Analyse critique des techniques d'estimation du nombre d'unites motrices.
Texte intégral
Figure
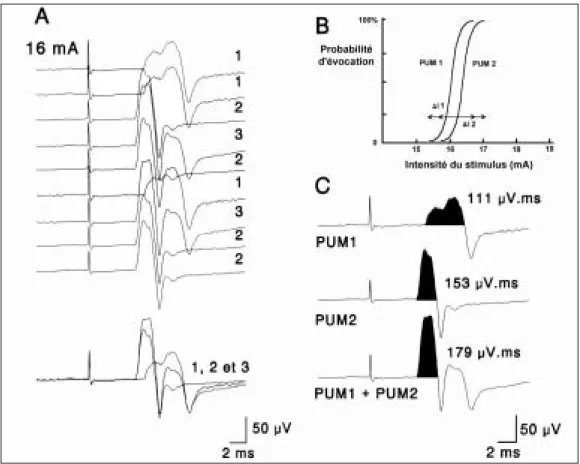
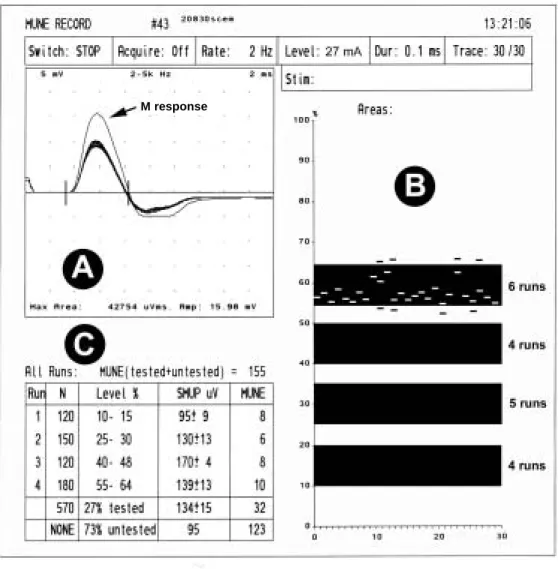
Documents relatifs
II./ Le vieux Yara a laissé son héritage dans un coffre dont la combinaison comporte les cinq chiffres x, y, z, t et h dans cet ordre, du système décimal.. Il a mentionné sur son
Elle est d’autant plus importante que la masse de la charge est grande et s’oppose à la mise en mouvement. Elle est caractérisée par le moment d’inertie J, qui s’exprime en
Le soumissionnaire remet, comme pièce constitutive de son offre, un document par lequel il marque son engagement à mettre en œuvre
Dans un circuit en série et dans une branche d'un ciruit en dérivation, plus on met de résistances les unes à la suite des autres et plus l'intensité diminue dans le circuit (ou
Montrer que les points A, B et C ne sont pas alignés. b) Donner une équation cartésienne du plan (ABC).. En préciser les
[r]
Il sera tenu compte de la rédaction dans la note finale.. 5- Donner un exemple algébrique de fonction qui admette ce tableau
Ancien élève de VEcole Polytechnique et de l'Ecole Supérieure d'Electricité de Paris. Licencié
