Caractérisation des transporteurs d’efflux de
silicium (Si) chez le soya
Mémoire
Julie Anne Wilkinson
Maîtrise en biologie végétale
Maître ès sciences (M.Sc.)
Québec, Canada
iii
RÉSUMÉ
Ces dernières années, l’intérêt pour l’accumulation de silicium (Si) chez les plantes a connu un essor considérable. Cet intérêt a été stimulé par la découverte des transporteurs d’influx et d’efflux de Si chez les plantes et par les nombreuses études qui rapportent les effets bénéfiques de son accumulation dans la lutte contre les stress. Récemment, la découverte d’un transporteur d’influx de Si chez le soya a permis de confirmer que l’espèce possède la capacité d’accumuler l’élément, alors qu’elle était initialement reconnue comme non-accumulatrice. L’identification subséquente de transporteurs d’efflux chez la même espèce (GmLsi2a, GmLsi2b, GmLsi2c) fait l’objet du présent mémoire. Le classement de 328 lignées de soya en neuf différents haplotypes selon le génotype rencontré chez les gènes GmLsi2b et GmLsi2c a permis de lier l’haplotype H1 à une accumulation accrue de Si chez le soya. Ce projet visait à vérifier si l’allèle spécifique de Lsi2 retrouvé chez de rares variétés de soya confère une absorption accrue de Si. Pour ce faire, nous avons utilisé un système hétérologue d'expression soit Arabidopsis thaliana, système déjà éprouvé pour le rôle de Lsi1 (Montpetit et al., 2012). L’accumulation de Si des différents allèles des gènes GmLsi2b et GmLsi2c ont été testés chez Arabidopsis. Les résultats obtenus suggèrent l’implication de l’haplotype H1 dans l’accumulation accrue de Si.
v
TABLE DES MATIÈRES
RÉSUMÉ ... iii
TABLE DES MATIÈRES ... v
LISTE DES TABLEAUX ... vii
LISTE DES FIGURES ... ix
REMERCIEMENTS ... xi
AVANT-PROPOS ... xiii
INTRODUCTION ... 1
HYPOTHÈSE ET OBJECTIF ... 3
CHAPITRE 1: REVUE DE LITTÉRATURE ... 5
1. Le silicium ... 7
1.1 Le silicium dans l’environnement ... 7
1.2 Le silicium dans les plantes ... 8
1.3 Effets bénéfiques ... 9
1.3.1 Effet sur les stress abiotiques ... 9
1.3.2 Effet sur les stress biotiques ... 9
1.4 Modes d’action ... 10
1.4.1 Hypothèse mécanique ... 10
1.4.2 Hypothèse du Si soluble ... 11
1.5 Les transporteurs de silicium ... 12
1.5.1 Les transporteurs d’influx ... 13
1.5.2 Les transporteurs d’efflux ... 15
1.5.3 Localisation et expression des transporteurs ... 16
2. Le Soya (Glycine max) ... 17
2.1 Découverte d’un transporteur d’influx ... 17
2.2 Découverte d’un transporteur d’efflux ... 19
2.3 Approche génétique ... 20
CHAPITRE 2: MANUSCRIT ... 23
CHARACTERISATION OF SILICON (SI) EFFLUX TRANSPORTERS IN SOYBEAN ... 25
RÉSUMÉ ... 25
ABSTRACT ... 26
INTRODUCTION ... 27
MATERIALS AND METHODS ... 29
Plant material and growth conditions ... 29
Isolation of promoter region, construction of promoters::GUS reporters and plant transformation ... 30
vi
Construction of plant expression vectors and plant transformation ... 31
Determination of Si concentration in transgenic Arabidopsis shoots ... 32
Statistical analyses ... 33
RESULTS ... 33
Analysis of promoter::GUS reporter constructs in Arabidopsis... 33
Cloning and characterization of the soybean Lsi2 genes ... 35
Controls ... 35
Promoters ... 36
GmLsi2 haplotypes ... 37
Analysis with the five best transformants ... 38
DISCUSSION ... 39
ACKNOWLEDGEMENTS ... 43
REFERENCES ... 44
CHAPITRE 3: CONCLUSIONS GÉNÉRALES ... 49
vii
LISTE DES TABLEAUX
CHAPITRE 1
TABLEAU 1.TRANSPORTEURS PUTATIFS DE SI IDENTIFIÉS DANS LE GÉNOME DE 17 ESPÈCES DE PLANTES SUR LES 25 ANALYSÉES.
... 15 TABLEAU 2.DIVERSITÉ DES HAPLOTYPES DÉTECTÉS AVEC DES SNPS SÉLECTIONNÉS DANS LES GÈNES GMLSI2B ET GMLSI2C À
PARTIR DE 328 LIGNÉES RESÉQUENCÉES PUBLIQUES ET PRIVÉES ... 20
CHAPITRE2
ix
LISTE DES FIGURES
CHAPITRE1
FIGURE 1.LOCALISATION DES TRANSPORTEURS LSI1 ET LSI2 ET VOIE EMPRUNTÉE PAR L’ACIDE SILICIQUE CHEZ LE RIZ DES RACINES JUSQU’AUX PARTIES AÉRIENNES ... 17
CHAPITRE2
FIGURE 1.GUS EXPRESSION DRIVEN BY THE CASP2 PROMOTER IN TRANSGENIC T2ARABIDOPSIS PLANTS ... 34 FIGURE 2.DIFFERENTIAL GUS EXPRESSION DRIVEN BY THE NIP5;1 PROMOTER IN TRANSGENIC T2ARABIDOPSIS PLANTS ... 35 FIGURE 3.SILICON (SI) ACCUMULATION IN SHOOTS OF ARABIDOPSIS CONTROL PLANTS GROWN IN THE ABSENCE (SI-) OR
PRESENCE (SI+) OF SOLUBLE SI ... 36 FIGURE 4.SILICON (SI) ACCUMULATION IN SHOOTS OF TRANSGENIC ARABIDOPSIS PLANTS HARBOURING THE GMLSI2B OR
GMLSI2C GENE WITH EITHER THE NIP5;1 OR THE CASP2 PROMOTER GROWN IN THE PRESENCE (SI+) OF SOLUBLE SI ... 37 FIGURE 5.SILICON (SI) ACCUMULATION IN SHOOTS OF TRANSGENIC ARABIDOPSIS PLANTS GROWN IN THE PRESENCE (SI+) OF
SOLUBLE SI WITH THE GMLSI2B GENE (A) OR GMLSI2C GENE (B) ... 38 FIGURE 6.SILICON (SI) ACCUMULATION IN SHOOTS OF FIVE TRANSGENIC ARABIDOPSIS LINES GROWN IN THE PRESENCE (SI+) OF SOLUBLE SI WITH THE GMLSI2B GENE (A) OR GMLSI2C GENE (B) ... 39
xi
REMERCIEMENTS
Je tiens tout d’abord à remercier mon directeur de recherche, M. Richard Bélanger, de m’avoir accueillie au laboratoire il y a maintenant trois ans. Merci pour la confiance que vous m’avez accordée dès le départ et pour m’avoir guidé par la suite tout au long de mon projet. J’ai hérité du projet un peu plus casse-gueule mais rempli de beaux défis! Je me serai creusé les méninges plus d’une fois afin de surmonter chaque étape, chaque protocole, ou encore chaque kit qui me donnait du fil à retordre. Plusieurs instants de petites frustrations sans lesquelles je n’aurais pas autant appris, ni même autant apprécié mon projet.
Merci aussi à M. François Belzile pour tous les précieux conseils lors des réunions du vendredi. Toujours une question pertinente sur un aspect auquel on n’a pas réfléchi, mais ce sont ces petites remises en questions qui nous permettent d’avancer.
Mes remerciements vont ensuite bien évidemment à Caroline Labbé, pour ta patience malgré mes allées et venues dans ton bureau. Et aussi pour ces après-midis à réfléchir, mais aussi à discuter de bien d’autres sujets! Un grand merci aussi à Julien Vivancos, qui m’a initié à la biologie moléculaire, moi qui étais encore une novice à mon arrivée.
Bastien, François, Maxime, Sarah, Marc-Olivier, Stéphanie, Aliyeh, Geneviève, Hicham, Marie-Hélène, Samuel, Gowsica, Joan, Rupesh et Huma, merci infiniment pour votre présence. Un petit clin d’oeil à Amandine avec laquelle j’ai cheminé tout doucement, au gré de nos discussions et encouragements mutuels.
Finalement, je tiens à remercier ma famille, particulièrement ma mère Clémence et mon conjoint Mathieu qui m’ont épaulée et soutenue dans tout ce processus. Je remercie aussi ma cousine Anne pour les séances de sport hebdomadaires qui changent les idées et qui me permettent de rester zen, no matter what!
xiii
AVANT-PROPOS
Ce mémoire comprend une revue de littérature qui fait état des connaissances actuelles en lien avec l’accumulation de silicium (Si) chez les plantes et son rôle dans la protection contre les stress biotiques et abiotiques. On y détaille aussi la découverte des transporteurs d’influx et d’efflux de Si qui permettent le transport du Si chez les plantes, et leur présence dans le règne végétal. La seconde partie du premier chapitre est dédiée à la découverte de transporteurs de Si chez le soya, une plante initialement classée comme non-accumulatrice. L’observation d’une accumulation différentielle de Si entre différents cultivars testés a donné lieu à une première série d’expériences au terme de laquelle il n’a pas été possible de lier le Lsi1 identifié à l’accumulation différentielle observée. Finalement, on insiste sur la découverte de trois transporteurs d’efflux (GmLsi2a, GmLsi2b, GmLsi2c) chez le soya. Deux de ces transporteurs (GmLsi2b et GmLsi2c) sont possiblement impliqués dans l’accumulation différentielle et font l’objet du présent mémoire.
Le second chapitre de ce mémoire dont je suis l’auteure principale traite de la caractérisation des transporteurs d’efflux (GmLsi2b et GmLsi2c) découverts chez le soya et contient l’ensemble de mes travaux de maîtrise. La découverte de ces transporteurs revient à M. Rupesh Deshmukh de même que les expériences réalisées en amont de mon projet. Les lignées transgéniques TaLsi1 d’Arabidopsis utilisées ont été générées par Jonatan Montpetit dans le cadre de son projet de maîtrise.
Le troisième et dernier chapitre de ce mémoire évalue la pertinence des essais réalisés et permet de les inscrire dans un cadre plus global.
Ce projet a été réalisé en partenariat avec la société Syngenta, entreprise spécialisée dans la vente de semences et produits phytosanitaires. Les résultats présentés sont inclus dans le dépôt d’un brevet avec la compagnie Syngenta.
1
INTRODUCTION
La culture du soya (Glycine max (L.) Merr.) est l’un des piliers économique de l’agriculture en Amérique du Nord. Tout près de 115 millions de tonnes y étaient produites en 2014, ce qui la classe au second rang en terme de production après la culture du maïs (FAOSTAT 2015). L’espèce appartient à la famille des légumineuses et permet la fixation de l’azote atmosphérique (N2) lors de la symbiose avec les bactéries du type Rhizobium. Cette
particularité en fait une espèce de choix dans une rotation, tout particulièrement avec le maïs (Zea mays L.), espèce très gourmande en azote. Aussi, la culture du soya a connu un essor considérable au cours des dernières décennies en raison de la haute teneur en protéines (40 %) et en huile (20 %) de ses fèves, que ce soit pour l’alimentation animale ou humaine.
L’intensification des productions agricoles est le principal moyen de satisfaire la demande croissante en nourriture qui découle d’une population mondiale sans cesse grandissante. Toutes les cultures sont toutefois exposées à différents stress et on peut facilement imaginer que les pertes se chiffrent en milliards de dollars chaque année. Le soya ne fait pas exception et sa production est affectée par de nombreux facteurs biotiques et abiotiques. Les facteurs abiotiques que l’on rencontre le plus souvent sous nos conditions sont les faibles températures et le gel. Les facteurs biotiques, incluent notamment les insectes, les mauvaises herbes et les maladies, ces deux derniers étant les principaux responsables des pertes de rendement rencontrées (Agriculture et Agroalimentaire Canada, 2006). Liu et al. (2015) estiment que les pertes causées par les pathogènes du soya s’élèvent à 4 milliards de dollars annuellement, et ce uniquement aux États-Unis. Il est ainsi très important que les efforts investis en recherche soient orientés vers la lutte contre les stress biotiques et abiotiques afin de limiter les pertes de rendement chez les producteurs.
L’importance du soya au niveau économique l’a doté d’un certain statut en recherche, d’où son génome bien annoté. Au fil des ans, des efforts considérables ont été investis afin de caractériser la diversité génétique des génotypes de soya sauvages et cultivés. À ce jour, plusieurs cultivars de soya qui présentent des phénotypes intéressants pour l’amélioration
2
génétique ont été étudiés et la recherche a été orientée principalement vers la résistance contre les stress biotiques et abiotiques puisqu’ils sont les principaux responsables des pertes économiques. Ces dernières années, un phénotype particulier a attiré l’attention en recherche : l’accumulation de silicium (Si). L’intérêt grandissant pour cet élément a été stimulé par la découverte des transporteurs d’influx et d’efflux de Si chez les plantes (Ma et al., 2006 ; 2007a) et par la multitude d’études qui rapportent les effets bénéfiques de son accumulation dans la lutte contre divers stress, notamment les maladies, les insectes et la sècheresse (Liang et al., 2015). Un transporteur d’influx (Lsi1) a été récemment identifié chez le soya, ce qui a permis de confirmer la capacité de l’espèce à accumuler l’élément, alors qu’elle était initialement reconnue comme non-accumulatrice (Deshmukh et al., 2013). Cette découverte a aussi permis d’expliquer les effets bénéfiques d’un apport en Si rapportés dans la littérature. En effet, les chercheurs avait remarqué ces dernières années qu’il est possible d’obtenir une résistance accrue contre la rouille du soya (Phakopsora pachyrhizi) (Arsenault-Labrecque et al., 2012), ou alors lors d’une infection par la pourriture phytophthoréenne (Phytophtora sojae) (Guérin et al., 2014) après que les plantes aient reçu un apport en Si. Chez le soya, on rencontre une grande variabilité pour l’accumulation de Si entre les cultivars tel que montré par Arsenault-Labrecque et al. (2012) et Deshmukh et al. (2013), et le transporteur d’influx identifié initialement ne permet pas d’expliquer la différence observée. C’est pourquoi les chercheurs se sont alors penché sur l’implication éventuelle d’un Lsi2. La découverte de trois transporteurs d’efflux (GmLsi2a, GmLsi2b et GmLsi2c) a ouvert la voie à de nouvelles hypothèses de recherche et a permis le classement de 328 lignées de soya en neuf haplotypes différents selon le génotype rencontré pour les gènes GmLsi2b et GmLsi2c (données non publiées). Cette classification a lié un seul haplotype (H1) à une accumulation accrue de Si chez le soya et sa présence est restreinte à de rares cultivars. Inversement, l’haplotype H5 est celui présent chez le plus grand nombre de cultivars (305 sur un total de 328), dont celui de référence (Williams 82) et est lié à une faible accumulation de Si.
3
HYPOTHÈSE ET OBJECTIF
Ce projet s’inscrit dans une démarche globale qui vise à caractériser les transporteurs de Si chez les plantes afin d’approfondir nos connaissances à propos d’un élément dont la complexité n’a d’égal que les bénéfices qui lui sont associés. Plus précisément, nous avons participé en partie à la caractérisation d’un transporteur d’efflux (Lsi2) de Si récemment identifié chez le soya. Pour ce faire, nous avons utilisé un système hétérologue d'expression dans Arabidopsis thaliana, système déjà éprouvé pour le rôle de Lsi1 au laboratoire. Nous avons émis l’hypothèse selon laquelle un allèle spécifique de Lsi2 retrouvé chez de rares variétés de soya confère une absorption accrue de Si. Pour y parvenir, l’objectif était de démontrer si la variation allélique des gènes GmLsi2b ou GmLsi2c présents chez l’haplotype H1 est responsable de la capacité différentielle d’absorption de Si rencontrée chez le soya.
CHAPITRE 1
7
1. LE SILICIUM
1.1 Le silicium dans l’environnement
Le silicium (Si) constitue 28 % du poids de la lithosphère (Liang et al., 2015), ce qui le positionne en seconde place au niveau de l’abondance (Epstein, 1999). Dans le sol, le Si se présente le plus souvent sous sa forme la plus simple et très faiblement soluble, la silice amorphe (SiO2) ou alors sous forme de silicates (Si-O-métal) (Sommer et al., 2006 ; Řezanka
et Sigler, 2008). L’élément n’est pas considéré comme essentiel puisque les plantes peuvent compléter leur cycle vital en son absence (Sachs, 1865 ; Epstein, 1999). Il n’en reste pas moins un constituant inorganique important malgré sa quasi-absence dans les publications scientifiques. La tendance s’inverse depuis le début des années 2000, et l’intérêt pour l’élément ne cesse de croitre. Le nombre de publications dont il fait l’objet le démontre, tout particulièrement celles qui rapportent les effets bénéfiques rencontrés chez les plantes. Afin d’être utilisé par la plante, tout élément doit être absorbé par celle-ci, et donc se retrouver sous une forme assimilable. Dans la solution du sol, seule une fraction infime de la silice se trouve sous forme soluble, soit l’acide silicique (Si(OH)4) (Epstein, 1999). Sa concentration varie entre 0,1 et 0,6 mM, ce qui est semblable à celle du calcium (Ca), du potassium (K) de même que celle de certains autres éléments majeurs, tout en étant nettement supérieure à celle du phosphore (P) (Epstein, 1994). La concentration en Si dans la solution du sol est déterminée par la nature de la roche-mère, la température, la taille des particules, le temps d’écoulement de l’eau interstitielle et le pH (Sommer et al., 2006). En résumé, le Si est présent dans le sol en grande quantité sous diverses formes dont seule une faible partie est disponible pour les plantes.
8
1.2 Le silicium dans les plantes
Malgré son omniprésence dans les sols, les plantes ne sont pas égales quant à leur capacité à accumuler le Si. Notons que toute plante cultivée contient une certaine quantité de Si, quantité pour laquelle il existe une grande variation. On retrouve dans les tissus d’une plante entre 0,1 et 10 % de Si sur une base de matière sèche (M.S.) (Epstein, 1994), variation qui a été attribuée à la capacité ou l’incapacité des racines à absorber l’acide silicique (Takahashi et al., 1990). À pH physiologique, ce dernier se polymérise en silice amorphe (SiO2 nH2O) lorsque la concentration excède 1,7 mM. La polymérisation étant un processus irréversible, il est impossible pour le Si de retourner sous forme soluble une fois déposé en silice amorphe dans la plante (Samuels et al., 1991). L’accumulation de Si est un processus passif ou actif selon l’espèce (Liang et al., 2015), et les plantes ont été divisées en deux groupes selon la quantité retrouvée dans les parties aériennes. On a les plantes dites accumulatrices chez lesquelles on retrouve plus de 1 % Si M.S. et les autres sont classées comme non accumulatrices (Deshmukh et Bélanger, 2015). Les accumulatrices sont principalement des monocotylédones comme le riz (Oryza sativa) et le blé (Triticum aestivum) (Epstein, 1999) au même titre que certaines espèces primitives telle que la prêle (Equisetum arvense) (Grégoire et al., 2012). Jusqu’à tout récemment, les dicotylédones étaient essentiellement classées du côté des non accumulatrices, avec pour seule exception les Cucurbitaceae (Hodson et al., 2005).
Avec le développement des nouvelles technologies de séquençage, la publication de génomes a connu un essor considérable. Une étude récente a analysé le génome de 25 espèces séquencées et a fourni les bases moléculaires précises permettant la classification des espèces végétales (voir section 1.5.1). Certaines dicotylédones dont le soya (Glycine max) font maintenant partie du groupe des plantes accumulatrices (Deshmukh et al., 2015). L’intérêt croissant pour l’élément vient principalement de son rôle bénéfique sur la santé de la plante, et ce tout particulièrement en situation de stress (Fauteux et al., 2005).
9
1.3 Effets bénéfiques
En milieu naturel, les plantes sont constamment exposées à des conditions adverses et plusieurs travaux ont montré qu’un apport en Si permet d’accroitre la résistance face aux stress biotiques et soutient une meilleure croissance en présence de stress abiotiques (Epstein, 1999). Chez les plantes, la croissance est fortement liée aux différents éléments du sol, qu’ils soient essentiels, toxiques ou bénéfiques. Les éléments toxiques affectent négativement la croissance de la plante alors que les éléments essentiels jouent un rôle crucial dans son développement. De leur côté, les éléments bénéfiques stimulent la croissance des plantes et peuvent s’avérer essentiels pour certaines espèces ou sous certaines conditions de croissance (Sahebi et al., 2015; Broadley et al., 2012). Le Si est un élément bénéfique et on dit de lui que son effet est prophylactique, c’est-à-dire qu’il s’utilise en traitement préventif. Autrement dit, il doit y avoir accumulation de Si avant l’exposition à un stress afin d’observer des effets bénéfiques (Epstein et Bloom, 2005).
1.3.1 Effet sur les stress abiotiques
De nombreuses recherches ont montré l’effet du Si sur l’atténuation des symptômes face aux stress abiotiques. Avec l’avènement des changements climatiques, une pression de plus en plus forte de la plupart des stress est à prévoir, notamment la sècheresse et la salinité. Selon Zhu et Gong (2014), l’application de Si dans de telles situations permet de réduire considérablement les effets négatifs sur les cultures. Aussi, le Si atténue le stress causé par certains éléments nutritifs (Fe, Mn) et métaux lourds (Al, Cd) (Liang et al., 2007) et peut également minimiser les effets d’une carence ou d’un excès des éléments majeurs, notamment l’azote et le phosphore (Ma et al., 2001). Finalement, les effets bénéfiques d’une application de Si sur la réduction de stress tels que les radiations, les températures extrêmes, le vent et la lumière excessive ont aussi été observés (Ma et al., 2004).
1.3.2 Effet sur les stress biotiques
L’effet bénéfique de l’accumulation de Si sur la réduction des stress biotiques a été observé à maintes reprises et de nombreux articles portent sur le sujet. Les recherches ont été orientées en partie sur son effet contre les maladies d’origine fongique et plusieurs
10
pathosystèmes ont été étudiés dont celui du concombre (Cucumis sativus) infecté par Pythium ultimum ou Sphaerotheca fuliginea (Chérif et al., 1992; Fawe et al., 1998), celui du blé infecté par Blumeria graminis (Bélanger et al., 2003) ou encore ceux du riz infecté par divers agents pathogènes comme Magnaporthe grisea et Bipolaris oryzae (Rodrigues et al., 2004; Dallagnol et al., 2011).
De nombreux chercheurs ont aussi observé que le Si confère un avantage aux plantes dans la lutte contre les insectes phytophages. Une revue de littérature effectuée par Reynolds et al. (2009) fait état des principales espèces végétales chez lesquelles le Si permet de réduire l’incidence des insectes piqueurs et broyeurs. On retrouve entre autres le riz, le maïs (Zea mays), la canne à sucre (Saccharum officinarum), le blé et l’orge (Hordeum vulgare). De plus, deux études récentes ont montré qu’un apport en Si chez le riz permet de réduire l’incidence de deux insectes, la pyrale des herbes (Cnaphalocrocis medinalis) et la cicadelle brune (Nilaparvata lugens) (Ye et al., 2013 ; He et al., 2015). À noter que Massey et al. (2006) rapportent que l’ajout de Si n’aurait aucun effet en présence d’insectes suceurs de phloème. De façon générale, le Si aurait un effet sur la diminution de la digestibilité, l’augmentation de l’abrasivité et de la dureté des tissus de la plante (Reynolds et al., 2009). À la lumière des observations rapportées dans la littérature, il semble évident que le Si protège la plante contre divers stress, qu’ils soient biotiques ou abiotiques. La question que l’on se pose alors est : Quels sont les mécanismes impliqués? Une question simple à priori, mais qui est encore aujourd’hui le sujet de nombreux débats et réflexions. La section suivante rapporte les connaissances actuelles, quoique encore incomplètes sur les modes d’action.
1.4 Modes d’action
1.4.1 Hypothèse mécaniqueLa première hypothèse énoncée pour expliquer les effets bénéfiques du Si est désignée sous le nom d’« hypothèse mécanique » (Jones et Handreck, 1967). Une fois l’acide silicique absorbé par les racines, il y a polymérisation dans les parties aériennes sous forme de silice
11
amorphe (non soluble) couramment désignée sous le nom d’opale ou phytolithe (Bauer et al., 2011). L’hypothèse propose que le Si non soluble déposé agit en tant que barrière physique (Jones et Handreck, 1967) en améliorant la rigidité et la force des tissus, mais aussi en entravant la pénétration par les agents pathogènes fongiques et les insectes (Ma et al., 2015).
1.4.2 Hypothèse du Si soluble
L’hypothèse de la barrière physique ne peut expliquer à elle seule la protection conférée par le Si, puisque l’on observe des changements métaboliques (induction des réponses de défense) suite à son application (Fauteux et al., 2005). Les fondements de l’hypothèse ont été rapidement ébranlés, principalement suite à des observations contradictoires effectuées entre autres par Samuels et al. (1991) et Chérif et al. (1992). Ces observations ont aidé à la formulation d’une nouvelle hypothèse, soit celle du « Si soluble ». Cette dernière propose que la forme soluble du Si agit sur le métabolisme de la plante en tant que modulateur des réactions de défense (Fawe et al., 2001 ; Fauteux et al. 2005).
Les recherches ont dès lors été orientées vers l’influence du Si sur l’induction des mécanismes de défense de la plante. Plusieurs études ont montré à l’aide de différents pathosystèmes que les plantes ayant reçu un apport en Si, comme le concombre, le blé ou le riz, vont résister plus efficacement à l’attaque d’un agent pathogène (Fauteux et al., 2005). Les mécanismes de défense sont alors activés par la production entre autres de protéines de défense, de composés phénoliques et de phytoalexines (Chérif et al., 1994 ; Fawe et al., 1998 ; Rémus-Borel et al., 2005). La transcriptomique a par la suite permis d’analyser sous un tout autre angle l’interaction hôte/agent pathogène. Les pathosystèmes comme Arabidopsis thaliana/blanc (E. cichoracearum) (Fauteux et al., 2006) ou celui blé/blanc poudreux (B. graminis) (Chain et al., 2009) traités avec du Si ont tous deux permis de mettre en évidence l’induction de gènes liés aux mécanismes de défense des plantes en situation de stress. Plus récemment, le mécanisme de « priming » des réponses de défense chez les insectes a été lié avec le Si (Ye et al., 2013).
12
Le « priming » se définit comme la sensibilisation d’une cellule ou d’un organisme menant à une réaction de défense décuplée (Conrath et al., 2015). En situation de stress, l’activation des réponses de défense est ainsi plus rapide et vigoureuse. Les processus liés au « priming» incluent notamment la résistance systémique acquise qui requiert la synthèse d’acide salicylique (Conrath et al., 2015). Récemment, une étude a été réalisée par Vivancos et al. (2015) à l’aide de la plante modèle Arabidopsis thaliana infectée par le blanc (E. cichoracearum). Les résultats obtenus indiquent que le Si contribue au mécanisme de « priming » même lorsque celui-ci est altéré. La nouvelle hypothèse énoncée veut que le Si participe à la formation d’une membrane physique pour ralentir les effecteurs en plus d’intervenir dans le mécanisme de « priming » (Vivancos et al., 2015). Cette hypothèse combine ainsi les deux hypothèses, et y ajoute la dimension de l’interaction avec les effecteurs.
Même si on ne comprend pas encore tous les mécanismes liés au mode d’action du Si dans la plante, la fertilisation des rizières du Japon inclut l’ajout de Si depuis le début des années 50 (Liang et al., 2015). Son utilisation est maintenant courante dans les rizières du Brésil, de la Chine et des États-Unis (Ma et al., 2015), de même que dans la culture de la canne à sucre (Savant et al., 1999). Rappelons cependant que certaines cultures ne semblent tirer aucun bénéfice d’un tel apport, ce qui a laissé les scientifiques perplexes pendant de nombreuses années. Tel que mentionné, ce ne sont pas toutes les plantes qui ont la capacité d’accumuler du Si et les mécanismes qui permettent l’accumulation, soit la présence de transporteurs, n’ont été découverts que récemment (Ma et al., 2006 ; 2007a). Le développement de pratiques culturales favorisant une agriculture durable est fonction de la compréhension de ces mécanismes, puisque l’application de Si pourrait permettre de réduire celle d’intrants chimiques.
1.5 Les transporteurs de silicium
Il a été établi qu’il existe une certaine corrélation entre la quantité de Si absorbée et les effets bénéfiques qui lui sont associés. Ainsi, ce sont les plantes dites accumulatrices qui tirent le plus grand bénéfice de l’effet protecteur du Si. Les travaux de Ma et al. (2001) ont
13
permis de mettre en évidence que le transport se fait de façon active chez certaines plantes, notamment le riz, puisque le flux de transpiration n’explique pas en totalité son accumulation. On sait aussi que pour observer des effets bénéfiques, le Si doit être absorbé par les racines, l’application foliaire étant nettement moins efficace (Guével et al., 2007). De façon générale, les transporteurs racinaires impliqués dans l’absorption de la plupart des éléments nutritifs sont bien caractérisés (White, 2012), mais ce n’est que récemment que les transporteurs d’acide silicique ont été identifiés chez les plantes. La variabilité importante dans l’absorption de Si que l’on rencontre au sein du règne végétal de même que l’observation des effets bénéfiques pourraient être expliqués en très grande partie par la présence ou l’absence de tels transporteurs.
1.5.1 Les transporteurs d’influx
Le premier transporteur d’acide silicique dans le règne végétal a été identifié en 2006 par Ma et al. et constitue une percée scientifique majeure. C’est suite au criblage génétique sur des lignées mutantes qui accumulent très peu de Si que les chercheurs ont isolé le premier transporteur, nommé Low silicon rice 1 (Lsi1). Il s’agit d’un transporteur de type canal qui facilite le transport passif à travers la membrane plasmique entre la solution externe (par exemple l’apoplaste) et les cellules de la plante (Ma et al., 2006). Il existe également un homologue très proche chez le riz nommé Lsi6 (Ma et al., 2006), responsable du transport de l’acide silicique hors du xylème, d’où son influence sur la distribution du Si dans les parties aériennes (Yamaji et Ma, 2009).
Les transporteurs de type Lsi1 appartiennent à la famille des Major Intrinsic Proteins (MIPs), des canaux transmembranaires qui facilitent et discriminent le transport de l’eau et de différents solutés à travers la membrane. Puisque la grande majorité des MIP retrouvées chez les plantes servent au transport de l’eau, on les appelle communément aquaporines (Maurel et al., 2008). Il existe cinq classes différentes de MIP dont les Nodulin-26-like Intrinsic membrane Proteins (NIP) qui permettent, outre le passage de l’acide silicique, celui de l’acide borique, de l’urée, du glycérol, de l’ammoniaque et de l’arsénite (Bienert et Chaumont, 2011). La découverte des transporteurs d’acide silicique a permis la création
14
d’un nouveau groupe, les NIPIII dont le filtre ar/R est composé des quatre acides aminés : GSGR (Mitani et al., 2008). Ils possèdent une structure caractéristique qui consiste en six domaines transmembranaires et leur spécificité est définie par le motif NPA (Asn-Pro-Ala) ainsi que par les quatre acides aminés (GSGR) qui forment le filtre de sélectivité aromatique/arginine (ar/R) (Mitani et al., 2008). Mitani-Ueno et al. (2011) ont mis en évidence la spécificité des transporteurs d’influx lors d’une étude de mutagénèse dirigée chez la citrouille en montrant que la substitution des résidus GSGR permet d’empêcher le passage de l’acide silicique. Cependant, chez Arabidopsis, la substitution des résidus AIGR du filtre ar/R en résidus GSGR, n’a pas permis d’augmenter la capacité d’absorption de Si. Cela indique que des facteurs autres sont impliqués dans le processus d’absorption. En effet, certaines plantes comme la tomate (Solanum lycopersicum) possèdent les caractéristiques énoncées précédemment, sans toutefois posséder la capacité d’absorber du Si (Mitani et Ma, 2005). Ce n’est que récemment qu’une étude a montré qu’en plus de la présence d’une protéine NIPIII avec un filtre ar/R composé des résidus GSGR, l’accumulation de Si se limite aux espèces possédant une distance précise de 108 acides aminés entre les deux motifs NPA (Deshmukh et al., 2015).
Puisque le riz n’est pas la seule espèce capable d’accumuler du Si, les chercheurs ont par la suite vérifié si la présence de transporteurs s’étendait à d’autres espèces. Au fil des ans, plusieurs ont été caractérisés notamment chez l’orge (Chiba et al., 2009), le maïs (Mitani et al., 2009b), la citrouille (Cucurbita moschata) (Mitani et al., 2011), le blé (Montpetit et al., 2012), et plus récemment le soya (Deshmukh et al., 2013). Un article publié il y a quelques mois dans lequel le génome de 25 espèces séquencées a été analysé a permis de prédire avec certitude 30 transporteurs d’influx de Si chez 17 espèces, tel que présenté dans le tableau 1 (Deshmukh et al., 2015).
15
Tableau 1. Transporteurs putatifs d’influx de Si identifiés dans le génome de 17 espèces de plantes sur les 25 analysées. Les transporteurs de Si ont été identifiés à partir de 985 aquaporines sur la base de la présence de deux domaines NPA, des filtres de sélectivité GSGR et de différents résidus Froger’s
Traduit de Deshmukh et al., 2015 1.5.2 Les transporteurs d’efflux
La découverte d’un transporteur d’influx chez le riz a été rapidement suivie de celle du Low silicon rice 2 (Lsi2) (Ma et al, 2007a). Chez les plantes, il est responsable du transport de l’acide silicique à l’extérieur des cellules. Appartenant à une famille très peu caractérisée de transporteurs putatifs d’anions avec 11 domaines transmembranaires prédits, il ne montre aucune similarité avec les transporteurs de type Lsi1 et semble davantage en partager avec les transporteurs d’efflux d’arsénite identifiés chez les bactéries et archées (Ma et al., 2007a). Ma et al. en 2008 ont en effet montré qu’il est impliqué dans le transport de l’arsénite chez le riz. Contrairement au mécanisme de transport passif des Lsi1, le transport d’efflux de l’acide silicique par le Lsi2 est un processus actif dirigé par le gradient de proton à travers la membrane plasmique (Ma et al., 2015). On dispose de relativement peu d’information concernant les Lsi2 et ils ont été caractérisés chez peu d’espèces. On les
Espèce Identification génique (Résidus
Froger’s, distance NPA-NPA)
Accumulateur de Si (référence)
Brachypodium distachyon BdNIP2-1 (LTAYF, 108), BdNIP2-2 (LTAYF, 108) Oui (Présente étude)
Cajanus cajan CcNIP2-1 (FTAYF, 108), CcNIP2-2 (LTAYL, 108) Oui (Hodson et al., 2005)
Carica papaya CpNIP2-1 (LSAYL, 108) ?
Citrus sinensis CSNIP2-1 (LTAYL, 43) ?
Elaeis guineensis EgNIP2-1 (LTAYL, 108) Oui (Gowda et al., 2004)
Fragaria vesca FvNIP2-4 (LTAYM, 108), FvNIP2-5 (LTAYV, 108) Oui (Kanto et al., 2004)
Glycine max GmNIP2-1 (LTAYF, 108), GmNIP2-2 (LTAYF, 108) Oui (Présente étude)
Musa acuminate MaNIP2-1 (LTAYF, 108), MaNIP2-2 (LTAYL, 108),
MaNIP2-3 (LTAYF, 108), MaNIP2-4 (LTAYF, 108) Oui (Henriet et al., 2006)
Oryza sativa OsNIP2-1 (ITAYF, 108), OsNIP2-2 (LTAYF, 108) Oui (Ma et al., 2006)
Prunus persica PpeNIP2-1 (LTAYV, 108) Oui (Le Blond et al., 2011)
Populus trichocarpa PtNIP2-1 (LTAYL, 108) Oui (Présente étude)
Ricinus communis RcNIP2-1 (LTAYI, 108) ?
Sorghum bicolor SbNIP2-1 (LTAYF, 108), SbNIP2-2 (LTAYF, 108) Oui (Lux et al., 2002)
Setaria italic SiNIP2-1 (LTAYF, 108), SiNIP2-2 (LTAYF, 108) Oui (Weichenthal et al., 2003)
Solanum lycopersicum SlNIP2-1 (LSAYI, 109) Non (Présente étude)
Vitis vinifera VvNIP2-1 (LTAYA, 108) Oui (Blaich et Grundhofer, 1997)
Zea mays ZmNIP2-1 (LTAYF, 108), ZmNIP2-2 (LTAYF, 108),
16
retrouve chez le riz (Ma et al., 2007a), mais aussi chez le maïs, l’orge (Mitani et al., 2009a) et la citrouille (Mitani-Ueno et al., 2011).
1.5.3 Localisation et expression des transporteurs
Les transporteurs montrent des patrons d’expression différents selon l’espèce végétale dont il est question. Chez le riz (Ma et al., 2006), l’orge (Chiba et al., 2009), le maïs (Mitani et al., 2009b) et le blé (Montpetit et al., 2012), le Lsi1 s’exprime principalement dans les racines, alors que chez la citrouille (Mitani et al., 2011) et le soya (Deshmukh et al., 2013), il s’exprime aussi dans les feuilles, tout comme les Lsi6 caractérisés (Yamaji et Ma, 2009; Yamaji et al., 2012; Mitani et al., 2009b). Le Lsi2 s’exprime dans les racines chez le riz (Ma et al., 2007a), l’orge et le maïs (Mitani et al., 2009a) alors que les homologues chez la citrouille (Mitani-Ueno et al., 2011) s’expriment dans les racines et les parties aériennes. Les protéines du Lsi1 et Lsi2 sont toutes deux localisées dans la membrane plasmique, mais s’expriment dans différents type de tissus et de cellules selon l’espèce végétale (Ma et al., 2015). Par exemple, les Lsi1 du maïs et de l’orge sont localisés de façon polaire dans les cellules épidermales, hypodermales et corticales (Chiba et al., 2009; Mitani et al., 2009b), alors que les Lsi2 de l’orge et du maïs sont uniquement localisés dans l’endoderme des racines (Mitani et al., 2009a). La localisation cellulaire ou tissulaire des Lsi1 et Lsi2 détermine en partie leur rôle dans le transport du Si et dans la capacité à accumuler du Si. La figure 1 montre la localisation des transporteurs chez le riz, de même que la voie empruntée par l’acide silicique suite à son entrée dans la plante. La présence des deux transporteurs (Lsi1 et Lsi2) est essentielle à l’accumulation de Si. Le « knock-out » de l’un ou l’autre entraine une diminution dans l’accumulation de Si (Ma et al., 2006 ; 2007a).
17
Figure 1 : Localisation des transporteurs Lsi1 et Lsi2 et voie empruntée par l’acide silicique chez le
riz des racines jusqu’aux parties aériennes (schéma : R.R. Bélanger et C. Labbé)
2. LE SOYA (GLYCINE MAX)
2.1 Découverte d’un transporteur d’influx
Jusqu’à tout récemment, le seul transporteur d’influx connu chez les dicotylédones était celui caractérisé chez la citrouille (CmLsi1) (Mitani-Ueno et al., 2011). Par conséquent, l’information dont on disposait quant à la dynamique fonctionnelle, structurelle et évolutive concernant cette famille de gènes était plutôt limitée chez les dicotylédones en général. Ce n’est que récemment qu’un second transporteur d’influx a été identifié, cette fois chez le soya (Deshmukh et al., 2013). Initialement reconnue comme une espèce non-accumulatrice de Si, très peu d’attention a été portée au potentiel de cette plante à accumuler l’élément. L’identification de deux transporteurs d’influx (Lsi1) chez le soya, le premier cas chez les légumineuses, a confirmé la capacité de cette plante à absorber le Si, permettant ainsi de justifier les effets bénéfiques observés dans la littérature. En effet, des études récentes ont
18
montré l’effet bénéfique d’une accumulation de Si chez le soya en situation de stress radiatif (UV-B) (Shen et al., 2010), ou encore suite à l’attaque par un agent pathogène (Arsenault-Labrecque et al., 2012 ; Guérin et al., 2014). Une recherche par homologie de séquence a mené à l’identification de deux gènes Lsi1; GmNIP2-1 et GmNIP2-2 s’exprimant tous deux dans les racines et les parties aériennes du soya. L’expression du GmNIP2-2 dans les ovocytes de Xenopus laevis, système d’expression hétérologue fiable pour tester la fonction des transporteurs de Si (Miller et Zhou, 2000), a confirmé la capacité génétique du soya à absorber le Si (Deshmukh et al., 2013). Les caractéristiques inhérentes au transport du Si que l’on retrouve chez les autres transporteurs d’influx caractérisés sont présentes, soit les deux domaines NPA séparés par une distance de 108 acides aminés (Deshmukh et al., 2015) de même que le filtre de sélectivité ar/R formé par les résidus GSGR (Deshmukh et al., 2013). Lors de l’étude, trois cultivars différents ont été utilisés soit Hikmok sorip, Williams 82 et Jack. Fait intéressant, les cultivars testés ont montré une variation importante quant à leur capacité à accumuler le Si. On sait déjà qu’il existe une grande variation interspécifique dans l’accumulation de Si, par contre l’étude de la relation intraspécifique s’est faite à plus petite échelle et est généralement reconnue comme étant plus faible (Hodson et al., 2005). Jusqu’à présent, la variation intraspécifique a été documentée chez quelques espèces notamment le riz (Ma et al., 2007b), l’orge (Ma et al., 2003) et le soya (Deshmukh et al., 2013) et les différences observées demeurent largement inexpliquées. La présence de Lsi1 a été confirmée chez ces trois espèces, et la structure moléculaire semble être conservée au sein d’une même espèce, ce qui écarte l’hypothèse d’une variation génotypique du Lsi1 afin d’expliquer la différence observée. Chez le riz, les différentes concentrations chez deux variétés ont été attribuées au niveau d’expression des gènes Lsi1 et Lsi2 (Ma et al., 2007b). Dans Deshmukh et al. (2013), la différence observée entre le cultivar Hikmok sorip et Williams 82 est plus du double, ce qui a incité les chercheurs à approfondir les recherches. Lors de la comparaison des séquences des gènes GmNIP2-2 des trois cultivars, aucune différence n’a été observée, éliminant l’hypothèse d’une divergence au niveau de la séquence protéique. De façon similaire, le niveau d’expression du gène GmNIP2-2 entre les trois cultivars étant identique, il ne pouvait ainsi être responsable de la variation au niveau de l’absorption (Deshmukh et al., 2013). Puisque la variation observée n’est imputable ni à
19
la structure, ni au niveau d’expression génique, l’hypothèse voulant que la disparité dans l’accumulation de Si soit attribuable à un transporteur d’efflux (Lsi2) a été émise.
2.2 Découverte d’un transporteur d’efflux
Après des recherches non concluantes avec le Lsi1 du soya, l’équipe du Dr Bélanger du Laboratoire de phytopathologie de l’Université Laval a décidé d’approfondir les recherches au niveau d’un transporteur d’efflux (Lsi2) chez le soya. Une des stratégies mises en œuvre pour y parvenir a été le développement d’une population de RIL (Recombinant Inbred Lines) entre les cultivars Hikmok sorip et Majesta. Hikmok sorip a été choisi en raison de sa capacité hors norme à accumuler le Si (jusqu’à 3,6 % M.S. selon Deshmukh et al. 2013) alors que Majesta présente des qualités agronomiques intéressantes, notamment face à la maladie. De ce croisement ont été obtenus 141 RILs de génération F8, au sein desquels 768 marqueurs SNP (single nucleotide polymorphism) ont été identifiés. La cartographie de QTL (Quantitative Trait Loci) a permis de mettre en évidence une région située sur le chromosome 16 du soya sur laquelle ont été identifiés trois gènes candidats (GmLsi2a, GmLsi2b, GmLsi2c) pouvant agir en tant que transporteur d’efflux de Si. Il s’agit de la première identification d’un locus génétique directement lié à une accumulation différentielle de Si chez les plantes. Le classement de 328 lignées de soya en neuf haplotypes différents selon le génotype rencontré chez les gènes GmLsi2b et GmLsi2c a permis de lier l’haplotype H1 à une accumulation accrue de Si chez le soya. La fonction d’efflux du gène GmLsi2c a été confirmée dans les ovocytes de Xenopus laevis (données non publiées).
20
Tableau 2. Diversité des haplotypes détectés avec des SNPs sélectionnés dans les gènes GmLsi2b et GmLsi2c à partir de 328 lignées reséquencées publiques et privées (Syngenta) (données non
publiées)
2.3 Approche génétique
Tel que mentionné précédemment, ce sont les plantes qui accumulent du Si qui tirent le plus grand bénéfice d’un apport. Il semble que l’amplitude des effets observés soit directement liée à la quantité de Si absorbée par une plante, d’où l’intérêt envers des cultivars montrant une accumulation supérieure de Si. La découverte fortuite de la capacité hors du commun du cultivar Hikmok sorip à accumuler du Si a ouvert la voie à de nombreuses hypothèses de recherche portant sur le soya et l’absorption de Si. Considérant aussi que le cultivar Hikmok sorip a montré un niveau de résistance significativement accru contre la rouille du soya lors d’essais avec du Si (Arsenault-Labrecque et al., 2012), cela laisse supposer qu’une augmentation de la capacité d’absorption de Si pourrait se traduire par d’importants bénéfices. Il a été démontré par le passé qu’il est possible de modifier la capacité d’absorption du Si chez une plante non accumulatrice, en l’occurrence Arabidopsis (Montpetit et al., 2012). En effet, Arabidopsis ne possède pas de Lsi1, mais possède un gène
21
homologue Lsi2 (Ma et al., 2015) et selon Montpetit et al. (2012), la plante accumule moins de 0,1 % de Si dans ses parties aériennes. Les résultats prometteurs obtenus avec des lignées transgéniques TaLsi1 (transporteur d’influx du blé) à l’aide de ce système d’expression nous ont donc encouragés à tenter de démontrer le potentiel de ce ou ces gènes (GmLsi2b et/ou GmLsi2c). Ce projet visait ainsi à vérifier si les allèles spécifiques de GmLsi2 retrouvés chez de rares variétés de soya confèrent une absorption accrue de Si.Ce projet s’inscrit dans le processus de propriété intellectuelle et les résultats seront inclus dans le dépôt d’un brevet.
CHAPITRE 2
MANUSCRIT
25
CHARACTERISATION OF SILICON (SI) EFFLUX TRANSPORTERS IN SOYBEAN
RÉSUMÉ
Chez le soya, l’accumulation de silicium (Si) est un phénotype qui varie grandement entre les cultivars. Suite à la découverte d’un transporteur d’influx (Lsi1) en 2013 par Deshmukh et al. aucun lien entre l’accumulation différentielle de Si et le gène n’a été établi. Par la suite, l’approche par QTL a permis d’identifier une région d’intérêt dans laquelle on retrouve trois gènes (GmLsi2a, GmLsi2b, GmLsi2c) apparentés au Lsi2. On rencontre pour les gènes GmLsi2b et GmLsi2c neuf haplotypes (H1 à H9) différents (données non publiées). H1 est le seul haplotype qui présente une accumulation accrue de Si. Ce projet visait à vérifier si les allèles spécifiques de Lsi2 retrouvé chez de rares variétés de soya confèrent une absorption accrue de Si. Pour ce faire, nous avons utilisé un système hétérologue d'expression dans Arabidopsis thaliana, et nous avons comparé l’accumulation de Si des plantes transgéniques générées. Dans un premier temps, les résultats ont révélé que le promoteur utilisé aux fins d'expression de Lsi2 pouvait grandement influencer le taux d'absorption de Si. De façon générale, il a été difficile d'observer des différences d'absorption entre les allèles et même avec ou sans les allèles. Toutefois, sur la base de la sélection des cinq meilleurs transformants de chaque allèle, les essais réalisés ont montré une différence d’accumulation significative de Si entre les allèles du gène GmLsi2c, alors qu’aucune différence n’a été observée chez le gène GmLsi2b. En dépit des limites associées avec l’utilisation d’Arabidopsis comme plante modèle, nos résultats supportent que les gènes à l’étude agissent comme transporteur d’efflux de Si. Il s’agit du premier système végétal dans lequel on observe une variation intraspécifique aussi élevée liée à la présence d’un transporteur de Si. Les connaissances acquises pourraient servir au développement d’une approche génétique combinant l’accumulation de Si et la lutte contre les stress chez les plantes.
26
ABSTRACT
A great variability for silicon (Si) accumulation is encountered between soybean cultivars. In 2013, a Si influx transporter (Lsi1) has been identified in soybean by Deshmukh et al., initially known as a non-accumulator, and thus confirmed the species’ ability to absorb the element. Deshmukh et al. (2013) tried to link the differential Si accumulation to Lsi1, in vain. Research was then oriented towards a possible Lsi2. A QTL approach led to the identification of three candidate genes closely related to Lsi2s (GmLsi2a, GmLsi2b, and GmLsi2c). Further genotypic analysis of GmLsi2b and helped differentiate nine haplotypes (H1 to H9) (unpublished data). Only H1 shows high Si accumulation and is only present in a few rare accessions. This project aimed to verify whether the specific Lsi2 alleles found in rare soybean accessions is indeed responsible for high Si accumulation. We used a heterologous expression system in Arabidopsis thaliana, and we compared Si accumulation in transgenic plants. Initially, results revealed that the promoter used for Lsi2 expression could greatly influence Si absorption. In general it was difficult to observe differences in absorption among genes and even among alleles. However, based on the selection of the five best transformants for each allele, results showed significant differential accumulation between GmLsi2c alleles, whereas no difference was observed for GmLsi2b. Despite the limitations associated with the use of Arabidopsis as a model plant, our results support that GmLsi2b and GmLsi2c acts as Si efflux transporters. This is the first report of an intraspecific variation in Si accumulation linked to the presence of Si transporters. This knowledge could be used to develop a genetic approach combining the accumulation of Si and the alleviation of biotic and abiotic stresses in plants.
27
INTRODUCTION
Soybean (Glycine max (L.) Merr.) is one of the most important cultivated crops in the world, and is a mainstay of the economy in North America (FAOSTAT, 2015). Demand for the crop remains strong and is continuously expanding because of its various uses (Wilson, 2008). Soybean is mainly exploited for oil production and as a source of protein in human and livestock food supply. One way to support the increasing soybean demand is through yield improvement and reduced losses. Indeed, soybean crop losses to diseases, pests and abiotic stresses cost billions of dollars every year (Tyler, 2007). It has been estimated that losses caused by soybean pathogens amount to over $4 billion dollars annually, in the United States alone (Liu et al., 2015). In that regard, the source of genetic diversity that is kept among the accessions of the world’s soybean germplasm collections can contribute to future advances in genetic improvement. Association of phenotypic traits with genotypic markers is a very useful way to identify unique accessions, which in turn provide valuable genes in variety development (Wilson, 2008). The genetic resources in Glycine max can be used in innovative ways, by the integration of genomic tools with modern breeding programs.
Over the last decade, the major seed companies have invested a lot of effort in breeding programs. With the advent of Next-Generation Sequencing (NGS), the number of crop genomes sequenced has increased steadily every year since the publication of the Arabidopsis thaliana genome in 2000 (AGI, 2000). The soybean genome was published in 2010 by Schmutz et al. and was the first legume species with a complete genome sequenced. Despite numerous studies, only a few cases of genes underlying important agronomic traits have been reported. The identification of candidate genes was found to be successful in some plant species including Arabidopsis (Verslues et al., 2014), maize (Li et al., 2013) and rice (Zhao et al., 2011). Subsequent allele mining of identified gene provides new sources of useful genetic traits, which is a key element for breeding purposes.
Soybean being an important cultivated crop with a well annotated genome, a lot of effort has been invested in the characterization of the genetic diversity of wild and cultivated
28
soybean genotypes over the years. One of the great challenges for plant breeders is to understand the biological determinants of quantitative phenotypic variation. To date, several soybean phenotypes exhibiting different traits improving agricultural performances have been studied. Research has been predominately oriented on resistance to biotic and abiotic stresses, since these are responsible for most of crop losses. A promising trait that has recently gained a lot of attention in many crops is silicon (Si) accumulation. This interest has been sparked by the discovery of specific Si transporters in plants (Ma et al., 2006; 2007) and by the multitude of reports showing the benefits of Si accumulation in alleviating various stresses such as diseases, insects, radiation and drought (Liang et al., 2015).
Silicon is present in the soil mostly in the form of amorphous silica (SiO2), and is absorbed
by plant roots only in the form of silicic acid (H4(SiO)4) released by SiO2 (Epstein, 1999).
Some plants can accumulate as much as 10 % Si dry weight in the leaf tissues (Eptein, 1994), but the concentrations vary greatly among plant species (Hodson et al., 2005). Monocots are generally recognized as strong accumulators with concentrations exceeding 1 % dry weight as opposed to most dicots that generally absorb less that 0.5 %. It is now known that the uptake of silicic acid is mediated by two Si transporters: Lsi1, which is described as an influx transporter (Ma et al., 2006), and Lsi2, an efflux transporter (Ma et al., 2007). Originally described in rice, homologous Si transporters of Lsi1 and Lsi2 have been identified in other plants, including maize, barley (Mitani et al., 2009), pumpkin (Mitani-Ueno et al., 2011) and soybean (Deshmukh et al., 2013; 2015).
Until recently, soybean was considered to be a low Si accumulator. However, work by Arsenault-Labrecque et al. (2012) and Guerin et al. (2014) highlighted that some soybean cultivars could accumulate more than 2 % Si. Based on an extensive analysis of aquaporins in plants, Deshmukh et al. (2013) identified functional Lsi1s in soybean, thereby confirming the species' ability to absorb Si. In the same study, the authors showed that some soybean cultivars could accumulate up to 4 times more Si than others. This striking difference led to the hypothesis that the differential expression of the two Lsi1 genes (GmNIP2-1, GmNIP2-2) accounted for the higher accumulation in some cultivars. However, further studies
29 indicated there was no difference in expression or in sequences of the aquaporins among cultivars.
In an attempt to identify the factor(s) explaining the differential absorption between soybean cultivars, Deshmukh et al. (personal communication) analyzed a population of 141 RILs made from a high Si-accumulating and a low Si-accumulating parent. This led to the identification of a single QTL encompassing three putative Lsi2 genes, hereafter called GmLsi2a, GmLsi2b, and GmLsi2c. The GmLsi2b and GmLsi2c genes have some structural differences compared to the alleles found in a low Si-accumulating parent, but it is unknown if these differences can indeed account for the different phenotypes in terms of Si accumulation.
It was previously shown that it is possible to increase Si accumulation in Arabidopsis thaliana, a plant lacking a Lsi1 transporter, by the insertion of an influx transporter from wheat (Montpetit et al., 2012; Vivancos et al., 2015). In the same manner, we hypothesized that we could observe differences in Si accumulation in Arabidopsis if they expressed highly functional Lsi2 from soybean. The objective of this work is to verify if the specific GmLsi2 allele found in rare soybean accessions is responsible for differential ability in Si uptake in high absorbing soybean cultivars. To this end, soybean Si transporter genes were introduced into Si-accumulating Arabidopsis transformants under the control of root-specific promoters, and the transgenic lines were evaluated for their ability to accumulate Si in their tissues.
MATERIALS AND METHODS
Plant material and growth conditionsFour different Arabidopsis genotypes [Colombia (Col-0; Ohio State University), TaLsi1 (Montpetit et al., 2012), TaLsi1 GmLsi2b and TaLsi1 GmLsi2c transgenic lines] were used in the present work. For all experiments, seeds were surface-sterilized (5 % bleach, 2 min), rinsed five times with sterile water and stored at 4C for 3 days to break dormancy. Col-0 seeds were directly sown on Veranda Container Mix (Fafard et frères) in a growth
30
chamber under long-day conditions (14 h of light at 22C, 10 h of dark at 19C, 55-65 % humidity and a light intensity of 150 mol/m2/s) and covered with plastic sheets for one
week. Seeds of TaLsi1 lines and T2 TaLsi1 GmLsi2 lines were selected on Murashige and Skoog Basal Medium with Gamborg’s vitamins (MS) (Sigma-Aldrich) containing hygromycin (15 mg/L) for TaLsi1 lines and kanamycin (50 g/ml) for TaLsi1 GmLsi2 lines. At day 10, seedlings of uniform size were transferred to pots containing Veranda Container Mix at a density of five plants per pot. Plants were treated with water containing 1.7 mM potassium silicate (K2SiO3). Only controls (Col-0 and TaLsi1 lines) received a treatment without
soluble Si, in which potassium chloride was used to provide an equivalent amount of potassium. Plants were maintained in a growth chamber as described above. Arabidopsis plants of different genotypes were used for experiments one month after transplanting.
Isolation of promoter region, construction of promoters::GUS reporters and plant transformation
The 2.5 kb region upstream of the initiation codon of the NIP5;1 gene (AT4G10380) was amplified from a BAC clone. The 290bp region upstream of the initiation codon of the CASP2 gene (AT3G11550) was amplified by PCR from genomic DNA extracted from Col-0 Arabidopsis plants using a high fidelity polymerase (Phusion, New England BioLabs). Primers were designed to amplify promoters and to introduce SmaI and HindIII or SbfI restriction sites (see Table 1). PCR products were cloned in pGEM-T easy using a ligation kit (Takara). Promoters were then cloned in TOP 10 E. coli cells and clones were screened for presence of insert with colony PCR. Next, plasmids were recovered from a fresh bacterial culture using the QIAprep Spin Miniprep kit (Qiagen). Finally, 1 g of pure plasmid DNA was digested with restriction enzymes to assess the presence of an insert of the expected size, followed by confirmation of the amplicons by DNA sequencing.
Promoters were inserted into the plasmid pBI121 (Clontech), a binary vector harbouring a GUS reporter gene. Promoters were inserted into the SmaI and HindIII or SbfI sites in order to replace the CaMV 35S promoter and ligation was assessed using a ligation kit (Takara).
31 Cloning in TOP10 E. coli cells for multiplication was made prior to cloning in Agrobacterium tumefaciens strain GV3101 for plant transformation.
Col-0 Arabidopsis plants were transformed using a modified floral dip method (Zhang et al., 2006). Independent transgenic lines (T1) were selected for Kanamycin resistance (50 g/ml) on MS medium (Sigma-Aldrich) and the presence of the regulatory regions was verified by PCR (see Table 1). T2 transgenic seeds were harvested and sown on MS medium containing Kanamycin (50 g/ml) for 10 days and transferred into Magenta boxes for growth. T2 transgenic plants were used for phenotypic analyses.
Histochemical GUS staining
Analyses were performed on three-week-old transgenic Arabidopsis plants. For histochemical localisation of -glucuronidase (GUS) activity, a -glucuronidase reporter gene staining kit (Sigma) was used according to the manufacturer’s instructions. Incubation was performed in the dark at 37C overnight and tissues were washed twice with 100 % ethanol until the chlorophyll pigments were completely bleached. Whole plants were observed directly under binocular (Olympus) and light microscopes (Olympus).
Construction of plant expression vectors and plant transformation
The two GmLsi2 soybean candidate genes (GmLsi2b and GmLsi2c) were identified and their cDNAs sequenced prior to this work (unpublished data). Following haplotype analysis, each gene has several distinct alleles, linked to one of the nine haplotypes identified. Here, we contrast two haplotypes, H1 and H5. We observe a unique allele combination for GmLsi2b and GmLsi2c for each haplotype, but genes were tested one at a time in our system. H1 is the haplotype present in high Si-accumulating cultivars, and H5 is the most common haplotype showing a phenotype of low Si accumulation. All genes (both alleles of GmLsi2b and GmLsi2c were present in either the H1 or H5 haplotype) were synthesized (Genscript) in pUC57 with SmaI and SacI sites to ensure sequence accuracy. A transgenic TaLsi1 line was used to express GmLsi2b and GmLsi2c. Conventional molecular cloning techniques were applied to generate the plant expression vectors. Binary vector pBI121 containing either a
32
NIP5;1 or CASP2 promoter was digested with SmaI and SacI in order to remove the GUS reporter gene. All synthesized genes were also digested with SmaI and SacI. Ligation of four different GmLsi2 alleles in a vector containing one of two promoters (for a total of 8 different constructs) was performed. Constructs were cloned in TOP 10 E. coli cells and clones were screened for the presence of the insert with colony PCR. Next, plasmids were recovered from a fresh bacterial culture using the QIAprep Spin Miniprep kit (Qiagen). Pure plasmid DNA was digested with restriction enzymes and sent for sequencing for confirmation. One positive clone for each construct was cloned in Agrobacterium strain GV3101 using a modified freeze-thaw method (Jyothishwaran et al., 2007) and, after validation with colony PCR, one clone per construct was selected for plant transformation. Arabidopsis was transformed according to a modified floral dip method (Zhang et al., 2006). Independent T1 transgenic lines were selected on MS medium (Sigma-Aldrich) containing Kanamycin (50 g/ml), and the presence of the GmLsi2 transgene was verified by polymerase chain reaction (PCR) (see Table 1). T2 seeds were harvested from independent transgenic lines bearing each construct, respectively, and sown on MS medium containing Kanamycin (50 g/ml). For all experiments, the phenotype of the T2 transgenic plants was analyzed.
Determination of Si concentration in transgenic Arabidopsis shoots
Transgenic lines TaLsi1, TaLsi1 GmLsi2 and Col-0 plants, treated or not with Si, were analysed in this study. The Si content in experimental plants was measured by colorimetric analysis following an HCL-HF extraction (Taber et al., 2002). Aerial parts of the plants from each treatment (5 plants per line) were collected and freeze-dried one month after the beginning of Si amendment. Samples were ground to a powder before Si analysis. For each treatment a minimum of five biological replicates were used.
33
Statistical analyses
Statistical significance was assessed with Student’s t-test and Dunnett’s test using JMP 12 software (SAS institute Inc.). Least square means were used to express the results. Standard error was used as the error bar in figures.
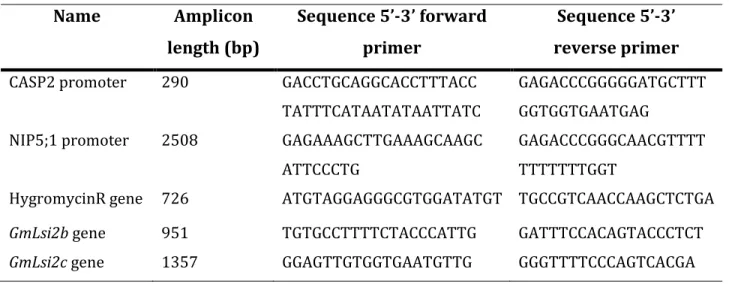
Table 1. List of primers used for screening of transformants
Name Amplicon length (bp) Sequence 5’-3’ forward primer Sequence 5’-3’ reverse primer
CASP2 promoter 290 GACCTGCAGGCACCTTTACC
TATTTCATAATATAATTATC
GAGACCCGGGGGATGCTTT GGTGGTGAATGAG
NIP5;1 promoter 2508 GAGAAAGCTTGAAAGCAAGC
ATTCCCTG
GAGACCCGGGCAACGTTTT TTTTTTTGGT
HygromycinR gene 726 ATGTAGGAGGGCGTGGATATGT TGCCGTCAACCAAGCTCTGA
GmLsi2b gene 951 TGTGCCTTTTCTACCCATTG GATTTCCACAGTACCCTCT GmLsi2c gene 1357 GGAGTTGTGGTGAATGTTG GGGTTTTCCCAGTCACGA
RESULTS
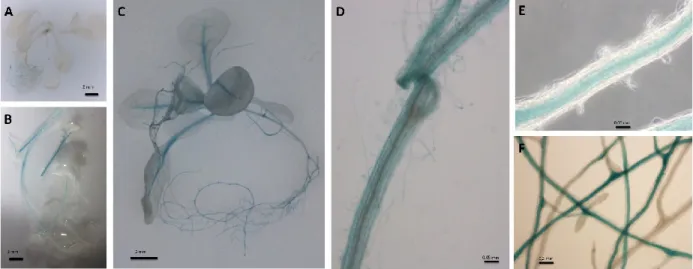
Analysis of promoter::GUS reporter constructs in Arabidopsis
To assess the tissue specificity of both promoters, we generated transgenic Arabidopsis plants expressing the -glucuronidase (GUS) gene. Transgenic Arabidopsis plants were generated under the control of two different promoters; the NIP5;1 promoter and the CASP2 promoter. Six independent transgenic lines were generated for the CASP2 promoter and 15 for the NIP5;1 promoter. GUS staining patterns were observed in young transgenic plants of the T2 generation in both shoots and roots. One of the pCASP2::GUS reporter constructs and three of the pNIP5;1::GUS reporter constructs showed no activity for the GUS gene (data not shown). Also, we observed a heterogeneous expression pattern of the reporter gene within both constructs. Indeed, although promoter expression was mostly found in roots, expression in aboveground tissue was also observed in some of the lines. For pCASP2::GUS, two lines showed activity exclusively in the roots, while three showed activity
34
in roots and aerial tissues (Figure 1). Microscopic observations revealed expression in the endodermis of mature roots supporting the report of Christ et al (2012) (Figure 1). For pNIP5;1::GUS, ten lines showed activity exclusively in the roots, while two showed activity in both roots and aerial parts (Figure 2). Microscopic observations revealed expression in all root cells, with high expression in the elongation zone as reported previously by Takano et al. (2006) (Figure 2).
Figure 1. GUS expression driven by the CASP2 promoter in transgenic T2 Arabidopsis
plants . Weak GUS expression in the roots (A), strong GUS expression in aerial parts (B), strong GUS expression in both roots and aerial parts (C), GUS expression in root cortex (D), and in root endodermis (E) and (F).