I
DEPOSITED BY THE FACULTY OF GRADUATE STUDIES AND RESEARCH
I_x
M
. \ Gc5-- \
':J4
\
M~GILL
UNIVERSITY
LIBRARY
UNA CC.
1941
'·
- -
--
J
LA DIAPAUSE CHEZ LES TENTHREDES
par
A.R.Gobeil
Soumise
a
la Faculte des Gradues de l'Universite McGillen accomplissement partiel des exigences requises pour le degre de
Docteur en Philosophie
TABLE DES MA.TIERES
Pages;
Introduction. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • . • 4 I. La Diapause et ses Causes ••••••••••••••••••••••••••••••••• 10 1. La glande d'hibernation et les glandes hormogenes ••• 11
2. La diapause: un phenomena hereditaire? •••••••••••••• 13 3. Influence des facteurs internes ••••••••••••••••••••• 16 4. Facteurs physiques ou externes •••••••••••••••••••••• 18
a. Temperature ••••••••••••••••••••••••••••••••••• 18 b. HUlllidi te. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 19 c. Lumiere ••••••••••• • ••••••••••••••••••••••••••• 21
5. Facteurs chimiques •••••••••••••••••••••••••••••••••• 22 6. La diapause et l'intoxication de l'organisme •••••••• 23
7. La theorie de Bodine •••••••••••••••••••••••••••••••• 25 8. La theorie de Cousin •••••••••••••••••••••••••••••••• 26
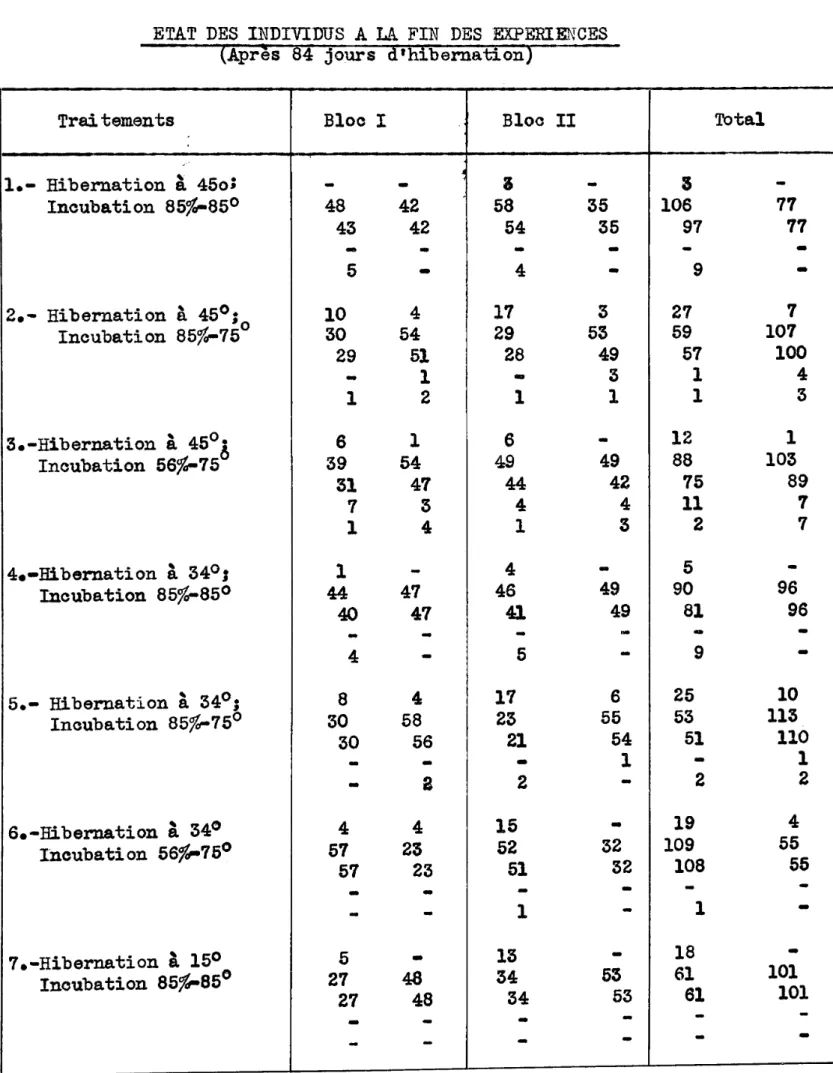
II. Materiel et Methodes••••••••••••••••••••••·•••••••••••·••• 31 1. Provenance des individus ••••••••••• • • • • • • • • • • • • • • • • • 31 2. Especes etudiees •••••••••••••••••••••••••••••••••••• 34 a. Diprion polytomum ••••••••••••••••••••••••••••• 34 b. Nematus erichsonii •••••••••••••••••••••••••••• 37 c. Autres espeoes •••••••••••••••••••••••••••••••• 40 3. Technique. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 40 a. Saison 1938-39 •••••••••••••••••••••••••••••••• 40 b. Saison 1940-41 •••••••••••••••••••••••••••••••• 42
III. Influence de la Temperature et de l'Etat HYgrometrique
de 1 'A-tmosphere •••••••••••••••••••••••••••••••••••••••••• 47
1. Action des temperatures d'hibernation ••••••••••••••• 47 a. Mortalite durant l'hi.berna.tion •••••••••••••••• 47 b. Limites vitales inferieures ••••••••••••••••••• 50 c. l'1:ortal i te durant la nympho se. • • • • • • • • • • • • • • • • • 50 d. Action des temperatures d'hibernation sur
l'e-mergence •••••••••••••••••••••••••••••••••••••• 54 2. Determination de la temperature et de l'humidite
optinla de nJ'Dlpho se. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 57 a. Developpement
a
60°F. dans un milieu tres sec. 57 b. Mortalite et developpementa
75°F. et 8QOF.dans des conditions hygrometriques variables •• 58 c. Limites vitales superieures ••••••••••••••••••• 61
3. Influence de variations brusques de temperature ••••• 63 a. Mortalite durant la nymphose •••••••••••••••••• 64 b. Emergence. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 67 4. Nymphose sans exposition aux basses temperatures •••• 69
IV.
Influence d'Irnmersions sur la Rupture de la Diapause ••••• 70 1. Technique et considerations generales ••••••••••••••• 70 a. Vitalite de Diprion polytomum dans l'eau •••••• 73 b. Constitution de l'humus et acidite des solsforestiers •••••••••••••••••••••••••••••••••••• 74 c. Preparation de solutions
a
pH5 et pH3 ••••••••• 772. Mortalite durant !'hibernation •••••••••••••••••••••• 78 a. Diprion polytomum ••••••••••••••••••••••••••••• 78
b. Nematus erichsonii •••••••••••••••••••••••••••• 83
3. ~ortalite durant la nymphose •••••••••••••••••••••••• 85
a. Immersions dans l'eau ••••••••••••••••••••••••• 91
b. ]mmersions dans des solutions acides •••••••••• 92
4. Ein.ergence. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 94
v.
Action d'Lmmersions dans l'Acide Sulfurique Concentre •••• l03 1. Resistance de Diprion polytomum dans H2so
4concentre. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 104
2. l,;~ortali te durant 1 'hibernation et la nymphose... • • • 107 3. Emergence.. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 111 VI. Influence du Milieu sur la Rapidite de la Nymphose •••••• 113 VII.
La
Diapause et 1'Intoxication des Oeufs ••••••••••••••••• 122 VIII.Discussion •••••••••••••••••••••••••••••••••••••••••••••• 126 Conclusions... • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 137 Litterature Citee •••••••••••••••••••••••••••••••••••••••••••• 141 P1anches ••••••••••••••••••••••••••••••••••••••••••••••••••••• 152 Appendice. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 156INTRODUCTION
Du point de vue purement scientifiqu~
la diapause des insectes est un sujet fort interessant et qui a donne naissance
a
une myriade de publications. Par ailleurs, le cote economique de ce phenomena est tres important, et aotuelle-ment au Canada, plus partiouliereaotuelle-ment dans 11est, oe sujet est degrande ao~talite pour les entomologistes forestiers.
En
effet, notre gros probl~me presentement, celui qui reclam.e le plusl'attention des entomologistes, comma cella des forestiers d'ailleurs, est cree par les ravages de la mouohe
a
soie euro-peenne de l'epinette, Diprion polytomum. Ce defoliateur, que l'on trouve dans la majorite des peuplements d1epinette de laprovince de Quebec exhibe dans oertaines regions deux et meme trois generations par annee, alors qu'ailleurs, plus au nord, la pronymphe peut demeurer end iapause dans son cocon jusqu
•!
6 ou 7 ans avant de donner naissancea
l'adulte.Cette longue diapause presente des desavantages mais aussi des avantages pour la survivance de l'esp~oe. Il est vrai que des individus subissant un arret de plusieurs annees dans leur developpament auront un potential biotique beaucoup plus bas que s'ils avaient 2 generations ou
plus par annee. Il est vrai aussi que dans la partie nord de
son habitat, les prinoipaux ennemis de ~.polytomum, les musareignes et les ec~reuils, se~ont des agents de controle plus effioaces, la pronymphe etant
a
leur meroi durant une plus longue periodeo Dtunautre oate# oertaines methodes de lutte chimique ou sylvicole~ de
....
meme que oertains faoteurs physiques ou biologiques auront moins de suooes paroe que n'affectant qu'une partie de la population. Ainsi durant l'ete 1940 une maladie ohez les larvas~ apparamment
oausee par un virus, a reduit la population larvaire de faQOn substantielle dans la Gaspesie, Gobeil (1940). Mais son effet sur la population future de l'insecte sera loin d'etre aussi sensible qu'il le fut, par example, dans le Ver.mont et le New Hampshire, alors qu'a oertains endroits la mouohe
a
soie futa
peu pres eliminee, apparemment par la m3me maladie• Dowde.n {1940). Pour toutes oes raisons une etude des faoteurs pouvant affecter la diapeuse de la mouohea
soie europeenne de l'epinette est done tres importa.nte.Comme resultat de l'epidemie de Diprion polytomum, le Service d1Entomologie du Ministere des Terres
&
Forets de la province de Quebec~
A
l'instar du Service Federal d'Entomologie~ organisait en 1938 un systeme de reconnaissance annuelle de la fauna entomologique de la provinceoEn
rapport avec cette reconnaissance, nous mettons en elevage chaque annee plusieurs rnilliers des pecimens dont la majorite appartiennentl la familla des Tenthredinidae lesquels presentent de longues diapauses ou hibernations. Si nous voulons eviter une
accumu-lation d'individus d1une annee
a
l'autre et faciliter d'autantnotre travail, nous avons done inter~t
a
reduire au minimum la duree de cette periode de repos; et c'est la le deuxieme motif de cette etude.Le but immediat de ce travail etai t done d'etudier l'aotion de certains facteurs sur la rupture de l'hi-bernation de tenthredes ayant veou dans leur milieu habitual
jusqu'au stage de codon, et determiner lequel ou lesquels de ces facteurs agissent avec le plus de force.
Pour cela n'ai d1abord etabli 1 'influence
de differentes temperatures d'hibernation et de nymphose afin d1en arriver
a
la temperature optimum pour chacun des deux etats.Aces temperatures optima et pour differentes periodes d'hiber-nation, j•ai fait agir soit durant l'hiberd'hiber-nation, soit durant la
nymphose~ soit encore durant l'hiber.nation et la nymphose, d'autres
facteurs ou agents susceptibles de provoquer la rupture. Puis,
a
l'aide d'analyses statistiques j'ai pu evaluer et comparerl 1influence de chacun de ces facteurs ou agents dont les
princi-paux furent les sui vants:_ : variations brusques de temperature ou chocs the~ques; atmosph~re desseohee et tres humide;
immersions dans 1 'eau;: immersions dans des solutions acides
a
des pH comparablesa
ceux qui existent dans le sol; immersions dans de l'acide sulfurique ooncentre.J'ai ensuite cherche
a
relier, si possible6 l'influenoe de oes facteurs aveo les conditions de ~lieu pouvantexister
a
l'etat naturel. Ehfin, conme les causes de la rup~resont apparemment en relation etroite aveo oelles de l'entree en diapause, j•ai fait entrevoir les agents possibles de diapauses durables ohez les tenthredes. Dans oe dernier cas, oe ne sont la, oependant, que des hypoth~ses, l'origine des individus utilises ne me per.mettant pas d'etre plus oategorique.
En
effet, oe~oifurent tous recueillis
a
l'etat de oooon, o'est-a-dire qu'a part quelques donnees bien generales sur le olimat de la n&gion, nous n'avions auoun detail sur les faoteurs ayant pu affeoter le deve-loppement anterieur de oes individus. Aveo Cousin; je orois qu 'il est diffioile de fai re des deductions vrei es, quant aux causes de la diapause, en observant des animaux recueillis dans leur milieu naturel. D'apr~s cet auteur pour pouvoir mettre oes causes en evidence, il est obligatoire d'avoir des elevagesmethodiquement suivis depuis l'oeuf jusqi'a l 1adulte, et de se
servir d'animaux ayant veou dans des conditions aussi semblables que possible. Le milieu natural ou exterieur en est un essential-lement fluctuant et la plupart du temps defavorable a l'insecte. C 'est pou·r~u~i.
1
bien que j 'aie oru pouvoir tirer certainescon-clueions sur les agents responsables de l'entree en diapause•
oelles-oi doivent done etre considerees aveo une oertaine reserve. Apropos de l'usage des statistiques,
quelques technicians sont encore reticents quant a son importance et
a
son utilite en biologiee C'est toutefois une grave erreur;et sans l'aide de !•analyse statistique il est souvent impossible d'arriver
a
des conclusions definies. Ceci est particuliereme.nt vrai lorsqu'il s'agit de recherohes experimentales comme dans le present aa.s. Si nous placons des individus dans deux milieuxdifferents et qu'un de ces milieux leur est fatal, il n'est peut• etre pas necessaire alors d'avoir recours aux statistiques pour tirer conclusion. Mais si, par oontre, !'animal complete son
oyole vital dans les deux milieux aveo seulement quelques nours de difference dans le developpement total, co~~ent pouvons-nous pretendre qu•un milieu est nettemant superieur
a
l'autre et que cette di~ference n'est pas simplement due au hazard ? Dans un tel cas. et on peut les multipliera
1 'inifi et les <n mpliquera
plaisir, la meilleure interpretation nous l'obtiendrons
a
l'aide des statistiques. Comma question de fait, sans les statistiquesje crois qu'il est impossible d'etudier
a
fond un probl~me de biologie et surtout de le faire avec un minimum d'experiences tout en obtenant le maximum de rendement.Les experiences preliminaires
a
ce travail furent executes au College MacDonald au cours de 1 'hiver 1938-39 sous la direction du Prof.Melville Du Porte, dont les conseils me furent alors tres utiles. Il me fait plaisir de lui enQ
·~Je suis aussi tr~s oblige au Prof.Samuel Graham de l'Universite pour avoir bien voulu me diriger dans la preparation de mon pro gramme pour 1 'hi ver 194()-41 et m' a voir fait de precieuses suggestions.
Enfin je tiens
a
remercier le Prof.R.s~~erby du College Macdonald et le Dr.J.W.Hopkins du Conseil National des Recherches pour l'aide qu'ils m'ont do.nne lors de
l'arrangement de mes experiences et de l'analyse statistique de mes resultats.
CHAPITRE I
LA DIAPAUSE ET SES CAUSES
Le ter.me diapause fut employe pour la premiere fois par Wheeler (1893) pour designer une periode de repos durant le developpement embr~ronnaire. Plus tard~ Henneguy (1904) a etendu le sens de ce mot
a
tout arret dans le developpe.mant ontogenetique d'un animal~ depuis la fecondation de l'oeufNous aurions done ainsi des diapauses
emb~onnaires, larvaires et nymphales.
Les mots hibernation et estivation sont aussi employes pour signifier les diapauses survenant durant l'hiver et l'ete. Shelford (1929) et Bodine (1932), cependant, font une distinction entre la diapause et l 1hiben1ation. Il faudrait
appeler hibernation les cas ou l'arr~t dans le developpement est cause par la tew~erature seulement (Bodine) ou par des conditions defavorables (Shelford). La diapause, d'un autre cote, serait inherente
a
l'espeoe (Bodine) et le developpement arreterait spontanement ( Shelford)o Ce serait la une distinc-tion tres importante s'il etait prouve que la diapause et un phenomena hereditaire ou spontane, mais plusieurs auteurs ne partagent pas cette maniere de voir.1
fi ·~ ..' ...
1a diapause est Pn sujet qui a ete tres disc·u:te et sur lequel des opinions variees ont ete emises. C'est pourquoi, je ne donnerai ici qutun apercu des plus importantes
et des p1us recentes theories. Pour pl,J.s ample information, on consultera Dubois (1896), Athanasiu (l909),l3aumberger(l914) Cousin (1932) Richards et I,Iiller (1937).
1. LA GLANDE D'HIBERNATIOH ET LES GLANDES HORMOliOGENES
earlier (1893) maintient que la soi-disant glande d 'hibernation es·t tr~s developpee chez taus les animaux hi ber-nants et e.gi t d 'abord comme entrepSt pour les corps gras. Plus
tard, la glande sert directement pour nourrir l'animal durnnt sa longue periode de jettne, en se trensfor.mant, quelques cellules
a
la fois, en une matiere de grande valeur nutritive. Cette transformation ser~it probableme.nt regularisee par les nerfs des pieces buccales sous le controle du systeme nerveuxcentral. Seu.ls les animatLX possedant la glal".de d 'hibernation entreraient en diapause.
Depuis les
travaux de earlier, cepen-dant, cette glande aete
trouvee chez plusieurs animaux non -hibernants tels le rat, le chat et le singe, Sheldon (1924), et elle ne serait qu'une for.me de tissu adipatL~ en1bryonnaire. Rasmussen (1924) l'etudia chez le marmot et ne oroit pas quece soit une glande au sens strict du mot. D'autres organes~
ne oontenant pas essentiellement des tissus de reserve, foul~
d '~1i b erna ti on. Toutefois, une comparaison entre cette glande et le tissu adipetL~ ordinaire chez le marmot~ indique que les principales cellules des deux tissus sont de types distincts et Rasmussen suggera pour la glande d'hibernation le nom de tissu
e.dipeux multicellulaire.
Kopec (1922) croit que lorsque l'organisme est physiologiquement prepare pour repondre
a
son influence, unecertaine substance, secretee par l'hypophyse, serait suffisante pour occasionner la pupaison. Au cours d'une autre serie d'experiences (1924), il nourrit des larves de Lymantria dispar avec des extraits de Thyroide mais an rapport avec la diapause~
il obtint des resultats negatifs. Il conclut en disant que sa
premi~re hypoth~se sur l'in~luence des glandes endocrines du cerveau sur le metabolisme des insectes n'etait nullement inva-lidee par ces resultats negatifs. Par ailleurs, des experien-ces de nutrition executees anteri eurer:1en t pe.r Kunkel (1918), sur des larvas de la mouche Lucilia indiquent que l'alimenta-tion
a
la thyroide contribueraita
reduire la periods de nymphose.-~'.1igglesworth (1934-36) a beaucoup etudie
l'influence des hormones sur le developpement des insectes.
1_2
Apres avoir opere des nymphes de Rhodnius prolixus et leur avoir enleve 1 'hormone provoquant les mues~ le tt corpus alla turn." si tue
dans la t$te~ il a montre que les sujets ainsi traites ne crois-saient pas du tout, mGme apr~s avoir absorbe un repas completo De tels individus sont en diapause; et la croissance ou la metamorphose
n'au~e. lieu que si l 'hormone requiGe est introdui te dans le sang. Ceci di t-il suggere naturellement que la diap~atse chez les autres insectes, quand elle n'est pas occ~sionnee par les effets directs
du ~lieu, peut resul~,:,er d 'tm affaiblissement temporaire de
l'hor.mone promouvant la croissance. Dans certains cas cet affai-blissement serai t peut-etre du
a
Ul1 rhythme i::.u1e et dans d'au-tres cas aux effets indirects des facteurs du milieu.
Cepen-da~t en depit des resultats tres interessants obtenus par oet auteur, il est loin dtctre prouve que la diapause soit due simplement
a
un deperissement temporaire de l'hormone favori-sant la croissance tel qu'avance pa.r Wigglesworth.2. LA DIAPAUSE: UN PHSNOMENE HEREDI TAIRE ?
Parmi ceux favorisant cette theorie, Pictet
(1904-1922)~ semble en avoir ete un des plus ardents defen-seurs. Des experiences qu'il fit avec Lasiocampa 3-~E~,
Porthe~ia chrysorrhoea, Artia caja et autres, il conclut que
----la diapause est heriditaire et independante des saisons. Pour-tant ce meme auteur lors de son travail sur Lasioca.m.pa quercus montra qu'apres 6 generations, elevees
a
22°C, le rhythme de la diapa,, se c:i sparai t apr~s une diminution progressive de gene-ration en genere.tion. Un tel rhythme ne serai t done pas inva-riable et pourrait ~tre modifie lorsque l'insecte vit dans un milieu optimum et constant.Tower {1906) pretend que pour certaines espeoes, par suite de !'adaptation, l'hibernation est devenue permanente
et inherente et les " changements de milieu ne peuvent pas differer cette habitude, bien qu'ils peuvent la modifier
a
un degre conside-rable dans ses caracteres insignifiants." Plus tard (1917), pour illustrer ceci, il se servit de la betea
patates Leptinotarsadecemlineat~, qui apres 3 ans de transplantation dans la region desertique de Tuscon, Ariz., ne peut se readapter
a
son climat primitif de Chicago. Il expliqua ceci par l'evolution. Mais pourquoi cette reaction ne pouvait-elle pas se repeter aussi facilement lors du retour dans le milieu de Chicago ? Tower essaya d'expliquer cela par certaine action physico-c~imique.Il est
a
re.marquer que les individus furent introduits dans le desert de l'Arizona au printemps mais ramenesa
Chicago 3 ans plus tarda
l'automne. Si le transfert avait eu lieu au printemps et non apres la seoonde generation, juste avant !'hibernation, une partie de la population aurait peut-etrepu, comma ce fut le cas
a
~1SOOn, se reacclimater aux conditions de Chicago.Baumbe~ger(l914) basant son hypoth~se sur les travaux de Child attribua d'abord la diapause
a
de la surali-mentation ou encorea
un regime defeotueux oausant ainsi des accumulations de substances inactives dans le oytoplasmee Dans un travail posterieur sur Drosophila melanogaster(lll7). il ne peut, cependant, provoquer la diapause, bien que la sura-limentation puisse exister ohez cette esp~ce. Il conolua alors, en disant que la diapause est un phenom~ne p9riodique inherenta
l'espece.3abcock est un autre partisan de la theorie voulant que la diapause soit rhythmique ou hereditaire. En
1927, Babcock fit une serie d'observations sur l'hibernation de la pyrale du mais. Dans 11Est du Massachusetts cet insecte
a deux generations. alors que dans l'Etat de New York~ ou le climat n'est pas tres different, on ne trouve qu'une seule gene-ration. Afin de determiner les facteurs responsables de cette anomalie, il placa des larves provenant des deux milieux en incu-bateur sous des conditions de temperature et d'hurnidite diffe-rentes
a
savoir: milieu normal d'hibernation, milieu froid et sec, milieu chaud et sec. Il suivit le developpement de cesindivi-dus durant 4 ans pour en arriver
a
la conclusion qu'une periode de repos est essentielle au bien-~tre general de l'espece. Bien que cctte periode de repos soit inherente, il concede cependant, qu1elle peut ~tre modifiee par les facteurs de milieu. Au coursde son travail Babcock remarque, tres
a
propos, que si beaucoup d'experiences ont ete faites sur la rupture de la diapause, il semble y avoir eu tres peu d'essais pour evaluer les facteursh , '
produisant ces p enomenes. Nous verrons plus loin que Cousin (1933) fit des etudes tres poussees et probablement les plus comprehensives non seulement d~s facteurs brisant ou eliminant la diapause, mais aussi de ceux qui en sont la cause.
Parmi les autres auteurs considerant la diapause comme heriditaire ou rhythmique, mentionnons: Bodine ( 1923), Holmquist (1928), Readio (1931) et Hamilton (1936). Tous
admettent, oependant que oette qualite irulerente peut etre modi-fiee
a
un plus ou mains grand degre par les fa.cteurs du milieu.3. INFL:TEFCE DES FACTEURS Il:TERNES
Parker et Thompson (1927) ont assooie la dia-pause de la pyrale du mais avec le developpement des organes reproducteurs. Au printemps., certains changements clefinis se produj_sent ohez les rudiments d 'organes geni taux. Ils ont
~onstate 4 phases de developpement des gonades males, et la deuxieme p!lase serai t le debut d 'une acti vi te physiologique
qui, suivcnt le cas, elimine ou met fin
a
la diapause.LA
ou il y a deux generations par annee, les gonades de certains in-dividus de la premi~re generation seraient deja dans la deuxi~ me phase, bien que les ohenilles n'en sont encore qu•au 4iemestade larvaire; ces individus ne presenteront done pas de diapause. Par contre, les larves du 5ieme stade larvaire dont les organes genitaux sont encore dans la premi~re phase de developpement devront necessairement hibernere Ce serait oe qui se presente dans les regions ou il n'y a qu'une seule generation par annee. De oes observations~ Parker
&
Thompson ont oonolu que les facteurs proauisant oet arr3t dans le developpement de l'inseote, agiraient alors que les
larves sont encore relativement jeunes, et seraient
a
un1
~~·' /
JIL_ ""
haut degre sinon co~pl~tement, independants des conditions pre-valant au milieu de l'ete. Comme l'a fait remarquer Cousin, cet etat particulier des organes genitaux peut caracteriser la diapause de Pyrausta nubilalis, mais n'en est pas le facteur determinant et les causes doivent etre cherchees ailleurs.
Les memes remarques s'appliquant au processus enzymatique etudie par Tovmsend(l926), durant 1 'hiberne.tion de la pyrale du pommier Carpocapsa pomonellaJ et le peu de catalase note par Spooner (1927) chez le meme insecte. Tous ces ohangements~
tout cormne la reduction dans les eohanges respiratoires, Pembrey (1901-1903), la perte d'eau, Rulot (1902), et de gaz carbonique, Vernon (1897), sont des modifications prenant place durant l'hi-bernation et caraoterisant la diapause~ mais, apparemment, n'en sent pas les causes determinantes.
Plusieurs auteurs ont d'ailleurs d~~ontre qu•un metabolisme reduit au minimum pouvait exister, non seulement ohez les insectes en diapause, mais chez plusieurs animaux
non-hiber.nants, te1s la souris1 le lapin, le pigeon, et pouvait
etre obtenu au moyen de differents faoteurs, entre autres la temperature et l'humidite • Ainsi d1apr~s Giard(l894), la
deshydratation progressive diminue les echanges respiratoires et en m3me temps tous les phenom~nes vitaux. Elle peut aboutir
a
un etat d'anhydrobiose ou vie latente par desseohement,dont le sommeil estival de nombreux animaux n•est qu'une variete remarquable, Caldwell(l925) a faita
peu pr~s les memes oonsta-tations pour les inseotes. De leur cSte~ Kayser et Ginglinger(1927) ont prouve que chez nombre d'animaux la valeur du quotient respiratoire est fonction de la temperature et augmente avec
ell e. Dans de tels cas, la reduction des echanges respiratoires ne serait qu'un phenomena se produisant durant des diapauses
causees soit par la temperature~ soit par la secheresse.
4. FACTEURS PHYSIQUES OU RXTERNES
a. Temperature
Il n'y a probablement pas de facteur qui ait ete plus etudie que la temperature. Ball (1901) a remarqua que chez Papilio machaon ce n'est pas necessairement le froid ou la chaleur qui apportera des modifications dans la diapause mais des variations de temperature dans les conditions du milieu ou l'individu a passe sa vie larvairee Sanderson ( 1908) croit
que dans les climats temperes, les insectes doivent d'abord 8tre soumis au froid pour sortir de leur torpeur hibernalee Suivant Balachowsky ( 1937), les facteurs responsables de la diapause larvaire chez la pyrale du po~er sont complexes, mais il a pu briser cette periode de repos apres 31 jours en
soumettant les chenilles
a
de b~sses temperatures,- 10°C suivi immediatement d'un sejoura
des temperatu~es elevees25°U.
Dans le cas de Pieris brassioae, 11hibernation ne serait qu'un ralantissement des phenomenes vitaux sous l'in-fluence immediate de la temperature, Pioard(l923). Il en est
d'autres espeoes parfois tres rapprochees dans la classification, chez lesquels la diapause hivernale est, au moins partiellement, sous la dependance de facteurs internes, et ne peut etre compl~
tement empechee par l'action des circonstances exterieures. Le meme auteur mentionne un autre cas different, celui de Melittobia acasta, un hymenoptera nettement homodyname, c'est-a-dire que le nombre de generations est non interrompu aux temperatures optima. Par centre, la diapause, une fois commencee, ne peut etre inter-rompue par l'elevation de temperature. D'apres Picard, il sam-
'
blerait dans ce cas que tout depende du degre ther.mique auquelse sont effectues les premiers stades.
D'autres travaux interessants traitant de
l 1effet de la temperature sur la diapause sont ceux de Greel~
(1901), Dreyer (1932), Brumpt (1934), Abaloos (1935) et Volkonsky (1937).
b.- Humidite
Bien qu'attribuant la diapause
a
d'autres causes, certains auteurs, Baumberger~ Babcock, Tower, etc •• reconnaissent que l'humidite joue un role important dans ce phenomena. D'autres experimentateurs vont encore plus loin et croient que l 1humiditeest le facteur de premier ordre, sinon le seul, concerne avec la diapause. Somme on l'a vul Ginrd (1894, 190~) attribue cette periode de repos
a
une dessication progressive et le developpe-ment nor.n~l ne reprendrait qu'apres la rehydratation des individus.),1r-.rchal (1897) explique de la meme faqqn les variations dans le nombre de genere.tions chez la j_iouche de Hesse. En 1 'absence d
'hu-midite suffisante les oeufs de la sauterelle brune Locusta parda-lina demeure en diapause pour des annees et ils n'ecloront que sous 1 'influence d 'une pluie les immergeant completement.,
Lovvne-bury (1915). Chez 1;;. bete
a
patate, Leptinotarsa decemlineata, il y a d'abord dessication avant l'hibernation et celle-ci ne peut etre rompue sans le contact de l'eau., Breitenbrecher (1918)et Fink (1925). Breitenbrecher ajoute de plus qu'un type d'hi-bernation peut etre produit en tout temps au moyen de la dessica-tion., excepte aux basses temperatures, alors qu'il peut y a.voir que peu de dessication. Tow.nsend (1926) bien que considerant
1 'l:..icernation c.c)~cw un stage necessaire dans le cycle vi tal de la pyrale du pommier, croit que le facteur le plus important pour rompre cette periode de repos ohez les animaux ~- sang froid, est la reabsorption de l'eau par les tissus. Cetta reabsorption active l'aotion des enzymes et sans elle la processus enzymatique qui, lui, est neoessaire pour mettre fin
a
!'hibernation, ne peut avoir lieu.21
c. Lumiere
Des resultats tres interessants furent obtenus par Sabrosky et ses collaborateurs (1933) en soumettant les oeufs
de la sauterell~, Acridium arenosum angustum
a
!'influence de ra-diations electriques blanche et violet. l~ormallement cette sau-terelle hivernea
l'etat d'oeuf, mais au moyen de ces irradiations, ils ont pu obtenir une generation d'hiver et briser la diapause, alors que la chose etai t impossible en sownettant simplement les oeufs aux tewperatures elevees de la serre.Par ailleurs~ Hufnagel et Nabias(192B) ont montre que 11aotion du raditun n'apporte aucune modification dans le
de-veloppement des Calligrapha et HYPonomenta, bien qu'ils aient pu s'assurer que les rayons de radium traversaient les oocons
et la chitine. Ray (1937) de son cote, pretend que ni le chorion ni le cuticula de 11oeuf de la sauterelle Melanoplus
differentialis transmettent les rayons ultra-violets. Le plus grand pou.rcentage .d' eclosion qu 'il a obtenu en soumettant les oeufs apr8s des radiations d1une minute serait du
a
certaintype d'irradiation secondaire d 'une de ces couches plutot
qu'a
l'action des rayons direo-t?ement sur l'oeuf. Ray a aussiconstate que des irradiations de 1/2
a
4 heures retardaient le developpement et pouvaient m~me avoir un effet mortel. 'Jeci concorde avec les experiences de Huss·ey, Thompson et Calhoun (1927) qui ont trouve que chez les larves deDrosophila, la duree moyenne de la periode prenymphale est une fonction croissante de la duree d'irradiation aux rayons
x.
5. FACTEURS CHIMIQUES
Des 1876, Duclaux~ en se servant des oeufs du ver
a
soie du mGrier, demontrait qu'on peut provoquer une eclo-sion precoce en plongeant ceux-ci pendant 2 minutes dans l'acide sulfurique au maximun1 de concentration. D'apres Giard (1894), il y aurait dans ce cas une deshydratation brusque suivie, sansdoute, d1une augmentation des echanges respiratoires bien
supe-rieure
a
celle des animaux normaux.Plus recemment, Sautet(l933) a pu rumener une reactivation rapide des larves d'Anopheles bifurcatus en pleine asthenobiose en ajoutant une goutte d'eau d'un agent oxydant.
eau de javel ou permanganate de potassium, pour chaque 100 c.o. d'eau. Dans cette reactivation, la chaleur jouerait le rSle de mordencage, mais par elle-meme, elle est incapable de faire cesser la diapause hibernale.
Cousin (1932) dans ses etudes sur la diapause des Muscides remarque qu'un sejour proionge dans un milieu toxique cree au moyen d'interventions chimiques ou mecaniques,
22
peut entrainer une diapause durable et meme la mort des indi-vidus. Par contra si le sejour est de courte duree, les composes chimiques existant dans les milieux toxiques agiraient comme agents reactivants du developpement, de la meme faQOn par example, que les chocs thermiques.
6. LA. DIAPAUSE ET L'INTOJaCATION DE L'ORG.ANIS1JIE
Dubois (1895-l900),Bellion (1909), Child (1915), Baumberger(l917) et Roubaud (1922-1937) sont les principaux
tenants de cette theorie sur l'intoxication. Dubois et son eleve Bellion attribuent cette intoxication ou automarcose
a
l'accumu-lation de ga~ carbonique dans l'organisme. L'anhydride carbonique agiraita
la facon d'un narcotique provoquant le sommeil hiber.nal. L'emission du gaz carbonique diminue beaucoup durant la torpeur et la fin de la diapause serait le resultat d'une reaction de l 1animal contra une concentration trop forte deco
2• Suivant la dose, le meme agent provoquerait done le sommeil ou le reveil. Ce reveil, toutefois ne peut avoir lieu sans la rehydratation des tissus et, d'apres Bellion, pour l'escargot l'etat hygrometrique de l'air est le facteur externa essential de la torpeur hibernale; la temperature n'etant qu'un facteur accessoire.
Recemment la th~se de Dubois fut reprise par Krumenskii et Paikin (1939) pour expliquer l'hibernation des adultes du charenQon de la betterave Cleonus punctiventris Ger.m. Des temperatures inferieures
a
34e7°F. provoque la diapause duoharenQon mais comme de telles temperatures ne se renoontrent pas naturellement en ete alors que certains adultes demeurent en
diapause, ils ont fait des recherches sur 11influanoe de la
con-centration d'oxygene et de gaz carbonique dans l'air. Ils ont trouve que lorsque l'atmosphere oontient plus de 9.5% de
co
et moins de 8.6% d'oxygene il y a diapause. L'air present dans
le sol, cependant, contient rarement un pourcentage aussi bas d'oxyg~ne et aussi eleve de
co
2• Un tel milieu pourrait 3tre orae que par la respiration dans les cellules tres compact des pupes ou il ne peut souvent y avoir
a
peu pres pas de ventilation. La condition des parois des cellules de la pupe dans le sol seraitdone d'apres eux le facteur le plus important dans la formation d'une atmosphere provoquant la diapausee
Suivant la theorie de Roubaud la diapause serait une inactivite spontanee~ independante des basses temperatures, due
a
une intoxication hereditaire et assimilablea
des proces-sus de fatigue liesa
une conditions de surcharge de produits d'excretion dans l'organisme. Ces produits peuvent etre desurates~ des carbonates d'ammoni~que ou de calcium, ou encore de la guanine. Un stage d1activite temporaire
a
de bassestem-peratures ( ather.mobiose) ou dans une atmosph~re tres seche (anhydrobiose) est necessaire pour per.mettre aux organes d1
ex-oration de purifier l'organisme afin que celui-ci puisse repren-dre son fonctionnement nor.mal. De ces diapauses vraies(asthe-nobiose) ou spontanees, ou l'arret evolutif survient sans cause apparente• Roubaud distingue les pseudo-diapauses ou diapauses banales dues
a
des conditions adverses de milieu. Enfin oette asthenobiose spontanee en plus d1etre obligatoire etheredi-tai.re se rnanifesterait tantot regulierement tantSt au contraire
')~
elle est cyclique. Comme on le voit• cette the~ie, critiquee par plusieurs, est assez elastique pour pouvoir s'adapter
A
tous les cas.Plus reoemment, 1935, Roubaud a attribue l'hi-bernation obligatoire de la mouche Phlebotomus pappatasii
a
11influence exercee par l'organisme mater.nel. Lorsque les ovules
sont retenus dans les ovaires pendant un temps a.normal, elles s'y surchargent d'elements inhibiteurs du developpement. Les effets de oe blocage se ferant sentir ulterieurement amenant la diapause. D'apr~s lui il s'agit bien la " d'un effet de sur-charge metabolique. puisque le forcage
a
la chaleur entraine habituellement la mort. C'est la detente prolongee, assuree par le repos de !'hibernation, qui per.met aux individus asthe-niques de ranimer leur activite."7. LA THEORIE DE BODINE
Tel que deja explique, Bodine(l923-1937), fait une distinction entre !'hibernation et la diapause, la premiere etant surtout occasionnee par la temperature alors que la der-niere est inherente
a
11espece et independante de la temperaturepour son occurrence. La plupart des experiences de Bodine ont eu pour sujets des sauterelles. Pour expliquer la diapause des
oeuf's de Melanoplus, il introduit !'expression 11 diapause factors"
ou facteurs de diapause, qui peuvent etre soit chimiqaes, physico-chimiques ou encore des facteurs physiolog~que-genetiques, presents
dans l'oeuf en quantite plus ou moins constante lors de l'ovi-position. Suivant Bodine, ces facteurs "x " sont extr&mement
sensitifs aux basses temperatures, au-dessous du zero ou seuil de developpement, et peuvent 3tre compl~tement detruits ou sup-primes. Una destruction lente et graduelle ou bien une reduction
dans l'efficaoite des faoteurs de diapause
"x"
survienta
des temperatures oonstantes elevees, au-dessus du zero de develop-pe.ment, jusqu•A ce que les faoteurs de developpement " a uprennent le des sus et que le developpement normal reprenne. Il me semble que nous avons la beauooup d1expressions savantes
simplement pour expliquer que la temperature est un faoteur important dans la diapause de Melanoplus differentialis.
8. LA THEORIE DE COUSIN
Dans ce qui est probablement le travail le plus approfondi et le plus oomplet sur oe sujet, Cousin (1932) soutient
que la diapause est le resultat d'une perturbation physiologique speoifique en liaison direote ou indireote avec le sejour de l'Inseete dans des milieux defavorables.
Cousin commenca d'abord par etablir les condi-tions optima neoessaires au·developpement puis elle fit varier les conditions de milieu afin de ehereher les relations entre les fluctuations du milieu et la :mar.che du developpement. Elle deter.mina ensuite des diapauses experimentales dans tous les stages en modifiant un ou plusieurs des facteurs suivants:
temperature, humidite, conditions de l'alimentation, atmosphere asphyxique ou toxique, compression des larvas, jeGne et lumiere. De telles diapauses durerent 3 ou 4 mois et dans certains cas persisterent meme dans des conditions optima.
D'autres experiences aveo le meme material, en grande partie Lucilia sericata, demontrent que les conditions brisant la diapause sont generalement liees tres etroitament avec oelles qui l'ont determinee. AinBi une diapause causee par une secheresse peut 3tre rompue par un sejour d ans un milieu ou 11air est sature de vapeur d 'eau. Mais lorsque la
duree de la diapause a
ate
assez longue ou que les facteurs qui l 1ont provoque ont agi de fa9on tr~s intense, cetterela-tion entre la cause de la periode de repos et la reprise du developpement n'est pas aussi evidente. Pour obtenir la rupture de la diapause, il peut 3tre alors necessaire d1
ad-joindre au facteur qui 1 •a ocoasionnee, d 'autres provoquant certains chocs et jouant le role d'excitant. Ceei illustre bien l'importance de conna1tre le passe des sujets sous ob-servation de m3me que lea conditions optima de developpe.ment. Plusieurs auteurs dans leurs recherches ont essaye d'etablir des conditions striotement normales, mais leurs fluctuations sont generalement plus nuisibles que bienfaisants
a
l'insecte.Mellanby (1938) croit, comme Cousin, que chez Lucilia sericata la diapause est occasio:nnee par des condi-tions de milieu defavorables. De plus cependant, il partage
!'opinion de wtgglesworth qu'il
y
a diapause parce que dans de tels milieux le " corpus allatum " ou anneau de Weismann ne fonctionne pas. D'apres lui les traitements varies employes par Cousin seraient des agents stimulateurs de cet organe, etil en serait de meme pour les divers chocs appliques par Roubaud. De cette revue de la litterature il resulte done qu'une foule d'hypotheses~ dont certaines sont maintenant desuetes, ont ete avancees pour expliquer la diapause chez les vertebras en general et les insectes en particulier. Les auteurs modernes sont ganeralement d1accord pour ne voir dans la soi-disa.nt
glande d'hibernation rien d1autre qu•une forme de tissu adipeux.
Les hormones~ tres probablement, influencent le developpement des insectes, mais il est loin d•etre prouve que leur absence ou leur reduction occasionne la diapause.
Il est vrai que lorsqu1une grande regularite
dans le developpament a existe depuis des annees, voir meme des siecles~ quelqu'un peut difficilement s•attendre
a
changer cette regularite aussit6t qu'il modifia le milieu. Ceci cepen-dant ne semble pas suffisant pour nous justifier d1attribuerla diapause
a
un rhythme heriditaire.En
effet, il a ete prouve par plusieurs qu'un tel rhythme peut etre modifie. Ainsi, pour ne citer qu'un cas parmi une multitude d1autres•Brumpt (1934) a demontre, que chez Lucilia bufionivora, la diapause qui semblait obligatoire est due en partie
a
la selection naturelle entrainant la disparition des individusn'ayant pu subir cette adaptation necessaire dans des condi-tions climatiques saisonni~res determinees. En effet, il a pu rompre prematurement cette diapause par la seule elevation
de la temperature ou par l'action combinee de la temperature et de l'humidite.
Il semble bien prouve qu'un ou une combinaison de facteurs physiques, tel la temperature• ou encore des facteurs chimiques sont dans certains cas responsable de la diapause, mais ce serait une erreur, en partant d'un de cas cas, de generaliser et attribuer ce phenomena
a
un seul de ces facteurs. Il a aussi eta signale que certains facteurs contrairea
l'insecte peuvent causer une espece d'intoxication contra laquelle l'individu reagit en entrant en diapause; oe n'est pas alorsa
l•intoxication ell~m3me mais au milieu toxique que l'on doit attribuer la diapauseeEnfin pour certains auteursJ la diapause pourrait etre le resultat d1un ensemble de circonstances
incluant
a
peu pr~s toutes les hypotheses emises. Ainsi Richards&
Miller (1937) voient dans la diapause un pheno-mene obligatoire, du soita
la presence d'une hor.mone inhi-bitrice, soit ~ l'affaiblissement temporaire d1hor.mones decroissance. De plus, elle pourrait se ptuduire normalemant en raison de l'heridite, ou elle pourrait ~tre produite,soit dans l'oeu£ par des facteurs agissant dans le sang de la m~re,
soit chez l'individu lui-meme, apres que l'oeuf est pondu, par l'action de certains facteurs de milieu.
Les resultats obtenus par tous oeux qui ont etudies oette question semble.nt s'expliquer beauooup plus
faoilement et aveo beaucoup plus de vraisemblanoe
a
la lumiere de la theorie de Cousin,a
savoir : que la diapause serait le resultat de sejours dans des milieux defavorables au deve-loppement de l'inseoteo Aveo Cousin, je orois que ohez tout inseoteou
il y a diapause, on pourra graduellement eliminer oelle-ci en elevant les individus dans des condi-tions optima.CH.APITRE II
MA.TERIEL ET METHODES
1. PRO'iTENANCE DES INDIVIDUS
Pour mes etudes preliminaires, saison 1938-1939, je me suis servi de cocons provenant des echantillons
reQUS au cours de 1 'ete en rapport avec notre reconnaissance relative aux insectes forestiers. Gobeil (1939). Nous avions des specimens nous venant d'a peu pras taus les comtes de la province ( tableau 1) et pour chaque individu une fiche don-nant entre autre details son identite, sa planta nourrici~re
et la date de formation du cocon. J1esperais ainsi pouvoir
montrer l'action du climat, de l'alimentation et des conditions saisonnieres sur la duree de la diapause, mais ce me fut impos-sible; les renseignements que je possedais etant de nature trop generale. De plus, mes sujets venant de differentes localites et ayant passe une partie de leur vie dans leur milieu natural., oertains facteurs particuliers sur lesquels je ne savais rien du tout auraient fort bien pu 3tre la cause premiere de ces longs sommeils. C'est la seule fa~on d'expliquer certaines reactions apparemment contradictoires obtenues en 1938-39.
Tableau 1
PROVENAN6E DES ESPECES ETUDIEES EN 1938-39 (Nombre de specimens)
Comtes Diprion Pikonema Pikonema Neodiprion po1ytomum alaskensis dimmockii abietis
~--Abitibi 162 60 52 24 .A.rgen~uil 46 1 1
•
Beauce 131 1•
•
Belleohasse 412 3•
•
Berthier 95 7 21 1 Bonaventure 673 6 25 2 Cr.L8.r 1 evoix 660 6 7 5 Chicoutimi 399 12 22 2 Compton 15 1 ,4•
Dorchester 297 1 4•
Frontenac 171 9 3•
Gasp~-Nord 2863 22 5•
Gaspe-Sud 2327 32 20 5 Ge.tineau 2 29 13 8 Joliette 20 4 8 1 Kamouraska 2308 123•
1 Le-belle 2 138 12 38 Lac- st-J ean 122 2 5•
Le.:violette 136 218 8 25 Levis•
•
•
•
L 'Islet 646 14 1 46 Lotbini~re 128 1•
0 Hs.skinonge 24•
•
•
IvTatane 647 9 4 1 Ms.tapedia 9706 409 24•
Megantic 31•
•
•
IL<>ntcalm 53 1•
66 Montmagny 19 2•
•
Pap in eau 45 28 18 46 Pontiao 4 11 21 31 Portneuf 690 15 32 5 Quebec 893 9 22 8 P..imouski 6046 30 33 10 Riv-du-Loup 583 2 4•
Roberval 61•
4•
Saguens.y 391 70 73 2 St-Maurioe 48 7 1•
Temisoamingue 43 1 3 6 Temisoouata 1064 15 37 31 Terrebonne 40•
•
•
Wolfe 41 34 3•
32044 1333 490 36332
Nematus erichsonii 1082 230•
•
•
•
•
37•
•
•
1•
895•
•
783 772 ••
•
••
2•
•
•
10•
294 49 319•
•
•
903 31 264 8•
•
•
5680Par ailleurs 11heterogenite des eohantillons
constituant mes lots a quelquefois invalide mes resultats et~
a
moins qu'elle fut tres signifioative~ il n•etait pas toujours possible d'attribuer aux traitemants la difference de develop-pement observee entre deux lots. En effet, il y avaitd'au-tres causes de variations~ regionales~ alimentaires, ou peut-etre m&me genetiques qui n'avaient pas ete eliminees et qui pouvaient avoir sur les sujets una plus grande influence que les traitements experimentes. Il m'est arrive par example d'avoir des pourcentages d1emergence tres differents pour des
lots places dans des conditions presqu1identiques. Sur
exa-men des eohantillons for.mant cas lots, j'ai constate que plusieurs individus, de volume inferieur
a
la moyenne. fai-saient partie d'eohantillons oontenant un nombre considerable de larves elevees dans des bocaux trap petits, d1ouinsuffi-sance de nourriture. Ces larvas etai ent evidemm.ent mains resistantes que les larves nor.males du deuxieme lot, de la un plus fort pourcentage de mortalite et mains d'emergence.
En 1940, j'ai reduit les causes incontro-lables de variations en me procurant un stock aussi homogene que possible. Taus les cooons de D.polytomum
(so.ooo)
furent---reouaillis
a
la m3me date, le 26 aoat et au meme endroit, dans un peuplementmele, (
ep~nette~ sapin et bouleau),a
Ste.Florence. Co.Matapediae Quant aux 25,000 cocons de N.erioh-soni, ils furent ramasses entre le 26 aout et le ler septembre dans le canton Thorne, comte de Pontiac.
2. ESPECES ETUDIEES
a. Diprion polytomum
Pour una meilleure comprehension des
expe-l~ences qui vont suivre quelques mots sur le cycle £•polyto.mum sont necessaires. Auparavant1 je dois d'abord attirer !'at-tention sur la classification de Benson {1939) qui inclue D.polytomum dans son nouveau genre Gilpinia separe de Diprion
surtout par le developpement des "cenchri" du meso- et meta-scutellum. Les earacteres invoques par Benson justifient peut-3tre !'erection de ce nouveau genre~ mais comme l 1usage
de ce nom est encore plutot restraint, j'ai cru preferable d'utiliser ici Diprion polytom~ nom sous lequel l'espece
est maintenant connue de tous les forestiers de ~ebeoo Cette tenthrede apparemment introduite d1Europe et trouvee dans la Gaspesie
a
l'etat epidemique en1930 ( Baleh
&
Simpson 1932) ne s'attaque qu'a l'epinette. L'insecte hiverne dans la mousse ou 11humusa
11etat larvairedans un eocon brun. Au debut de l'ete~ un poureentage plus ou moins grand des individus se transforment d 1abord en pronymphe
puis en n~he et finalement en adulte dont la plupart sont des femelles. En effet, les ~les sont tres rares et la reproduction est parthenogenesique. L1adulte est plutot
indolent et bien qu'il puisse volar sur de longues distances~
il ne prendra son vol que par des journees ensoleillees~
lorsque la temperature est relativement elevee. Comme on le verra au ohapitre VII,cette caracteristique est importante en
rapport avec la th~se de Roubaud sur la. diapause et l'intoxi-cation des oeufs. Ceux-ci sont pondus
8.
r ai son de un par feuille sur le vieux feuillage. L'eclosion de la larve se produit de 8a
12 jours plus tard; elle se nourrit de prefe-rence sur le vieux feuillage ne s'attaquanta
la nouvelle pousse que plus tard dans la saison, et seulement lorsqu'il ne luireste rien d1autre
a
sa portee. Au cours de sa vie larvairel'insecte mue 5 fois et passe par 6 stages, alors qu'ayant atteint pleine maturite, il se laisse choir de 1 'arbre pour tisser son oocon dans la mousse. Entre le sixieme stage lar-vaire, qui ne dure que quelques jours, et la pronymphe, cer-tains auteurs reconnaissent un autre stage, celui d'eony.mphe, forme sous laquelle l'insecte hiverneraite Dans oe travail l'eonymphe est tout simplement designe par le mot larvae
L'emploi de ce terme ne peut preter
a
confusion puisque cette etude ne porta que sur le developpementa
l'interieur du cocon comprenant trois stages,a
savoir: la larva ou eonymphe, la pronymphe et la nymphe.Dans le nord de la province il n'y a qu'une generation par annee alors que dans le district de Montreal et des Cantons de 11Est on peut avoir 2 generations et 3 dans
36
Dans la Gaspesie des adultes sont eolos de cocons en diapause depuis 6 ans, la duree de cette lethargie
variant suivant les regions et les annees. E'apr~s Baleh (1939) le pouroentage moyen d1emergenoe par annee
approohe-rait 20% dans la Gaspesie alors que dans le centre du Nouveau-Brunswick il serait d'environ 70"/o. Pour une m3me localite, ce pouroentage varie beaucoup d1une annee
a
11autre; dans le comtede Kwmouraska seulement 2% des larves hibernantes ont donne naissance
a
des adultesa
11ete 1934.Au
meme endroit pourles 3 annees qui suivirent, le pouroentage d'emergenca a varie de 12
a
281oe Suivant Baloh, la temperature et 1 1humidite)seraient des faoteurs de premi~re importance pour la rupture de la diapause. Un sejour d'environ 3 mois
a
des tel!g?eratures au-dessous du s euil de developpement serait neoessaire pour obtenir le maxumum d'emergence. Ce maximum serait de plus influence par le degre d1humidite et le contact de l'eau.Sous elevages eontr8les, Baleh a obtenu pour 21 generations
conseoutives une lignee sans diapause. D'un autre c8te, les descendants de femelles provenant de la Gaspesie montrent une forte tendance pour l'hiber.nation et,
a
!'exception d'un cas, il a eta impossible de l'eliminer apres la premi~re ou la deuxi~me generation. Tout en admettant !'importance de la temperature et de l'humidite, Balch croit done que ohez Depolytomum la diapause est hereditmre et les variationsb. Nematus erichsoni.
Les insectes hivernant dans le sol ont gene-ralement surabondance d'humidite. L'habitat du cocon de la mouche
a
scie de l'epinette~ bien que souvent sature, est, cependant. relativement sec, surtout lorsque la larva s'est developpee sur l'epinette blanche dans des peuplemants meles ou le sol est ge-neralement bien draine. Par contra le oocon de la mouchea
sciedu mel~ze etant dans un milieu presque toujours sature d'eau et souve.nt meme en contact avec 11eau. cette esp~ce est bien
repre-santatives des tenthredes hiber.nant en milieu excessive.ment h~dee Soumis
a
l•aotion des memes agents• cat insecte devrait done dans maints cas reagir differemment de ~.polytomum. C1est pourquoi~ en1940~ la plupart des experiences faites sur Diprion. furent repe-tees avec Nematus erichsoni.
La mouche
a
scie du meleze est una autretenthr~de indigene d1Europe. Sa presence fut constatee en
Ame-rique pour la premiere fois dans l'Etat du Massachusetts en 1881 par Sargent ( Hewitt.l912). Depuis la mouche
a
scie du meleze s'est revelee l'insecte le plus nuisible du mel~ze. Son cycle vital ne differe pas sensiblement de oelui de Diprion. La larva hiver.ne aussi dans son cocon sous la mousse. Ici encore lesmales sont peu frequents et la reproduction est parthenogenesique. Les oeufs sont pondus en rangees dans des fentes taillees sur les pousses terminales et lea larvas se nourrissent
eaolusive-ment des feuilles du meleze.
Il n'y a dans la litterature que tres peu de details sur la duree de la diapause de la mouche
a
scie du'1'
me eze. Parmi les nombreuses publications consultees, je n'en ai trouve qu'une seule, Baird (1922), ou il en soit fait mention. Lors de ses etudes sur le controle natural de Nematus erichsonii dans le Nouveau Brunswick, oet auteur remarqua qu'en 1921 environ 25% des larves etaient encore dans leur cocon apres un an d'hi-bernation. Depuis
1plusieurs annees de manipulation de oocons
de oette espeoe pour la propagation de parasites ont per.mis ~ M.Baird ( lettre du 12 nov. 1940) de constater que d'une annee
a
l 1autre et pour une meme looalite, il y a des variationscon-siderables dans la duree de l'hibernation. Generalement oepen-dant un tres petit pourcentage des larvas, moins de 1%, dameurent en lethargie pour plus d'un hiver, bien qu'il soit arrive que ce pouroentage atteigne 25% et que quelques larvas fussent encore
vi vantes dans leur cocon ~r~s 2 ans.
Les observations faites par J.J.DeGryse (lettre du 24 oct. 1940)
a
Indian Head de 1923a
1925, bien que de nature tres genera1e, indiqueraient que dans 1'6uest du Canada la diapause peut durer deux et meme trois ans.D'apres nos propres records (Genest, rapport Annual 1939) un tres faible pourcentage des larvas resteraient dans leur oocon pour plus d'un an• Sur un total de 1128 oocons ramasses entre le 11 mai et le 30 juin 1938, la majorite
a
St.Fabie.n,Co.Rimouski, 44% donnerent naissanoea
des adultesdurant l'ete alors que 17.4% etaient parasites. A 1 'automne 1 sur la balance des 435 cocons~ il n'y en avait que 9 contenant apparemment des larves vivantes, et au printemps 1939 lorsque ces cocons furent ouverts, il n'y avait qu'une seule larve vivante. Sur 326 cocons recueil1is au m3me endroit entre le
2.7 mai et le 12 juin 1939~ 46.~% produisirent des adultes et 38.2% des parasites. Le 13 aout il ne restait plus que 18 larves vivantes.
D'apr~s mes propres experiences 1940-41~ il n'y a pas de doute que certaines annees, dans certaines loca-lites, la plus grande partie des larvas subissent des diapauses qu'il est tr~s difficile de rompre. 11 est vrai que les traite-ments que j'ai fait subir a~~ individus de cette esp~ce caus~rent
dans plusieurs cas lOO% de mortalite. Il n'en reste pas mains, cependant, qu•a la fin de mes experiences j'avais encore plus d'un millier de sujets vivants et sur ce nombre je n'ai eu
moins d'une dizaine d'adultes. Pourtant les memes traitements
en 1938-39 avaient provoqu~s jusqu•a 40% d'emergence. Je ne puis expliquer cette difference de comportement qu'en
l'attri-buant aux d.onditions de milieu dans lesquelles vecurent ces individus avant leur arrivee au laboratoire, conditions ayant provoquees des diapauses tellement durables qu'il me fut, dans la majorite des cas, impossible de les briser au moyen des traitements employes.
c. Autres especes
Alors que les etudes de 1940 porterent exclusivement sur Diprion polytomum et Nematus erichsoni, quel-ques observations supplementaires furent faites en 1938-39 sur les tenthredes suiva.ntes, en plus bien entendu, des 2 especes ci-dessus mentionnees qui ont toujours consitute la plus grande partie du material. Nombre de specimens Pikonema alaskensis 662 Pikonema. dimmockii 234 Neodiprion lecontei 319 Neodiprion pinetum 108 Hylotoma pectoralis 273
'
Comme la plupart de ces especes furent reparties sur un total de 70 lots traites differemment, il va sans dire que le nombre d'individus dans chaque lot n'etait pas assez eleve pour en arriver
a
des conclusions definies, exoepte lorsqu'il s'est agi de milieux nettement defavorables.3. TECHNIQUE
a. Saison 1938-39
A la fin de l'ete 1938• les 13,500 cocons provenant de nos elevages en rapport avec notre inventaire des
insectes forestiers furent groupes en 27 lots variant de 350
a
600 oocons. Chaque lot fut constitue d1echantillons venant dedifferentes parties de la province et contenant de 1
a
10 indi-vidus places dans des fioles de 15 x 45 m.m. bouchees avec de 1 'ouate. Le ler octobre tout le mater.iel fut expediea
l'entre-pot frigorifique des Ports Nationauxa
Quebec et remise pour des periodes variant de 4a
14 semaines dans des chambresa
temperatures constantes soit: 32°F1 l5°F~ et 0°F, respectivement. Les abaissements de temperature furent graduels~ c'est-a-dire qu'a-vant leur sejour
a
0°F, par exemple~ les individus passerentDe l'Entrep8t frigorifique~ les oocons
furent expedi es au Coll~ge :Nfa.cdonald et amenes progressivement~ par bonds de 15°F ( 3 jours d'exposition
a
chaque echelon)~ aux temperatures de 60°F. 75°F~ 80°F~ 85°F, et 90°F~ dans une atmos-phere tres seche soit 20a
22% d'humidite relative dans les eh~'bres de 85°F. et 90°F~ environ 25% pour celles de 75°F~ et 80°F
0
et 35% dans la chambre de 60 F. Il est
a
remarquer qu•au cours du trajet de ·Quebeca
Ste-Anne de Bellevue { une journee) il me fut impossible de controler la temperature. On m'a assure~ ce-pendant, qu'elle osoillait entre 34 et 45°F.A leur arri vee au I·J1aoDonald, ohaque lot fut subdivise en 2 ou 3 sous-lots de 150
a
200 cocons et soumis aux temperatures de nymphose oi-haut mentionnees. Auparavant•cependant, un echantillon de 50 cocons par lot fut preleve et analyse afin de determiner le pouroentage de mortalite au oours de !'hibernation, oe pouroentage etant ensuite applique aux
sous-lots oonstituant le lot original.
Finalement, apr~s un sejour de 2 mois
aux temperatures de nymphose1 tous les oooons furent ouverts.
et le pouroentage de mortalite et de developpe.ment en incubateur fut calcule. Les chiffres obtenus furent alors utilises pour
estimer l 1aotion de differents milieux sur les individus.
b. Saison 194Q-4l
En 1940, la technique employee et les appareils utilises furent de beaucoup superieurs
a
1938. D1a-bord les variations dues
a
l'origine des individus furent re-duites au minimum en obtenant mon stock au mame endroit et versla meme date. A leur arrivee
a
Quebec, les oooons de ~.polyto mum fUrent divises en lots de 125 cocons et oeux de Neerichsonien lots de 110 oocons, places dans des tubes de verres de 22x 95 mm., dans lesquels l'aeration fut assuree en bouchant les extremites avec de la mousseline ( Pl.I, fig.A .)
Le ler septembre tous ces lots commen-caient leur sejour aux basses temperatures dans notre refri-gerateur General Electric