Sleep habits, academic performance, and the adolescent brain structure
Texte intégral
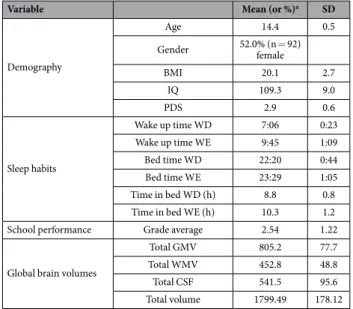
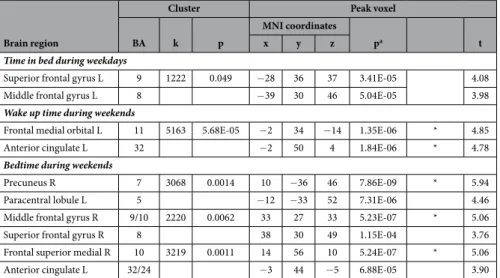
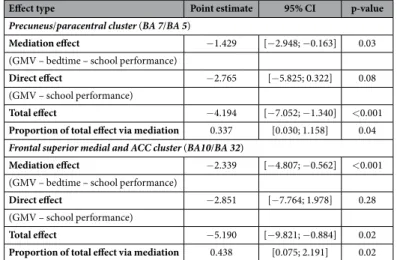
Figure
Documents relatifs
Even if the two-set snowmelt module presents better representation of MODIS SCA during winter and monsoon than the one-set version, the model still struggles to correctly represent
I asked Dick Peter whether he would take me in his lab to study the origin of the pituitary innervation using the brain lesioning technique developed in the goldfish and he said…
Nevertheless, the results of our model demonstrate that even in the presence of congestion effects in main cities, trade openness increases concentration toward them, inducing
Dans une précédente recherche (Amblard, 2010), j’ai montré comment s’opèrent les processus réflexifs, dans et hors de l’action pédagogique, chez des sujets usagers
Accurate image reconstruction for conventional (FFOV) helical scanning with axial truncation can be demonstrated using the chord method and the two-sided finite inverse
To further characterize the intron promoter and to deter- mine what portion or portions were necessary for promoter activity, we transfected parasites with constructs that
The choice of the excitations (I) is based on the expenence gained in the calculation of KX- spectra of Ne, Ar [5] and on using the smallness parameter characterinng the mixing of
We differentiate internal factors (cost of capital, operating expenses and loss provisions) from the external ones (regulation, competition and inflation).The outcomes of
