52,1 43 √ 40,8 30 √ 53,6 16 √ 52,6 30 √ 41,3 32 √ 57,6 20 √ 53 29 √ 41,5 38 Stérile
60,2 10 Stérile 53,6 27 Stérile 41,6 Inconnu √
54,1 26 Stérile 41,7 25 √
55,2 36 Stérile 42,1 32 Stérile
43,6 22 Stérile
22 Forage F2-27
Les quatre échantillons du forage F2-27 qui présentaient du pollen en quantité suffisante pour établir un spectre pollinique contenaient aussi des pièces macrofossiles (Figure 10). Dans le bas du forage, à 57,6 mètres, il y a de gros charbons, des aiguilles de conifères et des fragments de bois. Dans les niveaux à 27,1, 52,4 et 53,6 mètres de profondeur, les macrorestes étaient beaucoup trop dégradés pour permettre leur identification et leur dénombrement.À 47,5 mètres, on trouve beaucoup de petits fragments de charbons et des fragments de mousses brunes. À 41 mètres, on trouve de la matière ligneuse, des charbons, des sclérotes et des fragments de mousses brunes. Dans le niveau sus-jacent, à 38,4 mètres, il y a encore plusieurs gros fragments ligneux, des charbons de bois, des aiguilles, des feuilles de mousses brunes, des coquillages et des sclérotes de
Cenococcum. L’échantillon à 38,1 mètres contient très peu de sable et beaucoup de matériel
végétal. On y trouve exclusivement des restes ligneux, dont un fragment de bois d’épinette.
23
L’échantillon analysé à une profondeur de 28 mètres (8 cm3) contient du sable et un
peu de matériel végétal (3,0 %). Un total de 101 macrorestes a été extrait et ce, malgré le petit volume analysé (Figure 10). Ceux qui ont pu être identifiés au niveau de l’espèce sont pour la plupart des plantes aquatiques ou de milieux humides (Scirpus acutus, Potamogeton sp., Carex sp., Pycnanthemum virginianum). L’échantillon contient également des éphippiums, des akènes entiers et fragmentés de cypéracées (Schoenoplectus pungens,
Carex sp.), quelques petits fragments de cônes et des coquillages.
L’échantillon analysé à la profondeur de 25,9 mètres (15 cm3) contient du sable et
un peu de matière organique (3,4 %). Un total de 63 macrorestes ont été dénombrés (Figure 10). Les taxons macrofossiles sont Carex sp., Pycnanthemum virginianum et
Potamogeton sp. Il y a aussi 31 fragments de graines qui ne purent être identifiés.
L’échantillon contient également des coquillages et des fragments de bois.
L’échantillon analysé à la profondeur de 25,3 mètres (20 cm3) contient du sable et
un peu de matière organique (0,9 %). Le contenu en macrorestes est pauvre, seules six graines de plantes aquatiques (Scirpus acutus, Potamogeton sp.) et une graine de Carex sp. ayant été trouvées (Figure 10). L’échantillon contient également des coquillages et des éphippiums de Daphnia (bryozoaire d’eau douce).
L’échantillon analysé à 25 mètres (24 cm3) contient très peu de sable et beaucoup de
matière organique. Au total, 145 pièces ont été dénombrées (Figure 10). La majorité d’entre elles consistent en des graines de plantes aquatiques (Scirpus acutus, Potamogeton sp.,
Pycnanthemum virginianum). Quatre fragments d’aiguilles de conifères et des graines de Carex sp. ont aussi été trouvés. Vingt fragments de graines ne purent être identifiés en
raison de leur état de dégradation trop avancé. Enfin, l’échantillon contient des éphippiums de Daphnia, des opercules de sphaignes et des coquillages.
24 Forage C3-31
Dans ce forage, il y a présence de macrorestes à 5,8 mètres et de 33,5 à 53 mètres de profondeur (Tableau 3). Cette séquence est constituée en majeure partie par des sédiments glaciolacustres, du sable, silt et argile lacustre. On trouve dans la plupart des niveaux de très petits fragments de matière végétale et ligneuse et de petites particules de charbons de bois. Toutefois, la taille et le degré élevé de décomposition ont rendu les identifications impossibles.
Forage E1-30
Dans le forage E1-30, des macrorestes ont été trouvés dans quelques échantillons plus profonds que 30,9 mètres, soit dans l’unité constituée de sable alluvial (Tableau 3; Figure 6). Dans l’échantillon le plus haut (30,9 m), plus de 95 % des pièces végétales consistent en des fragments ligneux. Les autres pièces sont des charbons, des feuilles de mousses brunes, des graines de Potamogeton, un fragment de graine de Zannichellia
palustris, des coquillages, un fragment d’aiguille de Picea et des sclérotes de Cenococcum.
Aux profondeurs de 37,6, 38,2 et 39,5 mètres, on trouve des fragments de bois, du charbon et des sclérotes. L’échantillon à 40,5 mètres contient peu de sable et la majorité des pièces végétales trouvées sont des fragments ligneux (dont un fragment de Picea sp.) et des charbons. Les pièces végétales sont des fragments d’aiguilles d’épinette. Dans l’échantillon sous-jacent (40,7 m) la majorité des pièces consiste aussi en des fragments ligneux et des charbons de bois. D’autres pièces furent identifiées comme une graine de cf Luzula, des fragments d’aiguilles et un rameau de Picea sp. Un gros fragment d’épinette a également été extrait de cet échantillon ainsi que plus bas à 41,6 mètres. Des fragments de bois ont été extraits à 40,8 et 41,3 mètres, mais n’ont pas été identifiés botaniquement. Du charbon est aussi présent dans l’échantillon à 41,3 mètres. C’est à 41,7 mètres qu’on a trouvé les macrorestes les plus profonds du forage. Cet échantillon contient autant de sable que de matériel végétal et est constitué à 100 % de fragments ligneux. On y trouve aussi de petites particules de charbon et de gros fragments de Picea sp.
25
5. Discussion
Les résultats des trois datations 14C sur les forages Catt-28, E1-30 et F2-27 ont tous
donné des âges non-finis, soit supérieur à 43 500 ans 14C A.A. Les datations ont été faites
sur du bois ou de la matière organique préservée dans des sables alluviaux fins, tous sous-jacents à un till. Cela laisse supposer que ces niveaux organiques sont corrélatifs d’interstades du Wisconsinien, ou encore du Sangamonien.
Avec les données paléoécologiques récoltées dans l’unité de sable alluvial du forage F2-27, il est possible d’obtenir une image partielle de la composition du couvert végétal qui prévalait à certaines périodes du Wisconsinien moyen. Dans le bas du forage (de 38 à 60 m de profondeur), la présence de charbons de bois, d’aiguilles de conifère (épinette) et de fragments ligneux indique que les conditions environnementales étaient suffisamment clémentes pour permettre la présence d’un couvert forestier coniférien, sans doute assez ouvert. Le spectre pollinique du seul échantillon n’étant pas stérile (47,5 m) suggère aussi une forêt de type boréal comportant l’épinette (probablement l’épinette noire) et peut-être aussi le pin (probablement le pin gris) et le bouleau. Dans le cas du bouleau, il pouvait s’agir soit du bouleau à papier (Betula papyrifera), soit du bouleau glanduleux (Betula
glandulosa). L’abondance de spores et de grains de pollen de plantes invasculaires
(fougères, osmondes) laissent croire à des conditions locales humides. Bien que la concentration pollinique soit un outil permettant d’estimer la densité du couvert végétal, la très faible concentration pollinique de cet échantillon (3150 grains/cm3) traduit sans doute
aussi les mauvaises conditions de préservation du pollen dans le sable alluvial (Bajc et al., 2015).
Les conditions semblent s’être modifiées dans la portion supérieure de l’unité de sable alluvial, entre les profondeurs 25 et 38 mètres. Les seuls échantillons analysés contenant du pollen en quantité suffisante sont ceux entre 25 et 28 mètres. À 28 mètres, les données polliniques permettent de supposer un couvert végétal très ouvert, le spectre pollinique étant nettement dominé par les herbacées, principalement les Cyperaceae et
Artemisia sp. Les pourcentages de l’épinette et du bouleau sont faibles (environ 7 %
26
macrofossiles indiquent la présence d’au moins d’arbustes, de sorte que le couvert végétal devait ressembler à celui d’une toundra arbustive. Le dénombrement de plusieurs graines de plantes aquatiques et d’éphippiums de Daphnia indique la présence d’eau localement. Dans le niveau sus-jacent à 25,9 mètres de profondeur, les herbacées sont toujours dominantes. Par contre, le pollen arboréen, plus précisément celui de Picea sp., est plus élevé. Cette augmentation de la représentation pollinique de l’épinette est probablement liée au caractère de plus en plus ouvert du couvert végétal qui permettait l’enregistrement d’apports polliniques lointains de l’épinette, bien que la présence de fragments de bois dans les assemblages macrofossiles démontre qu’il y avait toujours des plantes ligneuses dans le secteur immédiat du lieu de forage. Dans l’échantillon analysé à 25 mètres de profondeur et immédiatement sous le till (Figure 6), le spectre pollinique est dominé par les herbacées (90 %), essentiellement des Cyperaceae. L’assemblage macrofossile comporte pour sa part quelques fragments de conifères et beaucoup de graines de plantes aquatiques (principalement Scirpus acutus). Les données polliniques et macrofossiles de la partie supérieure de l’unité indiquent fort probablement encore une fois l’existence d’une végétation similaire à celle d’une toundra arbustive, peut-être encore plus ouverte qu’antérieurement. La comparaison des données paléoécologiques entre le niveau le plus profond (47,5 m) et ceux plus haut (passage d’une forêt boréale ouverte vers un couvert végétal de type toundra arbustive) suggère une détérioration des conditions environnementales, fort probablement un refroidissement, ce qui est conforme avec la présence du till sus-jacent à l’unité de sable alluvial (Figure 6).
Les analyses macrofossiles du forage E1-30 ont été menées dans les unités constituées de sable alluvial (31 à 42 m) et de sédiments glaciolacustres proximaux (18 à 31 m), mais seuls les sables alluviaux contenaient des pièces macrofossiles (Figure 6). La composition des assemblages livrent une image similaire à celle déduite des données du forage F2-27. Le bas du forage est nettement dominé par des fragments de matière ligneuse. La présence de charbons de bois, de sclérotes de Cenococcum et de pièces macrofossiles d’épinette indiquent que les conditions environnementales étaient suffisamment clémentes pour permettre la présence d’un couvert arborescent sans doute assez ouvert mais comportant suffisamment de combustible pour permettre le déclenchement de feux (Higuera et al., 2011). Étant donné la très mauvaise préservation des grains de pollen dans
27
ce forage, il devient impossible d’interpréter plus précisément la composition et la structure du couvert végétal qui prévalait durant le Wisconsinien moyen. Il en est de même pour le forage C3-31 : les restes ligneux entre les profondeurs 33,5 à 53 mètres témoignent de la présence locale d’arbustes ou d’arbres pendant la période de temps représentée par l’accumulation des unités stratigraphiques correspondantes, mais il est difficile de s’avancer plus loin dans les interprétations.
Les données obtenues suggèrent que les conditions de préservation des grains de pollen, des spores et des pièces macrofossiles varient selon le type de dépôt. Les mauvaises conditions de préservation sont probablement dues au fait que la plupart des dépôts sont constitués de silts et de sable ayant un très faible contenu en matière organique. Ce genre de dépôt laisse des vides entre les sédiments qui ont sans doute favorisé l’oxydation, contrairement aux sédiments plus organiques au sein desquels les conditions anaérobiques ralentissent souvent le processus de décomposition. Les conditions de préservation exercent une grande influence sur les interprétations qui peuvent être tirées des données paléoécologiques provenant de dépôts aussi anciens (Bajc et al., 2015). Les niveaux dont la quantité et la qualité de préservation des témoins biotiques s’avèrent les meilleures sont ceux pour lesquels des conditions saturées en eau ont été identifiées et dont le contenu en matière organique était le plus élevé. De telles conditions s’avèrent propices à la préservation et c’est sans doute pourquoi, dans chaque niveau des actuels forages où ont été trouvés des pièces macrofossiles et du pollen en quantité suffisante, il y avait aussi présence de grains de pollen et spores de plantes invasculaires traduisant des conditions locales relativement humides (p. ex. : fougères). Outre l’oxydation qui est un processus important menant à la dégradation du pollen (Lebreton et al., 2010), les attaques microbiologiques par les bactéries et les champignons et, fort probablement dans le cas présent, les processus mécaniques (ex.: transport du pollen), peuvent aussi mener à la corrosion (Campbell, 1999). Il est aussi connu que le pollen de certains taxons soit davantage susceptible à la corrosion. Par exemple, le pollen d’espèces anémophiles comme le bouleau et certains grains dont l’exine est plus épaisse comme ceux de l’épinette et du pin peuvent être surreprésentés dans les assemblages polliniques en raison non seulement du fait qu’ils s’agissent de forts producteurs, mais aussi de leur plus grande résistance (Birks et Birks, 1980). En contrepartie, le pollen du peuplier se dégrade rapidement et se préserve mal (Mott, 1978).
28
Les présentes données paléoécologiques suggèrent que durant le dépôt des unités analysées, le climat était plus froid qu’aujourd’hui. La région de Wainwright est présentement caractérisée par une forêt-parc dominée par le peuplier faux-tremble. Aucun grain de pollen ni macroreste de cette espèce n’ont été trouvés dans les échantillons. Il n’est pas possible d’affirmer avec certitude que le peuplier n’était pas présent régionalement, compte tenu du fait que ses grains de pollen se préservent mal dans les sédiments et qu’il est très fortement sous-représenté dans les assemblages polliniques (Mott, 1978; Comtois et Payette, 1984; Campbell, 1999). En contrepartie, la composition des assemblages polliniques modernes (analogues modernes) de la forêt-parc au sud-est d’Edmonton, bien que très variable d’un endroit à l’autre, diffère de celle des spectres polliniques issus des forages en termes de diversité et de représentation de plusieurs espèces arborescentes et herbacées (Whitmore et al., 2005). En revanche, bien que la représentation pollinique de l’épinette soit relativement peu élevée dans la pluie pollinique actuelle (de 2 à 18 % selon les endroits), les assemblages macrofossiles suggèrent qu’il s’agissait de l’espèce arborescente prédominante. Quoi qu’il en soit, le couvert végétal reconstitué était différent de l’actuel et était à caractère boréal ouvert.
Les résultats des datations 14C ne nous permettent pas non plus d’affirmer avec
certitude que les dépôts analysés sont d’âge interstadiaire, mais leur position stratigraphique le suggère. En supposant que les échantillons proviennent du Wisconsinien moyen, les présents résultats confirment l’hypothèse qu’à cette époque, le couvert végétal était à caractère boréal et différent d’une forêt-parc. Le climat était donc aussi plus froid qu’aujourd’hui. Ces interprétations vont de pair avec celles issues d’autres recherches paléoécologiques en Alberta. Les résultats de McAndrews (1984), datant du Wisconsinien moyen, montrent aussi un faible taux de grains de pollen arboréens et arbustifs et des pourcentages élevés des herbacées. Nos résultats corroborent également ceux obtenus par Young et al. (1999), Burns (1991; 1996) et Paulen (2005) qui suggèrent tous la présence d’une toundra arbustive au Wisconsinien moyen en Alberta. De plus, le faible taux de préservation des grains de pollen et des pièces macrofossiles est une caractéristique commune à d’autres études menées sur des dépôts d’âge interstadiaire (p. ex. : Bajc et al., 2015).
29
6. Conclusion
Les données recueillies des analyses polliniques et macrofossiles ne permettent pas de donner une image parfaite de la végétation qui prévalait lors des interstades précoces du Wisconsinien moyen, mais elles indiquent que la végétation s’est détériorée devant la progression de l’inlandsis Laurentidien. Il est également possible d’affirmer que ce n’est pas seulement la toundra qui a colonisé la région de Wainwright, mais aussi la forêt boréale. Si les dépôts analysés proviennent bel et bien du Wisconsinien moyen, nos résultats confirment notre hypothèse que durant les interstades, le climat était plus froid que l’actuel et qu’une forêt coniférienne de type boréal a laissé place à un couvert végétal de plus en plus ouvert, et même à une toundra arbustive. Ces résultats correspondent aussi avec les ceux obtenus dans d’autres études faites en Alberta. De plus, la faible abondance du pollen et des pièces macrofossiles dans les forages indique que les conditions de préservation ont un rôle très important à jouer lorsque vient le temps d’interpréter la nature du couvert végétal (Bajc et al., 2015). Certains grains de pollen résistent mieux à la corrosion et sont donc par le fait même mieux représentés dans les assemblages polliniques.
Dans cette recherche, il aurait été intéressant de trouver des niveaux organiques intercalés entre deux dépôts de till. Cela aurait permis d’analyser l’évolution complète de la végétation, soit du recul jusqu’à la réavancée du glacier. De cette façon, il aurait été possible de mieux comprendre l’interaction entre la végétation et le glacier durant les interstades. En somme, les données paléoécologiques de cette étude indiquent que les dépôts analysés sont associés à des conditions interstadiaires. Pour leur part, les données radiochronologiques suggèrent le début du Wisconsinien moyen, voire même la fin du Sangamonien. D’éventuelles études pourraient être faites dans le secteur pour donner plus de précision aux données et à l’interprétation des résultats de cette recherche. Cela permettrait de démystifier si les échantillons sont associés au Wisconsinien moyen.
30
7. Références
Anderson, T.W. Matthews, J.V., Mott, R.J., Richard, S.H., Richard, P.J. H., Vincent, J.S. (1990). The Sangamonien Pointe-Fortune Site, Ontario-Québec Border. Géographie physique et Quaternaire 44 : 271-287.
Bajc, A.F., Karrow, P.F., Yansa, C.H., Curry, B.B., Nekola, J.C., Seymour, K.L., Mackie, G.L. (2015). Geology and paleoecology of a Middle Wisconsin fossil occurrence in Zorra Township, Southwestern Ontario, Canada. Canadian Journal of Earth Sciences 52: 386-404.
Bélanger, N., Carcaillet, C., Padbury, G.A., Harvey‐schafer, A.N., Van Rees, K.J.C. (2014). Periglacial fires and trees in a continental setting of Central Canada, Upper Pleistocene. Geobiology 12: 109-118.
Bhiry, N., Filion, L. (2001). Analyse des macrorestes végétaux. Dans: Payette, S., Rochefort, L. (éditeurs). Écologie des tourbières du Québec-Labrador. Les Presses de l'Université Laval, Québec, pp. 259-273.
Birks, H.J.B., Birks, H.H. (1980). Quaternary Palaeoecology. University Park Press, Baltimore, Maryland, USA.
Burns, J.A. (1991). Mid-Wisconsinan vertebrates and their environment from January Cave, Alberta, Canada. Quaternary Research 35: 130-143.
Burns, J.A. (1996). Vertebrate palaeontology and the alleged ice-free corridor: the meat of the matter. Quaternary International 32: 107-112.
Charlier, J., Charlier-Vanderschraege, D., De Konick, R., Dorval, G. (2009). Le Grand Atlas du Canada et du Monde, Troisième Édition. Groupe De Boeck. 201p.
Campbell, C., Campbell, I.D., Blyth, C.B., McAndrews, J.H. (1994). Bison extirpation may have caused aspen expansion in western Canada. Ecography 17: 360-362.
Campbell, I.D. (1999). Quaternary pollen taphonomy: examples of differential redeposition and differential preservation. Palaeogeography, Palaeoclimatology, Palaeoecology 149: 245-256.
Clet, M., Occhietti, S. (1995). Palynologie des sédiments de la fin de l’optimum climatique de l’interglaciaire sangamonien, île aux Coudres, estuaire du Saint-Laurent, Québec. Géographie physique et Quaternaire 49 : 291-304.
Comtois, P., Payette, S. (1984). Représentation pollinique actuelle et subactuelle des peuplieraies boréales au Nouveau-Québec. Géographie physique et Quaternaire 38 : 123-133.
Dyke, A.S., Prest, V.K. (1987). Late Wisconsinan and Holocene History of Laurentide Ice Sheet. Géographie physique et Quaternaire 41: 237-263.
31
Ecological Stratification Working Group. (1995). A national ecological framework for Canada. Agriculture and Agri-Food Canada, Research Branch, Centre for Land and Biological Ressources Research and Environment Canada, State of the Environment Directorate, Ecozone Analysis Branch, Ottawa/Hull. 125p.
Faegri, K., Iversen, J. (1989). Textbook of Pollen Analysis. 4th edition, revised by K. Faegri, P.E. Kaland and K. Krzywinski. Wiley, Chichester.
Fréchette, B., de Vernal, A. (2013). Evidence for large-amplitude biome and climate change in Atlantic Canada during the last interglacial and mid-Wisconsinan periods. Quaternary Research 79: 242–255.
Fréchette, B., de Vernal, A., Guiot, J., Wolfe, A.P., Miller, G.H., Fredskild, B., Kerwin, M.W., Richard, P.J.H. (2008). Methodological basis for quantitative reconstruction of air temperature and sunshine from pollen assemblages in Arctic Canada and Greenland. Quaternary Science Reviews 27: 1197–1216.
Harrington, C.R. (1978). Quaternary vertebrate faunas of Canada and Alaska and their suggested chronological sequence. Syllogeus 15: 1-105.
Harrington, C.R. (1990). Vertebrates of the Last Interglaciation in Canada: a review, with new data. Géographie physique et Quaternaire 44:375-387.
Heiri, O., Lotter, A.F., Lemcke, G. (2001). Loss on ignition as a method for estimating organic and carbonate component in sediments: reproductibility and comparability of results. Journal of Paleolimnology 25: 101-110.
Higuera, P.E., Chipman, M.L., Barnes, J.L., Urban, M.A., Hu, F.S. (2011). Variability of tundra fire regimes in Arctic Alaska: millennial-scale patterns and ecological implications. Ecological Monographs 21: 3211-3226.
Klassen, R.W. (1989). Le Quaternaire des plaines intérieures méridionales du Canada. Le Quaternaire du Canada et du Groenland. Commission Géologique du Canada. Géologie du Canada 1. pp. 145-184.
Landry, D. (2012). La néolithisation dans la région de Montréal depuis le Sylvicole moyen tardif : apport archéopalynologique. Mémoire de maîtrise Université de Montréal, Montréal, 217 p.
Lavoie, M., 2001.
Lebreton, V., Messager, E., Marquer, L., Renault-Miskovsky, J. (2010). A neotaphonomic experiment in pollen oxidation and its implications for archeopalynology. Review of Palaeobotany and Palynology 162: 29-38.
McAndrews, J. H. (1984). January Cave: Palynology. In “Quaternary Palynology, Extant Flora and Quaternary Geology South and West of Calgary”. Sixth International Palynological Conference Calgary, pp. 46-49.
32
Mott, R.J. (1978). Populus in late-Pleistocene pollen spectra. Canadian Journal of Botany 56: 1021-1031.
Mott, R.J. (1990). Sangamonian forest history and climate in Atlantic Canada. Géographie physique et Quaternaire 44: 257-270.
Martel, R., Gabriel, U., Parent, M., Pugin, A., Pullan, S., Lefrançois, G., Varfalvy, V. (2013). Evaluation of surface and ground water quality at the WATC, Wainwright, Alberta. Rapport de recherche R-1399, Institut national de la recherche scientifique – Centre Eau, terre et environnement, Québec, 331 p.
Paulen, R.C., Beaudoin, A.B., Pawlowicz, J.G. (2005). An interstadial site in the Birch Mountains, north-central Alberta. CANQUA Meeting, Program and Abstracts, Winnipeg, Canada, p. A66.
Ricketts, T. H., Dinerstein, E., Olson, D.M., Loucks, C.J., Eichbaum, W., Dellasala, D., Kavanagh, K., Hedao, P., Hurley, P.T., Carney, K.M., Abell, R., Walters, S. (1999). Terrestrial ecoregions of North America: a conservation assessment. Island Press, Washington DC. 485p.
Schweger, C.E. (1989). Paléo-écologie du corridor non glacé de l’Ouest canadien. Le Quaternaire du Canada et du Groenland. Géologie du Canada 1 : 527-535.
Stalker. A.M., Churcher, C.S. (1982). Ice age deposits and animals from the southwestern part of the Great Plains of Canada. Geological Survey of Canada, Miscellaneous Report 31. St-Onge, D.A. (1987). The Sangamonian Stage and the Laurentide Ice Sheet. Géographie physique et Quaternaire 41: 189-198.
Vance, R.E., Emerson, D., Habgood, T., 1983. A mid-Holocene record of vegetative change in central Alberta. Canadian Journal of Earth Sciences 20: 364-376.
Whitmore, J., Gajewski, K., Sawada, M.,Williams, J.W., Shuman, B., Bartlein, P.J., Minckley, T., Viau, A.E., Webb III, T., Shafer, S., Anderson, P., Brubaker, L. (2005). Modern pollen data from North America and Greenland for multiscale paleoenvironmental applications. Quaternary Science Reviews 24: 1828–1848.
Young, R.R., Burns, J.A., Rains, R.B., Schowalter, D.B. (1999). Late Pleistocene glacial geomorphology and environment of the Hand Hills region and southern Alberta, related to Middle Wisconsin fossil prairie dog sites. Canadian Journal of Earth Sciences 36: 1567-1581.
33
Annexe 1
Protocole de traitement chimique modifié de Faegri et Iversen (1989) en vue de l’analyse pollinique des forages C3-31 et F2-27
1- Prélèvement des sous-échantillons (2-3 cm3 selon le cas).
2- Contamination des échantillons par un marqueur (pollen de Lycopodium) : permettre de calculer la concentration pollinique (grains/cm3).
3- Hydroxyde de potassium 10 %: défloculation des sédiments pour l’étape du tamisage. 3- Tamisage à l’eau distillée : élimination des débris végétaux et de la matière minérale. 4- Acide fluorhydrique (HF): élimination de la silice.
5- Acide chlorhydrique: élimination des carbonates. 6- Acide acétique: élimination de l’eau.
7- Acide sulfurique et anhydride acétique (acétolyse): élimination de la cellulose. 8- Rinçage à l’eau distillée.
9- Rinçage à l’eau glycérinée.
10- Coloration au rouge neutre et montage entre lame et lamelle.