Contribution à l'étude de quelques marqueurs physiologiques de tolérance au déficit hydrique chez le blé dur : taille de semences en tant que critère de sélection
141
0
0
Texte intégral
Figure


+7

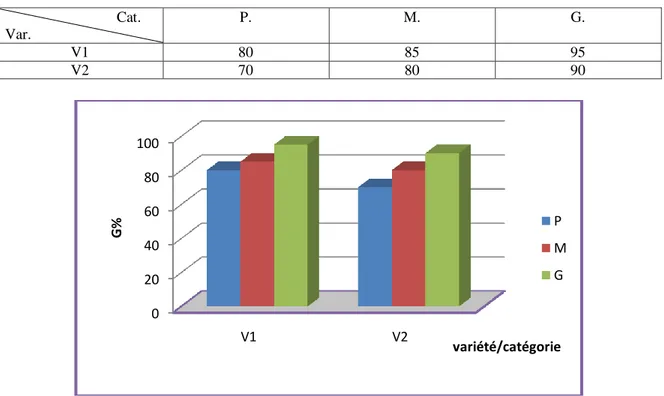
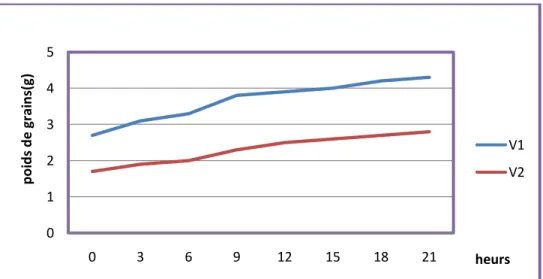
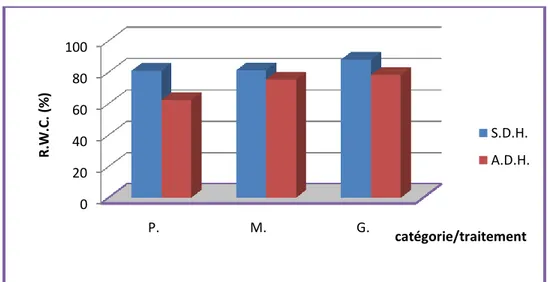
Documents relatifs
