Thèse
pour obtenir le grade de Docteur de l’Université de Poitiers Discipline : Sciences et Techniques des Activités Physiques et Sportives
présentée par
Joëlle ROUHANA
Flexibilité de l’intégration multi-sensorielle :
aspects spatiaux et temporels.
Directeur de thèse : Professeur
Yannick BLANDIN
Soutenance le 28 Novembre 2008
Composition du jury
Bernard THON Pr., Université P. Sabatier, Toulouse (Rapporteur). Brice ISABLEU MCF., Université Paris Sud 11.
Jean-Pierre ORLIAGUET Pr., Université P. Mendès France, Grenoble (Rapporteur). Lucette TOUSSAINT MCF., Université de Poitiers.
Yannick BLANDIN Pr., Université de Poitiers.
Centre de Recherches sur la Cognition et l’Apprentissage CNRS UMR 6234 MSHS 99, Avenue du Recteur Pineau, 86000 Poitiers
La patience est amère mais son fruit est doux……… « Jean Jacques Rousseau »
A mon père …
qui écoutait mes peines en fin de journée qui m’encourageait à chaque début de matinée qui pensait à moi à tout moment pendant ces années Jour après jour tes conseils m’ont construit et construit ce fruit que je t’offre aujourd’hui
Remerciements
Mes remerciements les plus profonds à madame Lucette Toussaint (MCF) et mon directeur de thèse Yannick Blandin (Pr.) qui, par leur soutien, m’ont permis de prendre confiance en moi. Je les remercie pour leurs conseils, leurs disponibilités et encouragements. Sans eux, ce travail n’aurait jamais abouti.
Je remercie les professeurs Orliaguet et Thon ainsi que monsieur Isableu (MCF) pour leur participation à l’examen de ce travail de thèse en tant que rapporteurs et membres du jury.
Je tiens à remercier particulièrement M. Caron pour son soutien.
Je remercie l’équipe « Attention et Contrôle », dirigée par le professeur Yannick Blandin pour m’avoir accueillie et mis à ma disposition un environnement propice à l’apprentissage de la recherche.
Je remercie énormément, pour sa gentillesse, son dévouement et sa passion, M. Pascal Martin, ingénieur en mécanique productique à l’IUT de Poitiers, concepteur d’une table motorisée spécialement pour servir les objectifs de ce travail.
Je remercie également M. Gérard Champenois, directeur du LAII à l’ESIP, qui a mis à ma disposition des stagiaires pour développer le programme de motorisation et d’automatisation de cette table. En ce qui concerne la partie informatique, je remercie Lionel Granjon et Elie Tohme qui n’ont jamais décliné aucune de mes demandes, ainsi que Yannick Berteaux, toujours présent au besoin.
Je remercie, pour leur gentillesse et leur patience non mesurable, tous les participants aux expérimentations effectuées dans le cadre de ce travail.
Hady, Toni, Magali, Christian et toute l’équipe de l’école de danse Rock in Galaxy. Je remercie également Monsieur et Madame Rousseau qui, par leur présence, m’encourageaient à garder espoir et à vouloir toujours mieux.
Mes remerciements les plus profonds à ma tante Foutine et à mes cousines qui ont cru en moi, à mon père, ma mère, mon frère et ma sœur qui ont tous survécu et m’ont aidé à survivre à mon éloignement des fois difficile.
Enfin et surtout,
RESUME
Ce travail examine la flexibilité de l’intégration multi-sensorielle dans ses aspects spatiaux et temporels. Dans un premier temps, nous nous intéressons à l’intégration des informations visuelles et proprioceptives dans la localisation d’une position. La performance est étudiée dans une tâche de reproduction passive d’une position spatiale (Exp. 1) en croisant trois modalités d’encodage et de reproduction (visuelle, proprioceptive, visuo-proprioceptive). Dans un second temps, nous nous intéressons à l’intégration des informations visuelles et auditives au cours du temps. La performance est étudiée dans une double tâche de temps de réaction de choix visuo-auditive. Deux tâches visuo-manuelle et auditivo-manuelle et une double tâche visuo- et auditivo-manuelle sont proposées en manipulant l’ordre de présentation des stimuli (Exp. 2), en introduisant une amorce sur l’ordre de présentation des stimuli (Exp. 3) et en recourant à une population d’experts en «Counter strike» (Exp. 4). Les principaux résultats ont montré que la similarité et la richesse bimodale des phases d’encodage et de reproduction sont optimales pour la localisation d’une position (Exp. 1). L’intégration visuo-auditive au cours du temps est influencée par les différences individuelles, en partie par la fiabilité de la modalité sensorielle et l’ordre de présentation des stimuli, et par l’expertise individuelle (Exp. 2, 3, 4). L’ensemble de ce travail met en évidence une flexibilité des processus d’intégration multi-sensorielle en fonction de multiples facteurs dont les uns agissent aussi bien sur l’intégration des informations spatiales que sur l’intégration des informations au cours du temps.
Mots-clés : Intégration multi-sensorielle, dominance, priorité de traitement, vision, proprioception, audition.
ABSTRACT
This work examines the flexibility of multisensory integration in its spatial and temporal aspects. First, we are interested in the integration of visual and proprioceptive information in localizing a position. Performance is studied in a spatial position reproduction task (Exp. 1) crossing three conditions of encoding and reproduction (visual, proprioceptive, visuo-proprioceptive). Second, we are interested in the integration of visual and auditory information during time. Performance is studied in a visuo-auditory dual task of choice reaction time. Two visuo-manual and auditory-manual tasks and one visuo and auditory-auditory-manual dual task were proposed handling the stimuli presentation order (Exp. 2), introducing a cue about the stimuli presentation order (Exp. 3) and resorting to a population of experts in «Counter strike» (Exp. 4). Main results showed that the similarity and the bimodal richness of both encoding and reproduction phases are optimal for localizing a position (Exp. 1). Visuo-auditory integration during time is influenced by individual differences, partly by the reliability of sensory modality and the stimuli presentation order, and by expertise (Exp. 2, 3, 4). This work revealed flexibility in multisensory integration processes according to multiple factors of which some act as well on the integration of spatial information and on the integration of information during time.
Keywords: Multisensory integration, dominance, processing priority, vision, proprioception, audition.
Remerciements... i
Résumé...iii
INTRODUCTION ... 1
PARTIE I - CONTEXTE THEORIQUE ... 3
Introduction : Flexibilité de l’intégration multi-sensorielle. ... 4
Chapitre 1 : Intégration des informations spatiales visuelles et proprioceptives... 6
I. Introduction. ... 6
II. Modèles d’intégration visuo-proprioceptive ... 7
II.1. Flexibilité en fonction de la fiabilité des modalités sensorielles... 7
II.2. Flexibilité en fonction de l’étape du traitement perceptivo-moteur... 10
II.3. Flexibilité en fonction de l’efficacité des transferts intermodaux... 11
III. Questions méthodologiques relatives à l’étude de l’intégration visuo-proprioceptive. ... 14
III.1. Précision de la modalité visuelle (V) et de la modalité proprioceptive (P). ... 14
III.2. Le transfert d’information entre la vision et la proprioception... 14
III.3. Précision en situation bimodale VP. ... 15
Chapitre 2 : Intégration des informations visuelles et auditives au cours du temps. 20 I. Introduction. ... 20
II. Attention et double tâche de temps de réaction de choix... 21
II.1. Différence entre tâches de TR simple (TRS) et TR de choix (TRC). ... 21
II.2. L’attention : ses aspects et fonctions... 22
III. Modèles d’interférence et supports expérimentaux. ... 24
III.1. Les modèles d’interférence... 24
III.2. Le paradigme expérimental. ... 26
III.3. Les principaux résultats expérimentaux : évidence d’un goulot d’étranglement central et structural. ... 29
stimuli visuel et auditif. ... 34
IV.3. Robustesse de l’effet de dominance visuelle de Colavita aux variations contextuelles. ... 37
IV.4. Effet de la modalité sensorielle sur le niveau d’alerte : vision versus audition. ... 40
V. Expertise, performance en double tâche de TRC et intégration sensorielle. ... 42
V.1. Effet de l’expertise sur la performance perceptivo-motrice en double tâche de TRC... 43
V.2. Effet de l’expertise sur l’évolution des modes d’intégration sensorielle.... 49
PARTIE II - TRAVAUX EXPERIMENTAUX ... 53
Introduction : Objectifs d’étude... 54
Chapitre 3 : Expérience 1 : Intégration visuo-proprioceptive dans une tâche de reproduction passive d’une position spatiale. ... 57
A. Introduction et présentation des hypothèses. ... 57
B. Méthode... 62
C. Résultats. ... 69
D. Discussion. ... 76
Chapitre 4 : Intégration visuo-auditive au cours du temps. ... 87
Introduction et présentation des hypothèses... 87
Expérience 2 : Intégration visuo-auditive et interférence en double tâche de TRC. ... 90
A. Introduction... 90
B. Méthode... 91
C. Résultats. ... 96
D. Discussion. ... 102
Expérience 3 : Robustesse des modes d’intégration visuo-auditive dans une double tâche de TRC. ... 106
A. Introduction... 106
B. Méthode... 108
C. Résultats : effet de l’amorce sur l’ordre des réponses visuelle et auditive. ... 110
A. Introduction... 115
B. Méthode... 117
C. Résultats. ... 118
D. Discussion. ... 125
Discussion... 128
PARTIE III - DISCUSSION GENERALE ... 139
REFERENCES BIBLIOGRAPHIQUES ... 146
INTRODUCTION
Pour interagir d’une manière efficace avec l’environnement, les individus sont amenés à effectuer des tâches plus ou moins complexes dans l’espace et dans le temps. La réalisation de ces tâches nécessite l’intégration par le système nerveux central de plusieurs modalités sensorielles codant des informations provenant d’un ou de plusieurs événements. Par exemple, pour localiser la main dans l’espace, le système nerveux reçoit des informations visuelles et proprioceptives renseignant toutes les deux sur la position de la main. En outre, pour jouer à un jeu vidéo d’action, le système nerveux réagit le plus rapidement possible aux informations visuelles et auditives renseignant chacune sur la réalisation d’une action différente. Que ce soit dans un cas comme dans l’autre, le problème est de savoir comment se réalise l’intégration multi-sensorielle entre ces différentes sources d’informations.
Les études sur l’intégration multi-sensorielle se sont développées et ont fait évoluer la compréhension des mécanismes sous-jacents. Ces mécanismes correspondent à un ensemble d’opérations complexes de transformations multidirectionnelles entre les informations sensorielles et motrices, dans lequel le système nerveux central pondère les différentes modalités sensorielles déterminant plusieurs modes d’intégration. Ces modes d’intégration sont flexibles. Ils dépendent de plusieurs facteurs que nous exposons dans ce travail.
Le but de cette thèse se place dans ce cadre de flexibilité de l’intégration multi-sensorielle. Le but est double : 1/ chercher à déterminer les modes d’intégration des informations visuelles et proprioceptives dans la réalisation d’une tâche de reproduction passive d’une position spatiale, 2/ chercher à déterminer les modes d’intégration des informations visuelles et auditives au cours du temps dans la réalisation d’une double tâche de temps de réaction de choix.
Dans un premier temps (Partie I), nous présentons une revue de littérature concernant la flexibilité de l’intégration multi-sensorielle au niveau comportemental. Un premier chapitre abordera les modèles et les moyens relatifs à l’étude de l’intégration visuo-proprioceptive. Un deuxième chapitre abordera la situation de double tâche de temps de réaction de choix, des aspects de l’intégration visuo-auditive ainsi que l’effet de l’expertise sur la performance en double tâche de temps de réaction de choix et sur les modes d’intégration sensorielle.
Dans un second temps (Partie II), nous présentons nos travaux expérimentaux. Une première expérience (chapitre 3, expérience 1) concerne l’étude de l’intégration visuo-proprioceptive dans une tâche de reproduction passive d’une position spatiale. Par la suite, une série de trois expériences (chapitre 4) concerne l’étude de l’intégration visuo-auditive au cours du temps (expérience 2), sa robustesse en fonction des variations contextuelles (expérience 3), et son évolution avec l’expertise (expérience 4).
Enfin, la discussion générale (Partie III) analyse l’ensemble des résultats pour les discuter dans le cadre de la flexibilité des aspects spatiaux et temporels de l'intégration multi-sensorielle.
PARTIE I
Introduction : Flexibilité de l’intégration multi-sensorielle.
L’intégration multi-sensorielle correspond à un ensemble d’opérations complexes de transformations multidirectionnelles entre les informations sensorielles et motrices dans lequel le système nerveux central pondère les modalités sensorielles. La flexibilité de l’intégration multi-sensorielle, observée au niveau comportemental par des modes d’intégration, est expliquée par une architecture neuronale complexe et par une pondération variable.
Concernant l’architecture neuronale, de multiples travaux ont montré que l’intégration multi-sensorielle n’est pas un processus intermédiaire indépendant entre les afférences et les efférences du système limité aux zones de convergence, notamment aux structures pariétales (Pouget, Deneve & Duhamel, 2002). Elle n’implique pas seulement la convergence anatomique des différents cortex sensoriels spécifiques unimodaux dans des régions cérébrales hétéromodales mais aussi l’interaction entre ces régions (Driver & Spence, 2000 ; Macaluso & Driver, 2005). Elle n’implique pas non plus des aires corticales différentes à des moments différents. Elle se fait par un recrutement simultané de populations de neurones qui sont distribuées dans des aires reliées entre elles par des connexions tout au long du réseau pariéto-frontal. Ces connexions sont assurées par des neurones fonctionnels localisés dans une unité de fonction de base intermédiaire (Pouget & Snyder, 2000 ; Pouget et al., 2002) ou par des neurones combinatoires reliés à des axes fonctionnels (Burnod, Baraduc, Battaglia-Mayer, Guigon, Koechlin, Ferraina, Lacquaniti & Caminiti, 1999). Cette architecture neuronale avec ses propriétés fonctionnelles (Pouget et al., 2002) et/ou combinatoires (Burnod et al., 1999) assure une flexibilité dans l’intégration multi-sensorielle en permettant au réseau neuronal d’assurer les liaisons intermodales et d’exécuter les transformations multidirectionnelles comme les transformations des coordonnées d’une modalité à une autre, des coordonnées sensorielles aux coordonnées motrices, et inversement.
Concernant la pondération des modalités sensorielles par le système nerveux central, elle est variable. En intégrant les informations provenant des différentes
modalités sensorielles, le cerveau affecte un poids plus ou moins important à chacune de ces modalités, suivant des combinaisons linéaires ou non linéaires (cf, Figure 1, d’après Oruc, Maloney & Landy, 2003). Il semble que la combinaison statistique des informations sensori-motrices provenant des différentes modalités ayant une fiabilité intrinsèque et contextuelle variable se fait au mieux selon une règle probabiliste comme le modèle de maximum de vraisemblance (Ernst & Banks, 2002) et les modèles bayesiens (Battaglia, Jacobs & Aslin, 2003 ; Deneve & Pouget, 2004 ; Knill & Pouget, 2004).
Plus le poids accordé à une modalité sensorielle est important, plus la modalité domine dans le traitement perceptivo-moteur réalisé. Toutefois, la dominance sensorielle n’est pas considérée comme le résultat d’un codage unimodal mais plutôt comme un cas particulier de codage multimodal au sein d'un continuum de pondération des informations sensorielles.
Des mécanismes d'optimisation de la pondération attribuée à chaque modalité sensorielle mettent en évidence une flexibilité de l’intégration multi-sensorielle. La variabilité de cette pondération se manifeste par des modes d’intégration flexibles observables au niveau comportemental et auxquels nous nous intéressons dans le cadre de la thèse. Cette flexibilité est déterminée en fonction de plusieurs facteurs dont : la fiabilité de la modalité sensorielle, les traitements réalisés, les transferts inter et intramodaux, l’attention allouée à la modalité sensorielle, les variations contextuelles, l’expertise et les différences entre les individus. Ces facteurs seront développés dans les chapitres suivants, par rapport au contexte spécifique d’étude.
fy fx fx + fy
B
A
C
Fiabilité f iD
Figure 1 : Illustration de la fiabilité bimodale (fi) en fonction de la fiabilité de chacune des modalités x et y (fx et fy). En intégrant les deux modalités sensorielles, fi peut tomber dans une des trois zones A, B ou C suite à des combinaisons linéaires (A- ligne droite et B- courbe du modèle de maximum de vraisemblance) ou non linéaires (C- modèle bayesien). Dans la zone A, fi est entre les fiabilités unimodales. Dans la zone B, fi est mieux que chacune des fiabilités unimodales mais inférieures à la somme des deux. La somme des deux fiabilités unimodales représente la valeur maximale d’une combinaison linéaire dans le cas où x et y ne sont pas corrélés. Dans la zone C, fi est meilleure que la somme des deux fiabilités unimodales. fi ne peut tomber dans la zone D qu’en cas d’interférence entre x et y. (D’après Oruc et al., 2003).
Chapitre 1
Intégration des informations spatiales visuelles et
proprioceptives.
I. Introduction.
Tous les jours, nous sommes amenés à déplacer la main pour saisir des objets, les maintenir dans une position ou à réaliser et reproduire des gestes particuliers. Ces performances nécessitent une bonne précision de la position de la main par rapport au corps qui nécessite la combinaison, par le système nerveux central, des multiples signaux sensoriels et moteurs. Deux sources sensorielles fournissent une information directe sur la position de la main : la proprioception et la vision. Pour interpréter la position de la main par rapport au corps, le système proprioceptif reçoit l'information des angles articulaires entre la main et le corps et/ou des commandes motrices. Pour interpréter la position de la main sur la rétine, le système visuel reçoit l'information de la position de l'oeil dans l'orbite, la position de la tête par rapport au tronc (fournie par la proprioception des muscles de cou) et/ou de la copie d’efférence. Une représentation cohérente, de la localisation de la main nécessite des interactions intermodales. Plus généralement, la précision dans la perception et la réalisation d’une position dépend du nombre des informations codées et de la manière dont elles le sont.
La littérature a largement abordé l’intégration visuo-proprioceptive dans des tâches topocinétiques comme dans les mouvements de pointage, d’atteinte manuelle, dirigés vers des cibles apparentes. Cependant, dans les tâches morphocinétiques, comme dans la reproduction de mouvement ou de position, moins d’études ont tenté de comprendre l’intégration visuo-proprioceptive. Pourtant, une tâche de reproduction est intéressante à investiguer. En effet, la reproduction est une étape du processus d’apprentissage. Par exemple, pour apprendre un mouvement, tout individu perçoit, mémorise et répète ce mouvement dans le but de le maîtriser. Ainsi, toute tâche de
reproduction spatiale offre la possibilité d’optimiser les conditions sensorielles d’un apprentissage perceptivo-moteur.
Dans ce chapitre, nous présentons les modèles d’intégration visuo-proprioceptive mis en évidence à partir des tâches topocinétiques et morphocinétiques, ainsi que les moyens d’étudier cette intégration.
II. Modèles d’intégration visuo-proprioceptive
En intégrant les informations visuelles et proprioceptives, le cerveau affecte un poids à chacune des modalités. Ce poids n’est pas constant. Il varie en fonction de la fiabilité intrinsèque et contextuelle de la modalité sensorielle, de la nature de la computation réalisée au niveau de l’étape du traitement perceptivo-moteur concerné et de l’efficacité des transferts intermodaux.
II.1. Flexibilité en fonction de la fiabilité des modalités sensorielles.
Des études montrent que le système nerveux central se sert de la fiabilité des modalités visuelle et proprioceptive en les combinant (van Beers, Sittig & Denier van der Gon, 1999 ; van Beers, Wolpert & Haggard, 2002b). Une modalité sensorielle est fiable lorsqu’elle est performante, dans son contexte actuel, sous entendant un apport informatif élevé et/ou dont la variabilité est minime. Plusieurs facteurs peuvent influencer la fiabilité des modalités visuelle et proprioceptive : la direction spatiale des mouvements et des positions par rapport au sujet, l’apport informatif de la modalité, la nature des signaux impliqués afférents et/ou efférents (rétiniens versus extra-rétiniens,
actifs versus passifs), la durée du codage spatial et la variabilité interindividuelle (Boulinguez & Rouhana, 2008 ; Coello, Milleville-Pennel & Orliaguet, 2004).
Des modèles récents ont montré que l’intégration de la vision et de la proprioception varie en fonction de la direction spatiale dans laquelle la tâche est accomplie : plan radial ou azimutal. En effet, la vision est plus fiable que la
proprioception dans la localisation proprioceptive d’une cible visuelle (V) ou proprioceptive (P) en direction azimutale qu’en direction radiale et inversement (Boucher, Velay & Paillard, 1992 ; Orliaguet, Coello, Martin & Prablanc, 1995 ; van Beers, Sittig & Denier van der Gon, 1998). Ainsi, dans une situation bimodale où la vision et la proprioception sont disponibles (VP), la vision domine dans le plan azimutal alors que la proprioception domine dans le plan radial (Snijders, Holms & Spence, 2005, 2007 ; van Beers et al., 2002b).
De plus, en manipulant contextuellement l’apport informatif de la vision et de la proprioception, des auteurs ont montré que le poids de l’une de ces modalités augmente avec la diminution des informations fournies par la deuxième modalité (Mon- Williams, Wann, Jenkinson & Rushton, 1997 ; Plooy, Tresilian, Mon-Williams & Wann, 1998). Prenons l’exemple d’un pointage vers une cible définie proprioceptivement et visuellement, quand les informations visuelles sont réduites à la vue de l’extrémité du doigt par l’intermédiaire d’une diode lumineuse, le poids de la proprioception augmente jusqu’à dépasser celui de la vision dans la localisation de la cible (Mon-Williams et al., 1997 ; Mon-Williams, Tresilian & Wann, 1998 ; Plooy et al., 1998). En revanche, quand la main est vue dans un contexte complètement illuminé, le poids de la vision augmente. Ceci, parce que la vision fournit plus d’informations qu’une simple information sur l’extrémité du doigt, comme les informations sur les segments du bras, les autres segments corporels et l’environnement. Dans ce cas, la vision domine la proprioception (Mon-Williams et al., 1997) ou bien contribue avec la proprioception à assurer une meilleure performance de pointage que celles des deux pointages visuel et proprioceptif séparément (van Beers, Sittig & Denier van der Gon, 1996).
A part la direction spatiale et l’apport informatif contextuel de la modalité sensorielle, d’autres facteurs influencent aussi la fiabilité de la modalité visuelle et proprioceptive.
La vision est plus ou moins précise en fonction des signaux impliqués dans le codage spatial et la durée de ce codage. En effet, le codage visuel se fait à partir des signaux rétiniens et extra-rétiniens qui sous entendent aussi la proprioception oculaire et la copie d’efférence. L’interaction entre ces deux signaux améliore le codage visuel. Si un des deux signaux est éliminé, la fiabilité de la vision diminue dans le sens où le codage spatial sera moins performant (Blouin, Gauthier & Vercher, 1995 ; Blouin, Gauthier, Vercher & Cole, 1996 ; Magne & Coello, 2002). Une relation existe entre le type de signaux impliqués et la direction du codage spatial. Alors que les signaux
extra-rétiniens sont performants dans le codage de la direction d’une position spatiale (Coello, 2005), les signaux rétiniens sont plus performants que les signaux extra-rétiniens dans le codage de la distance d’une position (Coello, 2002, 2005) et ce, surtout lorsque le contexte lumineux de la tâche réalisée est structuré (Magne & Coello, 2002 ; Martel & Coello, 2004). De plus, la position rétinienne des signaux visuels joue un rôle dans la direction du codage spatial. En effet, la vision centrale est responsable du contrôle de l’amplitude d’un mouvement (Bard, Paillard, Fleury, Hay & Larue, 1990) alors que la vision périphérique est plus performante que la vision centrale dans le contrôle de la direction du mouvement (Abahnini & Proteau, 1999 ; Bédard & Proteau, 2001 ; Paillard, 1982). Toutefois, au regard de la dominance des signaux rétiniens structurés dans le codage de la distance d’une position ou de l’amplitude d’un mouvement, la remarque suivante peut se faire. Le fait que la vision soit moins précise que la proprioception dans le plan radial peut être du au contexte lumineux du codage spatial. En effet, cette observation a été faite lorsque la vision fournit des informations sur l’extrémité du membre effecteur par une diode lumineuse dans un contexte peu lumineux et souvent dans le noir. Alors que lorsque le codage visuel se fait dans un contexte structuré lumineux, favorisant les signaux rétiniens, la précision de la vision dans le codage de la distance augmente (Martel & Coello, 2004). Quoi qu’il en soit, avec le temps, la précision du codage visuel diminue. Toutefois, l’effet de la durée qui sépare le positionnement d’une cible visuelle et son repérage ou celui de la rétention mnésique visuelle sur la performance de pointage et/ou de reproduction mnésique est controversé (Boucher et al., 1992 ; Posner, 1967).
Quant à la proprioception, elle est plus ou moins précise en fonction de la nature active ou passive et du temps du codage spatial. Par exemple, le déplacement actif d’un bras sollicite les commandes centrales et donc la génération d’une commande motrice. Par contre, lors d’un déplacement passif du bras, les signaux centraux reliés aux commandes motrices sont absents et ne peuvent pas calibrer les signaux afférents. Par conséquent, la localisation d’une cible proprioceptive est beaucoup plus précise quand la main est déplacée d’une façon active (volontairement) que lors d’un déplacement passif (main guidée par l’expérimentateur) (Paillard & Brouchon, 1968, 1974). La dégradation de la performance, lorsque seuls les signaux afférents proprioceptifs sont sollicités, est plus prononcée au niveau du codage de la direction que de l’amplitude d’un mouvement (Adamovich, Berkinblit, Fookson & Poizner, 1998). Toutefois, en
d’autant plus importante que la proprioception n’est pas calibrée par la copie d’efférence (Craske & Crawshaw, 1975) et ce, à partir de deux secondes (Paillard & Brouchon, 1968, 1974). En revanche, Desmurget et ses collaborateurs (2000) ont montré que la dérive proprioceptive ne survient pas avant vingt secondes. Ce qui montre que l’hypothèse d’une dérive proprioceptive est délicate et que des facteurs attentionnels et mnésiques peuvent entrer en jeu expliquant les divergences de la littérature (Desmurget, Vindras, Gréa, Viviani & Grafton, 2000 ; Horch, Clark & Burgess, 1975).
Les facteurs cités ci-dessus influencent la fiabilité relative des modalités visuelle et proprioceptive dans la perception et le contrôle moteur. Dans la plupart des études sur l’intégration visuo-proprioceptive, la perception ou le contrôle moteur visuel est associé à la proprioception oculaire. De même, la proprioception des membres corporels est souvent active donc elle sollicite la copie d’efférence. Pour comprendre l’intégration des afférences visuelles et proprioceptives, il faudrait éviter les mouvements des yeux et effectuer des déplacements passifs du membre corporel étudié. Nous connaissons le rôle des afférences dans la précision perceptivo-motrice sur le plan unimodal visuel ou proprioceptif. Toutefois, sur le plan bimodal, l’intégration de ces afférences reste peu connue.
II.2. Flexibilité en fonction de l’étape du traitement perceptivo-moteur.
Des études mettent en évidence une flexibilité de l’intégration visuo-proprioceptive en fonction de l’étape du traitement perceptivo-moteur (Bagesteiro, Sarlegna & Sainburg, 2006 ; Boulinguez & Rouhana, 2008 ; Lateiner & Sainburg, 2003 ; Sheidt, Conditt, Secco & Mussa-Ivaldi, 2005 ; Sober & Sabes, 2003). En effet, dans la planification d’un mouvement dirigé vers une cible visuelle, Sober et Sabes (2003) ont distingué deux étapes : la formation du vecteur de mouvement et la transformation de ce vecteur en commande motrice. La formation du vecteur du mouvement se fait à partir de l’estimation de la position initiale de la main et de l’estimation de la position de la cible. La transformation de ce vecteur en une commande motrice nécessite une bonne connaissance de la position initiale de la main. Bien que la performance de ces deux étapes soit influencée par la précision de la
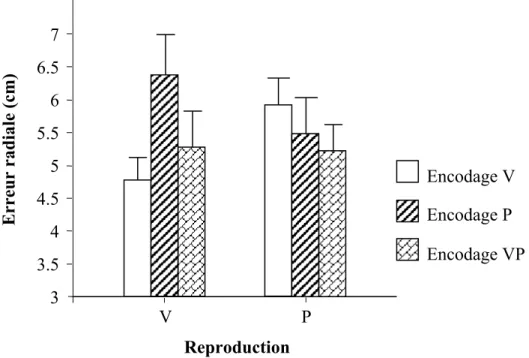
position initiale de la main, Sober et Sabes (2003) ont montré qu’elles reposent chacune sur une estimation différente de cette position à l’issue d’une intégration visuo-proprioceptive différente. La formation du vecteur du mouvement repose d’une façon dominante sur la vision (Sober & Sabes, 2003) et ce, en termes de direction (Lateiner & Sainburg, 2003) et de distance (Bagesteiro et al., 2006). En revanche, la transformation de ce vecteur en commande motrice repose d’une façon plus importante sur la proprioception (Sober & Sabes, 2003). Ce rôle de la proprioception ne se limite pas à la formation de la commande motrice mais il est important aussi pour le contrôle et les corrections en ligne du mouvement jusqu’à sa phase terminale (Bagesteiro et al., 2006). Cette différence des mécanismes d’intégration visuo-proprioceptive en fonction de la computation réalisée n’existe pas uniquement dans la planification d’un mouvement dirigé vers une cible visuelle. Elle est montrée aussi dans une tâche de reproduction d’un mouvement préalablement encodé. En effet, Boulinguez et Rouhana (2008) ont mis en évidence une intégration visuo-proprioceptive qui diffère lors de la phase d’encodage et lors de la phase de reproduction. En fonction de la variabilité interindividuelle, une dominance, soit visuelle, soit proprioceptive, s’installe lors de chacune des deux phases, la modalité non dominante étant complètement ignorée.
La flexibilité de l’intégration visuo-proprioceptive en fonction de l’étape du traitement perceptivo-moteur n’est pas étonnante. Déjà, à un niveau unimodal, les informations visuelles sont utilisées d’une manière différente selon la tâche réalisée. Par exemple, dans un mouvement dirigé vers une surface inclinée sur laquelle il faut poser un objet, le cerveau intègre les signaux binoculaires et monoculaires en donnant un poids plus important aux signaux binoculaires dans le contrôle du mouvement du bras que dans le jugement perceptif de la surface de placement de l’objet (Knill, 2005).
II.3. Flexibilité en fonction de l’efficacité des transferts intermodaux.
L’intégration de la vision et de la proprioception varie non seulement en fonction de la fiabilité de l’information véhiculée par la modalité sensorielle et de l’étape du traitement perceptivo-moteur, mais aussi en fonction de l’efficacité du transfert intermodal visuo-proprioceptif.
Il existe deux théories qui concernent les transferts intermodaux. Une première qui postule une transformation des coordonnées spatiales visuelles en coordonnées proprioceptives (vision-proprioception, V-P) et/ou inversement (proprioception-vision, P-V) (Connoly & Jones, 1970) et une deuxième qui postule l’absence de ses transformations de coordonnées (Gibson, 1966). Selon la première théorie, le transfert intermodal est plus efficace que le transfert intramodal (vision-vision, V-V ou proprioception-proprioception, P-P) (Orliaguet, 1981). Concernant les transferts intermodaux, la performance de P-V diffère souvent de la performance de V-P. Toutefois, ces performances sont conditionnées par la précision et la capacité mnésique des modalités visuelle et proprioceptive. En effet, dans le jugement perceptif d’une taille, les auteurs ont montré que la performance de V-P est moins efficace que celle de P-V (Connolly & Jones, 1970 ; Jones & Connolly, 1970) parce qu’un stockage en mémoire se fait dans la modalité dans laquelle la réponse est demandée et que la modalité visuelle est plus performance que la proprioception dans le domaine de la mémorisation spatiale (Jones & Connolly, 1970 ; Posner, 1967). Contrairement, dans une tâche d’appariement d’une position corporelle, les auteurs ont observé que la performance de P-V est plus efficace que la performance de V-P (Euzet & Gahéry, 1995, 1996, 1998). Selon la deuxième théorie, les modalités sensorielles sont capables de recueillir des informations spatiales de même nature et au même moment. Ainsi, si au niveau perceptif, les coordonnées spatiales sont détachables du canal sensoriel et se présentent sous forme amodale, alors elles peuvent être transférées vers une autre modalité sensorielle sans avoir recours à des transformations. Par conséquent, le transfert intermodal (V-P ou P-V) serait aussi efficace que le transfert intramodal (V-V ou P-P).
Toutefois, les observations des études comportementales ont appuyé la première théorie. Lors d’un transfert intermodal, le système nerveux central effectue des transformations des coordonnées spatiales d’une des deux modalités sensorielles en coordonnées spatiales de l’autre. Comme exemple, la transformation des coordonnées sensorielles de la main en coordonnées de la cible ou aussi la transformation des coordonnées de la modalité sensorielle de l’encodage d’une configuration ou d’un mouvement en coordonnées de la modalité de reproduction. Or, ces transformations sont une source d’erreur supplémentaire. De multiples études ont montré que l’intégration visuo-proprioceptive se fait en minimisant les erreurs liées à ces transformations et ce, en évitant de réaliser des transferts intermodaux. Par exemple,
lors de la planification du vecteur d’un mouvement dirigé vers une cible, l’intégration de la vision et de la proprioception varie en fonction de la modalité sensorielle de la cible. La planification de la distance du mouvement repose d’une manière dominante sur la vision lorsque la cible est visuelle alors qu’elle repose sur la proprioception lorsque la cible est proprioceptive (Sarlegna & Sainburg, 2007). Concernant la planification de la direction du mouvement, deux cas de figures sont mis en évidence lorsque la cible est visuelle. Certains auteurs ont montré que la vision et la proprioception sont combinées pour donner une meilleure représentation de la main dans sa position initiale (Rossetti, Desmurget & Prablanc, 1995). D’autres auteurs ont montré que l’estimation de la position de la position initiale de la main est dominée par la vision lorsque la cible est visuelle, alors qu’elle se fait par une combinaison d’une façon égale des informations visuelles et proprioceptives lorsque la cible est proprioceptive (Sober & Sabes, 2005). Quoi qu’il en soit, le rôle accru de la vision quand la cible est visuelle ainsi que celui de la proprioception lorsque la cible est proprioceptive sont expliquées par le fait que le cerveau minimise les erreurs liées aux transformations des coordonnées spatiales de la main vers les coordonnées spatiales de la cible définies dans une modalité sensorielle différente. De même, pour reproduire une configuration spatiale du bras encodée dans une situation bimodale conflictuelle, Orliaguet (1985) a montré que les sujets ont tendance à privilégier la modalité visuelle pour percevoir la position du bras lorsque la modalité de reproduction est visuelle alors que lorsque la modalité de reproduction est proprioceptive les sujets privilégient la modalité proprioceptive. Ceci, pour éviter la réalisation d’un transfert intermodal entre la phase d’encodage perceptif et la phase de reproduction.
Selon les études citées dans cette partie, la dominance de l’information visuelle ou proprioceptive serait due à une stratégie de minimisation des erreurs liées aux transformations entre les coordonnées des modalités visuelle et proprioceptive. Toutefois, si ces transformations n’engendrent pas d’erreurs, alors le cerveau n’aurait pas de raison de les éviter. Si ce dernier cas est possible, le transfert intermodal ne serait pas moins efficace qu’un transfert intramodal. Ainsi pour étudier l’intégration visuo-proprioceptive, il est important de vérifier l’efficacité des transferts d’informations entre les modalités visuelle et proprioceptive.
III. Questions méthodologiques relatives à l’étude de l’intégration
visuo-proprioceptive.
Pour étudier l’intégration visuo-proprioceptive et comprendre comment le cerveau combine ces informations, plusieurs points sont pris en compte. Il s’agit de 1/ la précision de chacune des modalités visuelle et proprioceptive dans la précision spatiale, 2/ l’efficacité du transfert intermodal par rapport au transfert intramodal et 3/ la précision en situation bimodale lorsque les modalités visuelle et proprioceptive sont disponibles simultanément. Nous présentons dans la partie suivante les principales méthodes d’études, leurs intérêts et limites.
III.1. Précision de la modalité visuelle (V) et de la modalité proprioceptive (P).
La précision de V et de P peut être évaluée de deux façons. Une façon directe est utilisée dans les tâches de reproduction. Elle consiste à comparer les performances spatiales obtenues dans des situations d’encodage et de reproduction identiques V (V-V) et P (P-P) appelées conditions intramodales (Boulinguez & Rouhana, 2008 ; Orliaguet, 1981, 1983). Une deuxième façon est indirecte et elle est utilisée dans des tâches d’appariement (van Beers et al., 1998). Elle consiste à dériver la précision à partir de trois conditions : localisation proprioceptive avec la main droite d’une cible visuelle, localisation proprioceptive avec la main gauche d’une cible visuelle et localisation proprioceptive avec la main gauche d’une cible définie par la main droite.
III.2. Le transfert d’information entre la vision et la proprioception.
Pour tester le transfert d’information entre les modalités visuelle et proprioceptive, des conditions intermodales sont souvent proposées. Une présentation1
1 Le terme présentation désigne la phase d’encodage dans les tâches de reproduction ou la réalisation du mouvement dans les mouvements dirigés vers une cible.
unimodale du stimulus (V ou P) est suivie d’un test2 unimodal sollicitant cependant une modalité sensorielle différente de celle de la présentation. Les performances de ces conditions intermodales (V-P et P-V) sont ensuite comparées à celle de la condition intramodale la moins précise (V-V ou P-P) pour évaluer l’efficacité du transfert intermodal (Orliaguet, 1981). De plus, les performances des conditions intermodales (V-P et P-V) sont comparées entre elles pour évaluer l’efficacité du transfert de la modalité visuelle vers la modalité proprioceptive et inversement (Connolly & Jones, 1970).
III.3. Précision en situation bimodale VP.
Pour comprendre comment le cerveau combine les informations visuelles et proprioceptives lorsqu’elles sont toutes les deux disponibles (situation bimodale VP), plusieurs méthodes sont utilisées. Ces méthodes sont groupées sous deux grands axes : le paradigme de discordances sensorielles et le paradigme de suppression/ajout d’informations sensorielles.
Paradigme de discordances sensorielles.
Le principe consiste à introduire un conflit perceptif entre les informations visuelles et proprioceptives dans les situations bimodales. Le but étant de créer une discordance entre la position réelle de la main perçue proprioceptivement et celle perçue visuellement. La discordance peut être introduite de plusieurs façons. Le plus souvent, elle se fait par l’intermédiaire de lunettes prismatiques (Boulinguez, Toussaint, Abed-Meraim & Collignon, 2001 ; Orliaguet, 1986 ; Rossetti et al., 1995 ; van Beers et al., 2002b). Elle peut se faire aussi à l’aide de la vision indirecte par l’intermédiaire d’un mannequin (Orliaguet, 1985) ou d’un miroir (Holmes, Crozier & Spence, 2004 ; Holmes & Spence, 2005). Les lunettes prismatiques, placées devant les yeux des sujets, peuvent déplacer le champ visuel dans les plans azimutal et radial, de façon à dévier la position réelle de la main codée sur la base des informations proprioceptives et celle codée sur la base des informations visuelles. Le miroir crée une illusion perceptive, qui est d’autant plus importante que la période de fixation est longue. Par exemple, regarder
dans le miroir deux mains dont les positions réelles sont distalement décalées par rapport au corps fait qu’au bout d’un certain temps ces positions sont perçues comme étant parallèles et à égale distance du corps (Holmes et al., 2004).
En introduisant des discordances visuo-proprioceptives, trois méthodes sont utilisées :
La première méthode consiste à proposer trois phases expérimentales successives : une phase de préadaptation, une phase d’adaptation et une phase de post-adaptation (van Beers et al., 2002b). La perturbation visuo-proprioceptive est introduite dans la phase d’adaptation. Par la suite, elle est réduite graduellement pour assurer une désadaptation. L’influence de la perturbation sur la performance spatiale de chacune des modalités est évaluée en comparant les phases de pré- et post-adaptation. La modalité qui s’adapte le plus est celle dont le poids est plus faible et donc la moins dominante.
La deuxième méthode consiste à tester la performance spatiale d’un test unimodal proprioceptif après une présentation bimodale avec un conflit visuo-proprioceptif (Boulinguez et al., 2001). Plus cette performance va dans le sens de la déviation visuelle, plus la contribution de la modalité visuelle est dominante.
La troisième méthode consiste à comparer les performances spatiales entre deux situations bimodales dont une comprend un conflit visuo-proprioceptif (Orliaguet, 1985 ; Rossetti et al., 1995). Ce qui permet d’estimer l’importance de la modalité sensorielle qui a été perturbée. Plus les performances des deux situations bimodales sont proches, moindre est le poids de la modalité sensorielle perturbée. Par contre, plus ces performances sont différentes et surtout dans le sens de la perturbation, plus la modalité sensorielle perturbée est dominante.
Paradigme de suppression/ajout d’informations sensorielles.
Le principe de suppression ou d’ajout des informations sensorielles est utilisé dans le paradigme de transfert d’apprentissage et dans les comparaisons entre situations bimodales et unimodales.
Le paradigme de transfert d’apprentissage consiste à proposer des phases d’acquisition de durées spécifiques (courtes ou longues) suivies par des phases de transfert. Dans la phase de transfert, des informations sensorielles sont ajoutées, supprimées ou restent identiques. La comparaison entre les résultats obtenus en phase de transfert et ceux obtenus en phase d’acquisition permet de mettre en évidence le poids relatif des informations sensorielles. La détérioration de la performance en
transfert, lorsque le contexte est différent de celui de l’acquisition, souligne la dominance des informations sensorielles présentes au cours de l’apprentissage (Proteau & Cournoyer, 1990 ; Proteau, 1992 ; Robin, Toussaint, Blandin & Proteau, 2005).
Le principe de suppression ou d’ajout des informations sensorielles s’applique aussi lors des comparaisons entre les situations bimodales (avec ou sans conflit perceptif) et les situations unimodales. Lorsque la performance des situations bimodales n’est pas différente de celle des situations unimodales, cela indique que la modalité de la situation unimodale est dominante, l’apport informatif de la modalité sensorielle qui a été supprimée étant quasiment nul (Boulinguez & Rouhana, 2008 ; Orliaguet, 1986). En revanche, lorsque cette performance est différente de celle des situations unimodales, cela indique que l’apport informatif de la modalité supprimée est important (Mon-Williams et al., 1997 ; Plooy et al., 1998). Plusieurs cas de figures peuvent être observés : une performance meilleure que celle des situations unimodales (van Beers et al., 1996) ou une performance intermédiaire (van Beers et al., 1999).
Intérêts et limites.
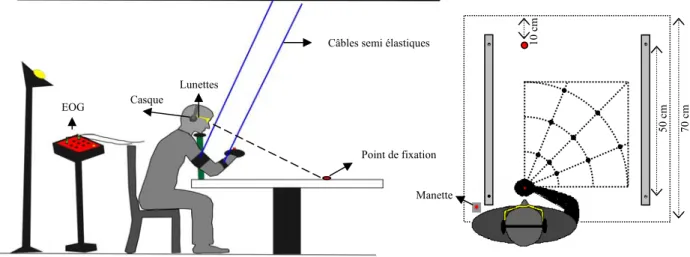
Les méthodes présentées ci-dessus permettent de comprendre l’intégration visuo- proprioceptive mais elles présentent des limites. L’utilisation d’un conflit perceptif, créant une discordance entre les informations visuelles et les informations proprioceptives, a un effet qui masque les mécanismes spontanés impliqués dans l’intégration sensorielle. Les sujets s’adaptent au conflit par l’intermédiaire d’une recalibration proprioceptive et d’un apprentissage visuo-moteur (Welch, 1986). De même, en utilisant une méthode d’adaptation ou d’apprentissage, les sujets s’adaptent à la tâche rapidement et d’une manière efficace en développant un modèle interne (Mackrous & Proteau, 2007) et des transformations appropriées après peu d’entraînement (Baraduc, Guigon & Burnod, 2001). Ces changements sensorimoteurs surviennent d’un essai à l’autre (Bernier, Chua & Franks, 2005 ; Redding & Wallace, 2004). De plus, la plupart des études interroge l’intégration visuo-proprioceptive dans des tâches de pointage, de mouvements dirigés vers une cible et dans des tâches de reproduction, en manipulant la disponibilité des informations sensorielles soit lors de la présentation du stimulus soit lors du test (voir le tableau 1 pour une illustration).
Pourtant pour comprendre les mécanismes impliqués dans l’intégration visuo-proprioceptive, il est intéressant de prendre en compte tous les facteurs qui peuvent intervenir et ce, en manipulant la disponibilité des informations sensorielles lors de la présentation et du test à la fois. Dans une tâche de reproduction de mouvement, une indépendance entre les phases d’encodage et de reproduction au niveau des modes d’intégration visuo-proprioceptive a été observée (Boulinguez & Rouhana, 2008). Par conséquent, dans ce type de tâche, il peut être intéressant de manipuler la présence des informations visuelles et proprioceptives lors des phases d’encodage et de reproduction pour tester l’intégration visuo-proprioceptive.
L’étude de Boulinguez et Rouhana (2008) résout les problèmes liés aux limites précédentes. Elle propose une méthode de manipulation de la disponibilité des modalités visuelle et proprioceptive en encodage et en reproduction à la fois, sans introduire de discordances sensorielles et en limitant le nombre des essais expérimentaux. Ces mesures méthodologiques supposent que lors de l’intégration visuo-proprioceptive, la recalibration proprioceptive, l’apprentissage visuo-moteur, la formation de modèle interne et l’adaptation d’un essai à l’autre ne sont pas possibles.
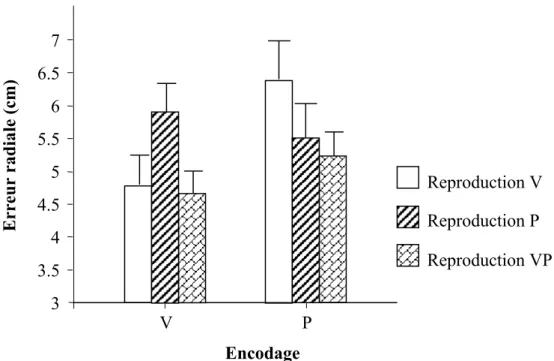
Tableau 1 : Illustration de la manipulation des informations visuelles (V) et proprioceptives (P) lors de la présentation (modalité du mouvement ou modalité d’encodage) et lors du test (modalité de la cible ou modalité de reproduction). La situation bimodale est représentée par VP, la situation unimodale par V ou P.
Présentation Test
van Beers et al, 1996
VP P V P P P Plooy et al, 1998 P P VP V
van Beers et al, 1999
P P P VP P V Holmes et al, 2004 P P VP P Mon-Williams et al, 1997 P P VP P Orliaguet, 1986 VP P V P P P
Pour cette raison, pour investiguer l’intégration visuo-proprioceptive dans une tâche de reproduction spatiale, nous nous baserons sur la méthodologie employée par Boulinguez et Rouhana (2008) comme point de départ. Des modifications seront apportées en fonction des questions que nous nous posons et qui seront exposées dans la partie II, chapitre 3.
Chapitre 2
Intégration des informations visuelles et auditives au cours du
temps.
I. Introduction.
Dans les parties précédentes, nous avons examiné la flexibilité des mécanismes d’intégration sensorielle qui se focalisent sur la compréhension de la manière par laquelle le cerveau intègre la vision et la proprioception dans la perception et le contrôle moteur. Cependant, les aspects temporels de l’intégration n’ont pas été soulevés. Pourtant, un problème important dans le traitement sensoriel devrait être clarifié : il s’agit de la manière dont le cerveau accumule et utilise les informations sensorielles au cours du temps.
Pour étudier cet aspect temporel, il est nécessaire de mettre en place une situation expérimentale adéquate. Tout d’abord, il est important d’introduire une concurrence temporelle entre les informations fournies par les différentes modalités sensorielles. La majorité des investigations portant sur l’intégration sensorielle dans les tâches spatiales, concernent des mouvements dirigés vers des cibles, des mouvements de pointage, de repositionnement etc... où une concurrence spatiale est introduite entre les modalités sensorielles disponibles (déviation prismatique par exemple). Dans ces tâches, les deux modalités sensorielles présentes renseignent sur le même évènement (localisation de la cible, position de la main). Toutefois, pour comprendre comment les informations sont cumulées et utilisées en fonction du temps, il est nécessaire de proposer au moins deux évènements différents. Ce qui nous amène à considérer une situation de double tâche de temps de réaction (TR), où deux tâches, sollicitant chacune une modalité sensorielle différente, doivent être réalisées. Il est important que les deux tâches interfèrent au niveau exécutif central. Pour cela, elles doivent être concurrentes sollicitant chacune l’étape de sélection de réponse. Généralement, cette situation pousse le système de traitement des informations à atteindre ses limites offrant ainsi une
situation intéressante pour comprendre les mécanismes d’intégration des informations sensorielles.
Dans la partie suivante nous évoquons les données qui aident à comprendre la situation de double tâche de temps de réaction de choix et l’interférence qu’elle génère, ainsi que leur utilité dans la compréhension de certains aspects temporels de l’intégration sensorielle.
II. Attention et double tâche de temps de réaction de choix.
II.1. Différence entre tâches de TR simple (TRS) et TR de choix (TRC).
Les tâches de TR trouvent leur origine dans les travaux de Donders (1868), qui voulait mesurer «la vitesse des actes psychiques ». Dans une tâche de TR, le sujet est invité à répondre le plus rapidement et le plus précisément possible à un stimulus sensoriel particulier. Le TR représente le temps minimal nécessaire pour produire une réponse correcte à une stimulation donnée. Il désigne alors la mesure chronométrique du temps qui sépare le début de la présentation d’un stimulus (entrée afférente) et le déclanchement de la réponse motrice volontaire à ce stimulus (sortie efférente). Il se déroule selon le schéma suivant : activation d'unités sensorielles par un stimulus, traitement de l'information par des neurones du système nerveux central (cerveau et moelle épinière), activation des unités motrices (Marieb, 1999). Ce traitement par le système nerveux central est plus ou moins compliqué selon la situation. Nous parlons de temps de réaction simple (TRS), lorsqu’un seul stimulus est présenté au sujet et une seule réponse motrice lui est demandée. Par contre, nous parlons de temps de réaction de choix (TRC) lorsque plusieurs stimuli sont présentés, incitant le sujet à faire un choix entre au moins deux réponses qui correspondent chacune à un stimulus.
Toute tâche de TR mobilise des ressources attentionnelles. Cette mobilisation est plus importante dans la réalisation d’une tâche de temps de réaction de choix que dans la réalisation d’une tâche de temps de réaction simple (Pashler, 1998). La principale
TRC, puisque celle-ci n’est pas unique et ne dépend pas de la seule apparition du stimulus mais de l’identification de sa classe d’appartenance et par conséquent elle sollicite des traitements centraux. Pour cette raison, nous évoquons dans la partie suivante les aspects de l’attention et ses fonctions dans la réalisation simultanée de deux tâches de TRC appelée double tâche de TRC.
II.2. L’attention : ses aspects et fonctions.
L’attention contrôle, module et supervise la quasi-totalité de nos activités perceptives et motrices. Que l’attention soit endogène ou exogène, son résultat est un filtrage et un engagement de ressources ayant une fonction d’aide, de régulation et d’optimisation des différentes étapes de traitement de l’information.
L’attention sélective et divisée.
L’homme est souvent confronté à de multiples informations qu’il faut traiter avec le plus de pertinence possible. La limitation des capacités de traitement implique : i) l’existence de plusieurs processus de sélection des informations les plus utiles afin de les traiter en profondeur et de faire abstraction des autres informations (attention sélective) et ii) le partage des ressources attentionnelles sur la multitude des informations utiles à traiter simultanément (attention divisée).
L’attention a ainsi une fonction de filtre et de tri des informations. Ce système de filtre peut s’appliquer sur les systèmes sensoriels, le système de mémorisation comme sur les systèmes exécutifs et moteurs. L’un des premiers modèles de l’attention a été proposé par Broadbent (1958) et repose sur la métaphore du filtre positionné en amont du traitement. Les informations pénétrant dans le système nerveux sont stockées temporairement dans une mémoire à court terme. Un filtre intervient en ne permettant l’accès qu’à un nombre restreint d’informations dans les étapes suivantes du traitement. Toutefois, l’existence de différents stades de traitement suggère que l’attention est susceptible d’intervenir comme un filtre à plusieurs étapes de ce traitement. Depuis Broadbent (1958), plusieurs modèles ont été proposés en variant la position présumée du filtre attentionnel dans le processus du traitement de l’information. Toutefois, quel que soit le modèle, le filtre attentionnel doit faciliter le traitement de l’information cible et altérer le traitement du distracteur.
Lorsqu’au moins deux informations sont pertinentes et doivent être traitées simultanément, le système fait appel à l’attention divisée. L’attention est aussi considérée comme un réservoir de capacités (ressources attentionnelles) susceptibles d’être investies en plus ou moins grande quantité (intensité) dans les différentes opérations mentales effectuées par le système de traitement (Kahneman, 1973). Le modèle de kahneman (1973) considère que notre système de traitement de l’information ne possède qu’un seul réservoir de ressources qui a une capacité limitée. Plus les ressources sont investies dans une tâche, meilleur est le traitement correspondant, et inversement. A l’opposé, d’autres théoriciens ont proposé l’existence d’une multitude de réservoirs de ressources attentionnelles différenciés, où chaque traitement vient de manière spécifique puiser la quantité de ressources dont il a besoin (Navon & Gopher, 1979 ; Wickens, 1980 ; Friedman & Polson, 1981).
L’attention exogène et endogène.
L’attention sélective et l’attention divisée peuvent être exogènes (dirigées par un stimulus externe) ou endogènes (contrôlées et volontaires) (Schneider & Shiffrin, 1977).
L’attention d’un individu peut être attirée (capturée) vers une information du monde extérieur, plus ou moins indépendamment de sa volonté : c’est l’attention exogène. Elle dépend des processus automatiques de traitement, concernant le plus souvent des informations soudaines et simples. Les effets en sont brefs si les processus endogènes ne viennent pas prendre le relais.
L’attention endogène est, quant à elle, subjective et volontaire. L’individu peut lui-même diriger son attention vers un certain endroit ou un certain type d’information en fonction de plusieurs facteurs. Cette composante est au cœur du processus de contrôle, dans la mesure où, même si l’attention de l’individu est attirée de manière exogène vers une information, cette information ne restera dans le foyer attentionnel que si l’individu le décide en maintenant l’attention de manière endogène (LaBerge, 1995). L’attention serait un équilibre entre les processus endogènes, permettant d’assurer la cohérence d’un comportement orienté vers un but, et les processus exogènes permettant une flexibilité de ce comportement aux modifications de l’environnement (Allport, 1989).
La limitation attentionnelle : interférence en double tâche.
Comme son nom l’indique, la double tâche est une situation expérimentale, où l’on demande à l’individu de réaliser obligatoirement deux tâches. Dans cette situation, le système traite simultanément deux informations pertinentes. Par conséquent, le système fait appel à l’attention divisée. Toutefois, que le réservoir de ressources attentionnelles à partager soit unique ou multiple, il est limité dans ses capacités. Cette limitation attentionnelle se manifeste alors par une interférence entre les deux tâches réalisées, interférence qui se manifeste par la dégradation de la performance d’une ou des deux tâches. L’interférence dans une tâche de temps de réaction de choix est généralement plus importante que dans une tâche de temps de réaction simple, et ce à cause de la difficulté de traitement au niveau des étapes centrales. Cette interférence est majoritairement expliquée par deux modèles que nous détaillerons dans la partie suivante.
III. Modèles d’interférence et supports expérimentaux.
III.1. Les modèles d’interférence.
Les deux modèles majeurs qui expliquent l’interférence en situation de double tâche sont le modèle de partage des capacités et le modèle de goulot d’étranglement (Pashler, 1994a, 1998).
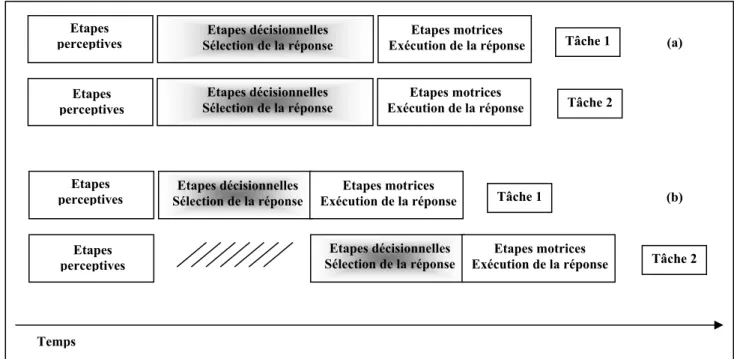
La manière la plus acceptée pour expliquer l’interférence en double tâche est d’assumer le partage des capacités de traitement entre les tâches. Comme nous l’avons vu dans la partie précédente, ces capacités proviennent d’un seul ou de multiples réservoirs. Toutefois quel que soit le nombre de réservoirs, puisque les capacités attentionnelles sont limitées et puisqu’elles sont partagées entre deux tâches, alors il y a moins de capacités pour chaque tâche individuelle et par conséquent la performance se dégrade. Dans une double tâche de TRC, le partage des capacités a lieu le plus souvent au niveau des étapes décisionnelles ou de sélection de la réponse (Figure 2 a). Deux idées clés caractérisent le modèle de partage des capacités. La première est que le
traitement des deux tâches se fait en parallèle avec une efficacité qui dépend de la quantité des capacités allouées aux deux tâches. La deuxième est que les individus peuvent varier volontairement la quantité d’allocation des capacités à chacune des deux tâches (Tombu & Jolicoeur, 2002).
Quant au modèle du goulot d’étranglement, il suppose que certaines opérations mentales ne peuvent pas être opérées en parallèle. Certaines opérations pourraient avoir besoin qu’un mécanisme unique leur soit dédié pour une période de temps. Lorsque les deux tâches ont besoin du même mécanisme en même temps, un goulot d’étranglement survient (Welford, 1952). Il peut y avoir un seul ou plusieurs goulots d’étranglement associés chacun à des étapes ou des opérations mentales différentes. Dans une double tâche de TRC, ces étapes critiques sont le plus souvent les étapes décisionnelles ou les étapes de sélection de la réponse qui ne peuvent opérer que d’une façon séquentielle (Figure 2 b). En contraste, les traitements des étapes qui précèdent ou suivent les étapes décisionnelles peuvent opérer en parallèle (Pashler, 1984). De ce fait, le modèle de goulot d’étranglement est un modèle de traitement mixte, sériel et parallèle.
Les deux modèles de partage de capacités et de goulot d’étranglement ont été proposés et validés à partir de l’interférence mise en évidence par le paradigme de la période réfractaire psychologique (PRP). Dans la partie suivante, nous détaillons le
Temps Etapes
perceptives Sélection de la réponseEtapes décisionnelles
Etapes motrices Exécution de la réponse Etapes perceptives Etapes décisionnelles Sélection de la réponse Etapes motrices
Exécution de la réponse Tâche 2
Tâche 1 (b)
Etapes
perceptives Sélection de la réponse Etapes décisionnelles
Etapes motrices Exécution de la réponse Etapes
perceptives
Etapes décisionnelles
Sélection de la réponse Exécution de la réponse Etapes motrices Tâche 2
Tâche 1 (a)
Figure 2: Modèle de partage des capacités (a), modèle du goulot d’étranglement (b) et traitement d’une double tâche de temps de réaction de choix.
III.2. Le paradigme expérimental.
Paradigme PRP : procédure générale.
L’interférence en double tâche de TRC est majoritairement étudiée par le paradigme PRP. La PRP est la période réfractaire psychologique par analogie à la période réfractaire des neurones (Telford, 1931). En effet, Telford (1931) et Vince (1949) ont montré qu’en présentant deux tâches de temps de réaction, nécessitant deux réponses rapides à deux stimuli différents, le temps de la réponse au deuxième stimulus est ralenti. En manipulant plusieurs variables, le paradigme PRP est utilisé par un vaste nombre d’études pour déterminer si les deux tâches impliquent les mêmes ressources attentionnelles et pour comprendre comment le système s’organise face à la limitation attentionnelle. Il permet plus précisément de savoir si le traitement se fait de façon mixte (parallèle et sérielle) avec un goulot d’étranglement, ou bien de façon parallèle avec un partage égale des capacités entre les deux tâches.
Dans le paradigme PRP, deux stimuli (S1 et S2) sont présentés en succession rapide, séparés par une période variable appelée SOA (stimulus onset asynchrony) ou aussi IIS (intervalle inter-stimuli). Le sujet doit émettre une réponse à chaque stimulus (R1 et R2, respectivement) le plus rapidement possible. Ce paradigme comprend deux types de conditions proposées (Pashler & Johnston, 1989 ; Ruthruff, Pashler & Hazeltine, 2003b) : une condition de tâche simple et une condition de double tâche. Dans la condition de tâche simple, un seul stimulus S1 ou S2 est présenté alors que dans la condition de double tâche, les deux stimuli S1 et S2 sont présentés. Selon l’objectif de l’utilisation de ce paradigme, plusieurs facteurs sont manipulés par l’expérimentateur et trois variables sont principalement observées.
Principales variables observées.
Les principales variables sont : le ralentissement du temps de réaction de la première et la deuxième tâche présentée (T1 et T2, respectivement) ainsi que l’intervalle inter-réponses (IIR).
Les ralentissements de T1 et T2 peuvent être mesurés de deux manières. Soit ils sont mesurés par la différence entre le temps de réaction à T1 et T2 en condition de double tâche (TR1d et TR2d, respectivement) et le temps de réaction à T1 et T2 en condition de tâche simple (TR1s et TR2s, respectivement). Dans ce cas, ils sont appelés PRP1 et PRP2 et représentent la quantité d’interférence susceptible d’agir sur T1 et T2,
respectivement. Soit ils sont mesurés par les différences entre deux TR1d ou TR2d recueillis dans deux conditions de double tâche qui diffèrent au niveau des facteurs étudiés. Dans ce cas, ils renseignent sur la nature et le locus de l’interférence.
Toutefois, l’étude du ralentissement de chacune des deux tâches n’a pas de sens si le compromis rapidité-précision n’est pas contrôlé. L’expérimentateur doit être sûr que le sujet répond le plus vite possible. Théoriquement le temps de réaction doit être enregistré lorsque le taux d’erreurs est nul ce qui est pratiquement impossible. Classiquement, les expérimentateurs acceptent un temps de réaction pour lequel ils enregistrent un taux d’erreurs inférieur à 5 % (Pachella, 1974).
Quant à l’intervalle inter-réponses (IIR), il représente la durée qui sépare les deux réponses R1 et R2. Généralement l’IIR = TR2 – TR1+ IIS. La valeur de l’IIR représente la quantité d’interférence entre les deux tâches T1 et T2 alors que le signe de l’IIR représente l’ordre dans lequel les deux réponses R1 et R2 sont émises. La distribution des IIR au cours des essais expérimentaux est souvent analysée. Selon que la distribution soit unimodale ou bimodale, elle renseigne sur l’ordre d’émission des réponses R1 et R2. Une distribution bimodale des IIR confirme le modèle de goulot d’étranglement. Dans ce cas, la distribution est large et signée montrant que R1 est émise avant R2 et/ ou l’inverse. Une distribution unimodale, peu variable et centrée autour de la valeur 0, montre que R1 et R2 sont groupées. Toutefois, cette caractéristique de la distribution ne permet pas à elle seule ni de confirmer ni de réfuter les deux modèles d’interférence cités précédemment.
Principaux facteurs manipulés.
Les trois principaux facteurs expérimentaux manipulés sont : l’instruction donnée aux sujets par consigne de l’expérimentateur, l’intervalle inter-stimuli (IIS) et la difficulté de la tâche.
Concernant les instructions, selon le modèle d’interférence étudié, l’expérimentateur accorde ou non une priorité à une des deux tâches proposées. Pour tester le modèle de goulot d’étranglement, les instructions données par l’expérimentateur accordent une priorité plus importante à la réalisation d’une des deux tâches, généralement, la première présentée. Ceci, en demandant aux sujets de répondre le plus rapidement possible à cette tâche (Pashler, 1984 ; Pashler & Johnston, 1989).
l’expérimentateur poussent le sujet à partager ses capacités, assignant une priorité égale aux deux tâches (Pashler, 1994b ; Ruthruff et al., 2003b). Ceci en laissant toute liberté aux sujets de répondre aux deux tâches dans l’ordre qu’ils veulent (Tombu & Jolicoeur, 2002) ou bien en exigeant de répondre aux deux tâches en même temps (Ruthruff, Pashler & Klaassen, 2001). Les études testant le modèle de goulot d’étranglement assignent souvent une priorité à une des deux tâches. Toutefois ce goulot d’étranglement peut refléter soit une limitation structurale inhérente à l’architecture cognitive soit une mise en attente stratégique volontaire induite par les instructions de l’expérimentateur pour valoriser une tâche par rapport à l’autre. Pour cette raison, les consignes d’absence de priorité entre les deux tâches ne permettent pas uniquement de tester si le partage des capacités a lieu mais aussi de comprendre la nature de ce goulot d’étranglement, qu’il soit structural ou stratégique (Ruthruff et al., 2001).
En ce qui concerne la manipulation de l’intervalle inter-stimuli (IIS), elle s’effectue à deux niveaux : la valeur absolue et le signe. La valeur de l’IIS représente la durée qui sépare la présentation des deux stimuli. Le signe de l’IIS représente l’ordre de présentation des deux stimuli. Le modèle de partage des capacités ainsi que celui du goulot d’étranglement ont chacun des prédictions différentes en ce qui concerne l’effet de la variation de la valeur et du signe de l’IIS sur le ralentissement de chacune des tâches T1 et T2, ainsi que sur la valeur et le signe de l’IIR. Le modèle de partage des capacités est confirmé si 1/ la réduction de la valeur de l’IIS augmente le ralentissement de T1 et T2 d’une façon égale (Tombu & Jolicoeur, 2002) et/ou, 2/ la variation de la valeur de l’IIS n’a aucun effet ni sur la valeur ni sur le signe de l’IIR. Le modèle de goulot d’étranglement est confirmé si 1/ la réduction de la valeur de l’IIS augmente le ralentissement de T2 mais pas celui de T1 (Pashler & Johnston, 1989) et/ou, 2/ la variation de la valeur de l’IIS n’a aucun effet sur la valeur de l’IIR mais le signe de l’IIS détermine strictement le signe de l’IIR (Pashler, 1994b ; Ruthruff et al., 2003b).
La difficulté de la tâche se fait, quant à elle, en manipulant l’intensité du stimulus et la compatibilité stimulus-réponse. La variation de la difficulté de la tâche révèle l’existence ou non d’un goulot d’étranglement ainsi que son locus. La difficulté de la tâche peut être variée par l’intensité du stimulus et/ou par la compatibilité stimulus-réponse. Chacun de ces facteurs agit sur une étape différente du traitement de l’information : l’intensité du stimulus agit sur les étapes perceptives alors que la compatibilité stimulus-réponse agit sur l’étape de sélection de la réponse. Les modèles de partage des capacités et celui du goulot d’étranglement ont chacun des prédictions