Évaluation de l’impact de l’ajout de flocons de
macroalgues sur le développement et la bioactivité d’un
fromage fonctionnel de type Camembert
Mémoire
Attara HELL
Maîtrise en Sciences et technologie des aliments
Maître ès sciences (M. Sc.)
Québec, Canada
Évaluation de l’impact de l’ajout de flocons de
macroalgues sur le développement et la bioactivité d’un
fromage fonctionnel de type Camembert
Mémoire
Attara HELL
Sous la direction de :
Lucie Beaulieu, directrice de recherche
Steve Labrie, codirecteur de recherche
iii
Résumé
Les macroalgues Palmaria palmata et Saccharina longicruris sont riches en nutriments et contiennent des molécules antioxydantes ainsi que des inhibiteurs de l’enzyme de conversion de l’angiotensine (ECA). C’est également le cas du fromage de type Camembert. Cependant, l’impact de la combinaison de ces deux aliments sur la bioactivité globale est inconnu. Une analyse de la composition nutritionnelle des algues montre que P. palmata était la plus riche en protéines et en sucres totaux. S. longicruris avait un contenu plus élevé en fibres totales, minéraux totaux, sodium, potassium et lipides. Les capacités antioxydante et inhibitrice de l’ECA de l’extrait soluble > 1 kDa de S. longicruris étaient supérieures à celle de P. palmata. Trois différents traitements de fromages modèles ont été étudiés : un contrôle (CC), un modèle contenant 2 % de P. palmata (C2PP) et un autre contenant 2 % de S. longicruris (C2SL). Durant l’affinage (20 jours), l’évolution du pH du caillé des trois traitements était significativement similaire, partant de 4,89 et terminant à 6,77. La même tendance a été observée pour la capacité antioxydante (ORAC), débutant de 0 pour finir à 41,28 mmol TE/g de fromage. La capacité inhibitrice de l’ECA des trois traitements était significativement similaire au jour 0 (13,20%) et au jour 20 (58,27%). Par contre, C2SL n’avait pas la même courbe d’évolution que CC et C2PP. Ces résultats ont permis de valider la richesse nutritionnelle et des bioactivités des macroalgues. Aucun changement n’a été observé sur le développement et la bioactivité finale des deux fromages aux algues.
iv
Abstract
Seaweeds Palmaria palmata and Saccharina longicruris have high nutritional value and contain antioxidants and angiotensin-converting enzyme (ACE)-inhibitor compounds. This is also applicable to Camembert-type cheese. However, the impact of combining these two foods on the global bioactivity is unknown. The nutritional composition of the two dried seaweeds was characterized. P. palmata had the highest protein and total carbohydrate contents. S. longicruris had the highest contents in total fiber, total minerals, sodium, potassium and lipids. The antioxidant capacity (ORAC) and ACE-inhibitor activity of S. longicruris soluble extract > 1 kDa were higher than P. palmata. Three different types of cheese were studied: cheese control (CC) without seaweeds, cheese with 2% of P. palmata (C2PP) and cheese with 2% of S. longicruris (C2SL). During ripening (20 days), the curd pH of all treatments was significantly similar, starting from 4.89 and finishing at 6.77. The same trend was observed for their ORAC values, starting at 0 and then reached 41.28 mmol TE/g of cheese. The ACE-inhibitor capacity of the three treatments was significantly similar, at day 0 (13.20%) and day 20 (58.27%). On the other hand, C2SL did not have the same evolution curve as CC and C2PP. These results have shown and validated the nutritional value and bioactivities of seaweeds. Also, at the concentration of 2% of seaweeds, no impact was observed on the development and final bioactivities of the two seaweeds cheeses.
v
Table des matières
Résumé ... iii
Abstract ... iv
Liste des tableaux ... vii
Liste des figures ... viii
Liste des abréviations ... ix
Remerciements ... x
Avant-propos ... xi
Chapitre 1 : Introduction ... 1
Chapitre 2 : Revue de littérature ... 4
2.1. Les macroalgues ... 4
2.1.1. Généralité ... 4
2.1.2. Composition nutritionnelle ... 8
2.1.3. Effets bénéfiques des macroalgues sur la santé ... 14
2.1.4. Digestibilité ... 17
2.1.5. Produits alimentaires et aliments fonctionnels additionnés de macroalgues 18 2.2. Le fromage ... 19
2.2.1. Généralité ... 19
2.2.2. Différentes variétés de fromages ... 19
2.2.3. Fabrication du fromage ... 20
2.2.4. Les caillés modèles ... 23
2.2.5. Défauts de contamination du fromage ... 23
2.2.6. Rôles des constituants du fromage lors de l’affinage ... 24
2.2.7. Bioactivités ... 28
2.2.8. Fromages fonctionnels ... 32
2.3. Fromages aux algues ... 33
Chapitre 3 : Hypothèse et objectifs ... 35
3.1. Hypothèse de recherche ... 36
3.2. Objectifs spécifiques ... 36
Chapitre 4: Evaluation of the impact of addition of seaweed flakes on the development of bioactivities in functional Camembert-type cheese ... 37
Abstract ... 38
vi
Materials and Methods ... 41
Chemicals and Reagents ... 41
Biological Materials ... 42
Nutritional Composition of Seaweeds ... 42
Soluble Seaweed Extracts (SSE) ... 43
Soft Cheese Model Curd Production (SCMC) ... 43
Seaweeds Flakes Selection for Treatments ... 44
SCMC Preparation and Ripening Conditions ... 44
SCMC Water Activity (Aw) ... 45
Water-Soluble Extract (WSE) from SCMC ... 45
Angiotensin-Converting Enzyme (ACE)-Inhibitory Activity ... 45
Oxygen Radical Absorbance Capacity (ORAC) Assay ... 46
Statistical Analysis ... 47
Results and Discussion ... 47
Nutritional Composition of Seaweeds ... 47
Seaweeds Extracts and Detection of Bioactivities ... 49
Soft-Cheese Model Curds (SCMC) Water Activity (Aw) ... 51
The pH Evolution in Core of SCMC During Ripening ... 52
Detection of Bioactivities in SCMC During Ripening ... 53
Conclusion ... 57
Acknowledgments ... 58
Chapitre 5 : Discussion et conclusion générales ... 59
5.1. Discussion générale ... 59
5.1.1. Retour sur l’hypothèse de la recherche ... 59
5.1.2. Limites des méthodes expérimentales ... 60
5.1.3. Défis technologiques ... 61
5.1.4. Défis du projet de recherche global ... 62
5.2. Conclusion et perspectives ... 64
Annexes ... 65
Fromages aux algues commercialisés ... 65
Photos des prototypes des fromages aux algues ... 68
vii
Liste des tableaux
Tableau 1. Composition des acides aminés de différentes algues et d'autres aliments ... 10 Tableau 2. Différentes bioactivités des produits laitiers fermentés et leurs protéines
précurseures ... 29 Table 3. Nutritional composition of seaweeds in percentage of dry weight (% w/w dry
weight). ... 47 Table 4. Protein contents and bioactivity of soluble seaweed extracts (> 1 kDa). ... 49 Table 5. Aw of SCMC without ripening starter cultures. ... 51 Tableau 6. Différentes variétés de fromages aux algues commercialisés et leurs
viii
Liste des figures
Figure 1. Description de la filière subflottante utilisée pour la culture des laminaires ... 6 Figure 2. Saccharina longicruris (a) et Palmaria palmata (b) ... 7 Figure 3. Structures de phlorofucofuroeckol A, dieckol et dioxinodehydroeckol isolés
d’Ecklonia stolonifera ... 13 Figure 4. Mécanismes potentiels des phlorotannins sur l'ECA et les radicaux libres ... 16 Figure 5. Variétés des fromages classés selon leur méthode de coagulation et d'affinage
et leur texture. ... 20 Figure 6. Protocole de production de fromage à pâte molle et à croute fleurie de type
Camembert ... 22 Figure 7. Contribution du sel pour la conservation, la sécurité et la qualité générale du
fromage ... 25 Figure 8. Gradient des composés responsables de l’alcalinisation du caillé du fromage
Camembert ... 27 Figure 9. Évolution de l'activité inhibitrice de l'ECA de différents fromages à pâte ferme et
semi-ferme d'origine suisse à partir d’extraits hydrosolubles. ... 31 Figure 10. pH evolution of the core curds during ripening ... 53 Figure 11. ACE-inhibition of curds during ripening ... 56 Figure 12. Oxygen radical absorption capacity (ORAC) of curds during ripening expressed
in mmol TE/g of curd (2 mg/mL). ... 57 Figure 13. Fromage prototype avec Palmaria palmata ... 68 Figure 14. Fromage prototype avec Saccharina longicruris ... 68
ix
Liste des abréviations
A Absorbance
AAPH 2,2′-azobis(2-methylpropionamidine) dihydrochloride ACIA Agence canadienne d’inspection des aliments
Aw Activité de l’eau
CC* Fromage modèle témoin sans ferments d’affinage CC Fromage modèle témoin
C2PP* Fromage modèle contenant 2% de Palmaria palmata sans ferments d’affinage
C2PP Fromage modèle contenant 2% de Palmaria palmata
C2SL* Fromage modèle contenant 2% de Saccharina longicruris sans ferments d’affinage
C2SL Fromage modèle contenant 2% de Saccharina longicruris CN Caséines
DPPH 2,2-Diphenyl-1-picrylhydrazyl DW Dry weight (poids sec)
ECA Enzyme de conversion de l’angiotensine (ACE en anglais) EPA Eicosapentaénoïque
FAO Food and Agriculture Organization of the United Nations (Organisation des Nations unies pour l’alimentation et l’agriculture)
GAE Équivalent d’acide Gallique HHL N-hippuryl-His-Leu
IC50 Concentration inhibitrice médiane N.D. Not detected (Non détecté)
ORAC Oxygen Radical Absorbance Capacity (Capacité d'absorption des radicaux libres)
RH Relative humidity (Humidité relative)
SCMC Soft cheese model curd (Caillé modèle de type pâte molle) SD Standard deviation (Écart-type)
TE Trolox equivalent (Équivalent Trolox) w/v Weight per volume (Poids par volume) w/w Weight per weight (Poids par poids)
x
Remerciements
Pour commencer, je tiens à remercier ma directrice de recherche, Dre Lucie Beaulieu, qui m’a permis d’entreprendre un projet de recherche qui me passionne énormément. Son encadrement, sa disponibilité et sa patience m’ont permis d’être un meilleur scientifique. Avoir travaillé avec son équipe m’a également permis d’explorer de nouveaux domaines de la science et d’étancher ma soif d’apprendre et ma curiosité. Aussi, je suis très reconnaissant de sa confiance en mes capacités et de m’avoir laissé réaliser l’aboutissement de mon projet, c’est-à-dire la production pilote des fromages dans une fromagerie commerciale.
Je voudrais également remercier mon codirecteur, Dr Steve Labrie, qui m’a guidé et aidé à approfondir mes connaissances dans le domaine fromager. De plus, son encouragement et son soutien à produire les prototypes de fromages aux algues ont amplifié davantage ma motivation à réussir le projet. De plus, ses conseils sur l’organisation des idées de travail m’ont permis d’atteindre les objectifs.
Je souhaite spécialement remercier Diane Gagnon pour son temps, ses conseils, son support et son encouragement, tout au long de mes expérimentations. Aussi, je tiens à remercier Marie-Hélène Lessard pour son aide et qui m’a montré les bonnes méthodes de travail. Également, je voudrais remercier Céline Paquin, Mélanie Martineau, Pascal Lavoie et Pierre Côté pour leur soutien technique. Aussi, Frédéric Lehance qui a été toujours présent pour mes petits soucis techniques et à mettre la joie dans le laboratoire.
J’aimerais remercier les organismes subventionnaires, le Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ) et la Fondation INITIA. Également, je souhaite souligner la coopération de nos partenaires pour ce projet de recherche soit l’INAF, Merinov et la Fromagerie des Basques.
Finalement, je tiens à remercier mes amis et collègues de travail qui ont toujours été là pour m’encourager. Puis, je désire simplement dédier toute ma gratitude à ma famille qui a fait de grands sacrifices afin que je puisse réaliser mes rêves et de franchir ce niveau d’étude.
xi
Avant-propos
Les travaux de ce mémoire de maîtrise ont été réalisés dans le cadre du projet de « Développement de fromages fonctionnels renfermant des peptides bioactifs de macroalgues d’origine québécoise » soutenus par le programme Innov'Action agroalimentaire du MAPAQ.
Le chapitre 1 correspond à la mise en contexte et à l’introduction du projet afin de souligner la pertinence de ce projet. Par la suite, au chapitre 2, une revue de littérature a été réalisée sur les caractéristiques nutritionnelles et les bioactivités des macroalgues, ainsi que ceux des fromages. Au chapitre 3, l’hypothèse et les objectifs de recherche y sont présentés.
Le chapitre 4 présente l’article scientifique rapportant la méthodologie et les résultats obtenus. Cet article, ayant comme titre «Evaluation of the impact of addition of seaweed flakes on the development of bioactivities in functional Camembert-type cheese», est actuellement en préparation pour être soumis au journal scientifique International Journal of Food Science and Technology. M. Attara Hell est l’auteur principal. Les coauteurs, Dre Lucie Beaulieu et Dr Steve Labrie, ont contribué à l’obtention des fonds de recherche et la supervision des travaux des corrections de l’article. Ils ont été également la directrice et le codirecteur de ce projet de maîtrise et ont offert l’encadrement nécessaire pour la réussite du projet.
Finalement, le chapitre 5 propose une discussion générale sur les résultats obtenus, ainsi qu’une conclusion et les perspectives du projet. Sont présentés à la fin du mémoire, les annexes contenant une liste des fromages aux algues commercialisés et des photos illustrant les fromages prototypes aux algues produits lors du présent projet, suivi de la bibliographie du mémoire et de l’article.
1
Chapitre 1 : Introduction
Les macroalgues sont des aliments qui suscitent de plus en plus d’intérêt. Leur production mondiale ne cesse de croître. Entre 2005 et 2014, elle a plus que doublé pour atteindre 27,3 millions de tonnes annuellement, ce qui représente une valeur économique mondiale de 6,4 milliards de dollars américains. La production mondiale provient essentiellement de culture de macroalgues en provenance d’Asie. La Chine et l’Indonésie sont les principaux producteurs avec 85,7% de la part mondiale (FAO, 2016). La consommation de ces macroalgues est également concentrée en Asie. À titre d’exemple, au Japon, la consommation moyenne de macroalgues est de 1,9 kg/an par habitant (Matsumura, 2001). Or, les pays occidentaux s’intéressent de plus en plus aux macroalgues (Hein, 2016; Mouritsen, 2012; Mouritsen et al., 2012; Rioux et al., 2017).
Traditionnellement, l’industrie alimentaire occidentale s’est surtout intéressée aux polysaccharides extraits des macroalgues utilisés comme ingrédients technologiques (Hernandez-Carmona, 2013; Mabeau & Fleurence, 1993). Aujourd’hui, la consommation des macroalgues tend à changer, car on les utilise de plus en plus comme aliment entier ou comme ingrédient. Une émergence de nouveaux produits développés à base de macroalgues marque donc les nouvelles tendances alimentaires (Rioux et al., 2017). À titre d’exemple, des produits comme des pâtes entièrement faites d’algues sont disponibles sur le marché (World Food Innovations, 2017). Plusieurs groupes de recherche se penchent aussi sur l’incorporation de macroalgues ou de leurs extraits solubles bruts dans des aliments, comme le pain, le yogourt, le jambon ou même la crème glacée (Barbieri et al., 2016; Fitzgerald et al., 2014b; Mouritsen et al., 2012; Prabhasankar et al., 2009; Tavares Estevam et al., 2016). Ces dernières recherches avaient pour but de développer de nouveaux aliments avec des saveurs typiques ou d’en améliorer les bienfaits sur la santé, afin de créer des aliments fonctionnels. En effet, les macroalgues sont riches en nutriments et contiennent notamment une quantité considérable de fibres, de minéraux et de protéines (Mabeau & Fleurence, 1993; MacArtain et al., 2007; Rioux
2
et al., 2007; Tibbetts et al., 2016). De plus, plusieurs études ont démontré les bioactivités des algues, qui seraient bénéfiques sur la santé humaine par leurs effets antioxydants, anticancéreux et antihypertenseurs potentiels (Beaulieu et al., 2015; Boisvert et al., 2015; Hamed et al., 2015; Liu et al., 2015; Plaza et al., 2008).
Le fromage est un aliment qui est déjà bien établi dans la culture occidentale. Riche en nutriments, il fait partie des produits laitiers qui sont suggérés dans le guide alimentaire canadien. Il est conseillé de consommer entre 2 à 4 portions de produits laitiers par jour (Santé Canada, 2011). Aussi, cet aliment fermenté est également reconnu pour sa bioactivité. Tout comme les macroalgues, les fromages contiennent naturellement des composés ayant des effets antioxydants et antihypertenseurs (Choi et al., 2012; Crichton & Alkerwi, 2014; Crichton et al., 2012; Gupta et al., 2009; Sieber et al., 2010).
La combinaison des macroalgues et du fromage est une idée originale qui permet de créer un aliment fonctionnel. Le concept d’incorporer des macroalgues aux fromages a été exploité en Europe où ils sont déjà commercialisés, notamment Le Ti Pavez aux algues (Bretagne, France) et le Carrigaline Dillisk Seaweed Cheese (Irlande). Cependant, les produits existants sont de fabrication artisanale et aucune étude scientifique n’a exploré l’impact de la présence des macroalgues sur les bioactivités globales du fromage ainsi produit.
Des études ont démontré l’amélioration de la capacité antioxydante de certains fromages assaisonnés de fines herbes comparativement à leur formulation originale. Cela a été démontré notamment pour le feta (Apostolidis et al., 2007), de même que pour du fromage bleu dans lequel l’incorporation de poudre de brocoli a été effectuée (Sharma et al., 2011). Quant aux macroalgues, l’incorporation de 4% d’hydrolysat de protéines de la macroalgue Palmaria palmata dans le pain a démontré des effets inhibiteurs de l’enzyme de conversion de l’angiotensine-I, qui est responsable de l’hypertension artérielle (Fitzgerald et al., 2014b). Ces résultats prometteurs suggèrent une possible synergie de la combinaison des macroalgues et du fromage pour en faire un « super aliment » ou un aliment aux propriétés nutritionnelles améliorées.
3
Le présent projet vise à caractériser l’impact de l’ajout de flocons de macroalgues au fromage de type Camembert. Afin d’y arriver, la composition nutritionnelle des macroalgues P. palmata et S. longicruris sera caractérisée, ainsi que la mise en évidence des activités biologiques de leur extrait aqueux. Par la suite, le pH des caillés modèles de type Camembert incorporés de flocons de macroalgues et leurs bioactivités seront mesurés tout au long de l’affinage.
Cette étude est un premier pas vers la compréhension de l’impact de l’incorporation des macroalgues dans le fromage de type Camembert. Ceci permettra également de développer un nouveau type de fromage en Amérique du Nord. La création de fromages contenant des macroalgues faits d’ingrédients produits et récoltés au Québec permettrait aux fromagers québécois de se démarquer des produits importés de l’étranger dont le volume est estimé à près de 22 000 tonnes par année (Agriculture et Agroalimentaire Canada, 2016).
4
Chapitre 2 : Revue de littérature
2.1. Les macroalgues
2.1.1.
GénéralitéLes algues sont considérées comme les ancêtres de la vie terrestre et sont divisées en deux grandes catégories : les microalgues (unicellulaires) et les macroalgues (pluricellulaires) (Campbell & Reece, 2007). Ce sont les macroalgues qui ont fait l’objet de cette étude. Ces dernières sont regroupées sous trois espèces différentes selon la nature de leurs pigments : les algues brunes (Phéophycées), les algues rouges (Rhodobiontes) et les algues vertes (Chlorophyceae) (MacArtain et al., 2007). Leur structure biologique est plus simple que les végétaux terrestres puisque les macroalgues ne possèdent ni racines, ni tiges, ni feuilles véritables. Elles sont composées de crampon qui ressemble à des racines leur permettant de se fixer à une surface immobile, un stipe en guise de tige et de frondes servant comme surface de la photosynthèse (Campbell & Reece, 2007; MacArtain et al., 2007).
2.1.1.1. Production des macroalgues
En 2014, la production mondiale de plantes aquatiques estimée par la FAO était de 27,31 millions de tonnes. Parmi toute cette production, surtout concentrée en Asie, les macroalgues Kappaphycus alvarezii, Eucheuma spp. et Laminaria (Saccharina) japonica étaient les plus exploitées (18,65 millions de tonnes). En Amérique du Nord, la production aquacole correspondait à seulement 0,89% de la production mondiale (FAO, 2014). Elle tendra certainement à augmenter pour répondre à la demande asiatique puisque, depuis quelques années, ceux-ci sont confrontés à des problèmes importants de pollution en plus de contraintes géographiques telles que la raréfaction des zones marines disponibles (Litzler, 2012).
Les macroalgues sont exploitées de deux façons : par l’algoculture (la culture d’algues) et la récolte des algues sauvages. Les algues sont obtenues généralement par la récolte directement de leur environnement de croissance ou en bord de plages, qui sont cependant de moins bonnes qualités que celles cultivées (Beaulieu et al., 2016). Cette faible qualité est principalement expliquée par le manque de
5
contrôle sur leur condition au moment de la récolte, notamment quant à leur âge et à l’uniformité des espèces disponibles (Beaulieu et al., 2016). C’est pour cette raison que 95,5% de la production mondiale des algues est faite en aquaculture afin d’assurer une production plus homogène (Beaulieu et al., 2016). Au Québec, les macroalgues sont encore obtenues par les récoltes sauvages, mais la tendance change puisque de nouvelles opportunités d’affaires s’ouvrent afin de répondre à la part de marché laissée vacante par l’Asie (Beaulieu et al., 2016). La méthode de culture sur filière (corde) développée en Asie se fait de plus en plus présente pour la production des algues laminaires (Lachance et al., 2013). De façon expérimentale, plusieurs algues rouges, comme P. palmata, peuvent être cultivées en bassin, à petite échelle dans les laboratoires ou dans de grands bassins (Lionard et al., 2014). L’algoculture se fait sur des fermes marines, qui sont des espaces délimités directement en mer (Lachance et al., 2013). Toutefois, la première étape de cette production se déroule dans des écloseries terrestres. Elle commence avec la fixation des plantules des macroalgues sur des cordes. À la suite d’une croissance de cinq semaines, qui s’effectue durant l’automne, les algues fixées aux cordes sont transportées aux fermes marines où elles sont mises à l’eau, fixées sur des filières subflottantes. Les cordes sont maintenues à des profondeurs contrôlées grâce à des flotteurs et sont attachées à des structures de béton au fond de l’eau à chacune de leur extrémité (Figure 1). Les filières sont initialement maintenues à une profondeur de 5-7 m, à la protection des intempéries de l’hiver. Au printemps, elles sont remontées à une profondeur de 3-4 m afin de permettre aux algues de maximiser leur croissance (Lachance et al., 2013). Finalement, les algues laminaires, comme S. longicruris, sont récoltées au début de l’été, lorsqu’elles ont atteint une longueur de 2 à 3 m, soit à un âge d’environ 8 à 10 mois (Lachance et al., 2013; Tamigneaux et al., 2013).
Pour les macroalgues de plus petites tailles, comme P. palmata, leur culture en bassin est possible. Les avantages de la production en bassin sont le contrôle du développement de la biomasse et la facilité de la récolte (Le Gall et al., 2004). En général, ce sont de jeunes macroalgues récoltées dans la nature qui sont mises
6
dans les bassins pour y être cultivées. La taille des bassins de culture peut varier selon les producteurs, par contre le principe reste essentiellement similaire. Les lieux de production sont généralement près des côtes puisque c’est l’eau de la mer qui y est directement pompée pour alimenter les bassins. Ainsi, grâce au système de pompage, l’eau des bassins est continuellement renouvelée. Selon les besoins, elle peut également être enrichie d’azote et d’autres éléments nutritifs essentiels pour la croissance des macroalgues (Beaulieu et al., 2016). De l’air est aussi injecté dans les bassins afin d’oxygéner l’eau et de simuler le mouvement des vagues dans la mer (Le Gall et al., 2004). Finalement, la luminosité peut être aussi contrôlée selon le lieu où les bassins sont installés, à l’extérieur ou à l’intérieur. L’exposition à la lumière est ajustée en fonction des besoins de croissance des macroalgues. Plusieurs études se sont d’ailleurs intéressées à l’amélioration du rendement de la production (Pang & Lüning, 2004). La Figure 2 présente deux macroalgues exploitées au Québec, S. longicruris et P. palmata.
Grâce à leur disponibilité au Québec et l’intérêt pour ces macroalgues, plusieurs entreprises productrices ont vu le jour en Gaspésie durant les dernières années. Les entreprises Varech Phare Est (2017) et Un Océan de Saveurs (2017) en sont des exemples.
Figure 1. Description de la filière subflottante utilisée pour la culture des laminaires
7
Figure 2. Saccharina longicruris (a) et Palmaria palmata (b) (Pêches et Océans
Canada, 2013; Tamigneaux, 2014).
2.1.1.2. Utilisation et consommation
Les macroalgues ont une utilité en technologie alimentaire. Leurs polysaccharides sont extraits pour être ajoutés dans des formulations d’aliments à titre d’agents texturants (Hernandez-Carmona, 2013). L’alginate provenant des algues brunes est utilisé comme agent humectant dans les aliments préparés et procure une texture tendre et uniforme aux aliments surgelés. Quant à l’agar, il remplace la gélatine dans les formulations traditionnelles des bonbons en gel. L’agar est également utilisé pour faire des desserts de gelée végétarienne. Le gel d’agarose est également utilisé dans le domaine de la biologie moléculaire pour la réalisation de gel électrophorétiques. Aussi, la carraghénane, par sa capacité stabilisante, peut être incorporée dans les jus de fruits ou le lait au chocolat pour mettre en suspension les ingrédients solides, comme les pulpes ou la poudre de cacao (Hernandez-Carmona, 2013).
En consommation humaine, les macroalgues sont déjà bien ancrées dans les habitudes alimentaires des populations côtières, et ce, même depuis la préhistoire (Dillehay et al., 2008). Actuellement, c’est surtout en Asie, principalement au Japon, en Corée du Sud, en Chine et aux Philippines, que les macroalgues sont incluses dans la culture gastronomique (Mouritsen, 2012). Les macroalgues peuvent être consommées crues en tant que salade ou incorporées dans les soupes. Ces légumes de mer libèrent, lors de la cuisson dans l’eau, un goût salé et d’umami,
8
compte tenu de leur quantité importante en minéraux et la présence d’acides glutamique et aspartique (Barbieri et al., 2016). Le Nori (Porphyra yezoensis) qui est bien connu de la population occidentale, est grillé et mis en feuille et sert abondamment dans la cuisine japonaise notamment dans les sushis. Au Canada, l’algue rouge P. palmata est une tradition bien établie des populations d’origine européenne de Nouvelle-Écosse et du Nouveau-Brunswick (Chopin & Ugarte, 2006). Elle est séchée et notamment servie comme apéritif dans les bars (Mahadevan, 2015). Plus récemment, la popularité des algues a même gagné le domaine des boissons alcoolisées. L’algue brune S. longicruris fait partie des ingrédients pour le Gin du St-Laurent (Distillerie du St-Laurent, 2017) et de la bière de la brasserie Marshall Wharf Brewing Co. (Spiegel, 2014).
2.1.2. Composition nutritionnelle
Comme tous les végétaux, la composition des algues est variable selon l’espèce et son environnement de croissance (Banerjee et al., 2009). Les algues sont reconnues pour leur valeur nutritionnelle élevée. Elles sont entre autres une excellente source de glucides, de protéines, de vitamines et de minéraux (MacArtain et al., 2007).
2.1.2.1. Protéines
Teneur en protéines des algues
Les algues rouges sont reconnues pour leur teneur élevée en protéines. Cette valeur peut atteindre 47% de la masse sèche alors que les algues brunes en contiennent entre 3 et 15% et entre 9 et 33% pour les algues vertes (Fleurence, 2004; Fujiwara-Arasaki et al., 1984; Young & Smith, 1958). Ces protéines sont composées d’une très grande diversité d’acides aminés essentiels dont la composition est très variable en fonction des espèces. Le profil en acides aminés essentiels de ces protéines est souvent comparable à d'autres protéines alimentaires telles que les légumineuses ou les œufs (Fleurence, 2004). Par exemple, pour une espèce d’algue brune comme Laminaria digitata, les acides glutamique et aspartique représentent 18% de la teneur totale en acides aminés (Augier & Santimone, 1978). En revanche, cette variabilité dans les proportions en acides aminés, selon les espèces, peut être
9
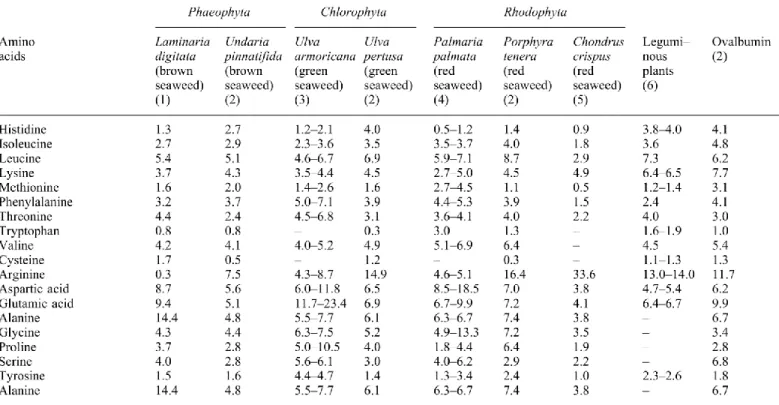
intéressante selon l’utilisation visée. Par exemple, l’algue P. palmata pourrait être utilisée comme un supplément en méthionine (2 mg/100 g d’algue sèche), selon une recommandation de la FAO (Fleurence, 1999). Le Tableau 1 présente la composition des acides aminés des différentes algues en comparaison avec d’autres aliments.
Facteur de conversion de la teneur en azote en teneur de protéines des algues Les méthodes de quantification de la teneur en protéines dans les aliments, comme celle de Kjeldahl, mesurent la teneur en azote total qu’ils contiennent. Cette quantité d’azote est convertie ensuite en quantité de protéines via un facteur de 6,25 (AOAC, 2002; Jones, 1931). Cependant, les algues contiennent une proportion d’azote qui n’est pas d’origine protéique. En effet, l’azote peut provenir des pigments, comme la chlorophylle et la phycoérythrine, et de composés inorganiques, comme le nitrite, le nitrate et l’ammoniac (Lourenço et al., 1998). Ainsi, l’utilisation du facteur de conversion traditionnel peut surestimer la valeur de protéines des algues. Même si la teneur en protéines peut varier par rapport à la teneur en azote d’une espèce d’algues à l’autre, des études ont été réalisées afin de déterminer le facteur de conversion adéquat. Le facteur de 4,92 a été suggéré par Lourenço et al. (2002) pour une estimation plus précise de la teneur totale en protéines des algues.
10
Tableau 1. Composition des acides aminés de différentes algues et d'autres
aliments (tiré intégralement de Fleurence, 2004).
(1) (Augier & Santimone, 1978); (2) (Fujiwara-Arasaki et al., 1984); (3) (Fleurence, 1999); (4) (Morgan et al., 1980); (5) (Young & Smith, 1958); (6) (Fowden, 1954).
En plus de leur potentiel nutritionnel, les acides glutamique et aspartique retrouvés dans L. digitata sont également source du goût de l’umami. Cette algue est, entre autres, très proche de L. japonica qui est à l’origine de la découverte du glutamate monosodique, un rehausseur de saveur très utilisé dans la cuisine asiatique (Marcus, 2005). D’autres acides aminés contribuent également au goût des macroalgues. L’alanine et la proline contribuent au goût sucré des macroalgues, alors que l’isoleucine, la leucine et la valine retrouvées dans le dashi de P. palmata sont responsables de l’amertume (Mouritsen et al., 2012).
2.1.2.2. Lipides
Les algues contiennent une faible teneur en lipides, soit jusqu’à 5% de la masse sèche (Nelson et al., 2002). La qualité des acides gras contenus dans les algues est non-négligeable puisque leur teneur en acides gras Oméga-3 et Oméga-6 représente jusqu’à la moitié de la totalité des acides gras. De plus, le ratio de ces
11
deux acides gras insaturés (ω6/ω3) est plus faible que 10 ce qui est idéal du point de vue nutritionnel (Mahan & Escott-Stump, 2000). Le plus haut ratio retrouvé dans les macroalgues était de 1,32 avec Himanthalia elongata, alors que P. palmata était à 0,13 (Sánchez-Machado et al., 2004). Peu importe la provenance de la macroalgue, cultivée ou sauvage, des chercheurs ont montré que P. palmata avait une abondance en acides myristique (C14:0), palmitique (C16:0) et eicosapentaénoïque (EPA) (C20:5 ω3) (Mishra et al., 1993).
2.1.2.3. Minéraux
Les algues marines retrouvées dans un milieu océanique contiennent une concentration importante en minéraux. La teneur totale en minéraux varie en fonction des espèces d’algues, mais globalement, elle peut atteindre jusqu’à 45% de son poids sec (Sánchez-Machado et al., 2004; Tibbetts et al., 2016; Wang et al., 2010). Puisque ces végétaux vivent dans l’eau salée, leur teneur en sodium est très élevée. Une algue rouge comme P. palmata en contient 255,2 mg/100 g sur base humide, alors qu’une algue brune comme L. digitata en contient beaucoup plus, 624,6 mg/100 g sur base humide. D’autres minéraux importants sont également présents en proportions importantes. Par exemple, pour une portion de 8 g sur base humide de P. palmata, la teneur en potassium et en calcium est de 74,4 mg et 584,8 mg, ce qui représente 10,6% et 16,7% de l’apport quotidien suggéré, respectivement (Committee on Medical Aspects of Food and Nutrition Policy, 1991). En ce qui concerne l’iode, celui-ci peut atteindre 3 650% de la valeur quotidienne suggérée (Institut de Phytonutrition, 2004; MacArtain et al., 2007).
2.1.2.4. Sucres
La quantité totale de sucres des macroalgues est, la plupart du temps, déterminée par différence, soit par la soustraction de la masse totale par la somme de la teneur en lipides, en protéines, en minéraux et en humidité (Tibbetts et al., 2016). La variation de la teneur en sucres peut être très grande selon la méthode utilisée pour mesurer les autres constituants, ce qui rend parfois difficile la comparaison entre les études (MacArtain et al., 2007).
12
Dans le cas des macroalgues, ce sont surtout les polysaccharides qui intéressent le domaine de la technologie alimentaire et de la nutrition. Les polysaccharides sont retrouvés en grande quantité dans la paroi cellulaire et ont un rôle très important pour la structure, la solidité et la flexibilité de la plante. Ces sucres sont aussi mis en réserve comme source d’énergie et régulent l’équilibre ionique dans la cellule (Gupta & Abu-Ghannam, 2011b).
Comme mentionné précédemment, l’alginate, la carraghénane et l’agar sont utilisés comme agents stabilisants et texturants. L’alginate provient des algues brunes, alors que la carraghénane et l’agar proviennent des algues rouges (Mabeau & Fleurence, 1993). D’autres polysaccharides sont également présents dans les algues. Parmi eux, on retrouve des xylanes, de la cellulose et des fucanes (Mabeau & Fleurence, 1993). La plupart de ces polysaccharides ne sont pas digestibles par l’humain, et sont donc considérés comme des fibres alimentaires (Southgate, 1990). Comparativement aux végétaux terrestres, les algues en contiennent autant ou même plus. Par exemple, pour une portion de 8 g de l’algue brune L. digitata ou de l’algue rouge P. palmata, la teneur en fibres totale est respectivement de 6,2 et de 5,4 g (Institut de Phytonutrition, 2004). Comparativement à du riz brun (3,8 g) et une banane (3,1 g), les algues en contiennent presque le double (Institut de Phytonutrition, 2004).
2.1.2.5. Vitamines
À la suite de stress imposés dans leur environnement, les macroalgues produisent des composés de protection, dont les vitamines, qui ont un rôle comme antioxydants. Les vitamines hydrosolubles (B et C) et liposolubles (A et E) sont présentes dans les algues (MacArtain et al., 2007). La vitamine E, qui est un puissant antioxydant, est présente en grande quantité dans l’algue P. palmata (1,296 mg/8 g sous base humide d’algues) alors que l’algue laminaire L. digitata n’en contient que seulement 0,275 mg pour une portion de 8 g (sous base humide) (Institut de Phytonutrition, 2004). De plus, contrairement aux plantes terrestres, les algues contiennent la vitamine B12 surtout connue pour son abondance dans les sources animales (Norziah & Ching, 2000). Watanabe et al. (1999) a démontré que
13
la vitamine B12 provenant des algues est biodisponible lors de la digestion. Pour combler 30% du besoin quotidien en vitamine B12, seulement une quantité de 3 g de P. palmata séchée suffisent (CEVA, 2009).
2.1.2.6. Polyphénols
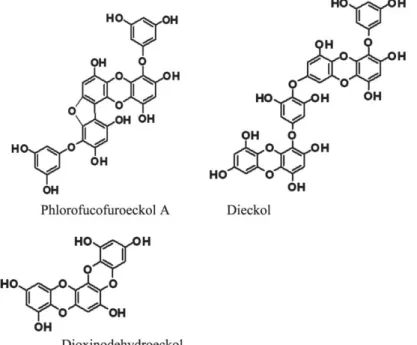
Les algues brunes contiennent des polyphénols suscitant beaucoup d’intérêt à cause de leur bioactivité. Reconnus comme antioxydants et ayant la capacité d’inhiber l’ECA, les phlorotannins, sous-groupe des tannins, sont des polymères de phloroglucinol (Kim et al., 2009). La Figure 3 présente les structures du phlorofucofuroeckol A, du dieckol et du dioxinodehydroeckol isolés d’Ecklonia stolonifera. La teneur en phlorotannins est généralement supérieure dans les algues brunes provenant de l’océan Atlantique et de la région tempérée de l’océan Pacifique comparativement à celles des régions tropicales (Targett & Arnold, 1998). Cette différence pourrait être expliquée par le rôle principal de ces polyphénols dans la construction des parois cellulaires et leur rôle secondaire en tant qu’agent de protection contre les radicaux libres qui sont formés lors d’un stress oxydatif environnemental (Arnold & Targett, 2003).
Figure 3. Structures de phlorofucofuroeckol A, dieckol et dioxinodehydroeckol
14
2.1.3.
Effets bénéfiques des macroalgues sur la santé
Un nombre croissant de recherches démontrent des effets bénéfiques potentiels sur la santé des composés provenant des macroalgues. Plusieurs auteurs ont fait des revues de littérature sur ce sujet (Beaulieu et al., 2016; Gupta & Abu-Ghannam, 2011a; Hamed et al., 2015; Kılınç et al., 2013; Yuan & Walsh, 2006). Les protéines et les peptides natifs ou résultants d’une hydrolyse des macroalgues ont démontré des potentiels bioactifs, par exemple, des capacités antioxydantes, antihypertensives in vitro et in vivo (Fitzgerald et al., 2012; Fitzgerald et al., 2014) et antibactériennes (Harnedy & Fitzgerald, 2011). D’autres effets bénéfiques potentiels des macroalgues incluent également les activités anti-cancers, anti-tumorales, antidiabétiques, antiallergiques et antivirales (El Gamal, 2010; Hamed et al., 2015; Harnedy & Fitzgerald, 2011; Tierney et al., 2010). Également, plusieurs études ont démontré la capacité antibactérienne et antifongique des algues (Beaulieu et al., 2015; Peres et al., 2012; Singh et al., 2015).
Dans ce présent projet, les capacités antioxydantes et inhibitrices de l’ECA (antihypertensives) in vitro sont celles qui ont été principalement étudiées.
2.1.3.1. Antioxydants
Les activités antioxydantes provenant des différents composés des macroalgues ont été étudiées à plusieurs reprises dans la littérature (Munir et al., 2013). Comme mentionné préalablement, leur environnement de croissance étant très changeant et parfois rude, les algues sont exposées à des stress qui provoquent l’apparition de radicaux libres à l’intérieur de leurs cellules (Yuan et al., 2009). Ceci explique pourquoi les algues ont développé un moyen de défense en produisant plusieurs composés antioxydants (Yuan et al., 2009). La quantité de certains de ces composés varie en fonction de la saison. En effet, les algues produisent moins de glutathions, d’ascorbates, de caroténoïdes et de tocophérols durant l’hiver et le printemps puisqu’elles sont moins exposées à la lumière à cette période (Aguilera et al., 2002; Burritt et al., 2002; Yuan, 2007). La présence des polysaccharides sulfatés, comme le fucoïdane, la laminarine et l’acide alginique, contribue également
15
à la capacité antioxydante de certaines algues (Rocha De Souza et al., 2007; Wijesekara et al., 2011).
De plus, les polyphénols contribuent grandement aux propriétés antioxydantes des algues. La quantité de polyphénols mesurée dans S. longicruris est de 0,41 ± 0,15% de sa biomasse sèche (Schiener et al., 2014), alors qu’une quantité de 12,8 µg d’équivalent à l’acide gallique a été mesurée dans des extraits méthanoliques de P. palmata (Yuan & Walsh, 2006). Les phlorotannins sont les plus étudiés et font partie de la famille des polyphénols retrouvés en grande quantité dans les algues brunes. Leur pouvoir antioxydant est plus élevé que certains autres antioxydants connus. Comme l’ont montré Kim et al. (2009), la concentration d’efficacité médiane du pouvoir antioxydant mesuré par la méthode DPPH de trois des phlorotannins isolés d'E. stolonifera est plus basse que l’acide ascorbique (10,3 ± 0,5 µM), soit le phlorofucofuroeckol A (4,7 ± 0,3 µM), le dieckol (6,2 ± 0,4 µM) et le dioxinodehydroeckol (8,8 ± 0,4 µM).
Les protéines et les peptides des macroalgues ont également été étudiés pour leur pouvoir antioxydant (Harnedy & Fitzgerald, 2011). L’activité est associée aux protéines de la famille des phycobiliprotéines, aux peptides carnosine et au glutathion (Fleurence, 2004; Sekar & Chandramohan, 2008; Shiu & Lee, 2005). D’autres études ont identifié la protéine RuBisCo comme précurseur des peptides bioactifs chez les macroalgues (Beaulieu et al., 2016; Bondu et al., 2014). Les méthodes d’extraction combinées avec une hydrolyse enzymatique, ou non, ont été réalisées afin de favoriser une extraction d'une plus grande concentration de protéines. Des extraits d’algue P. palmata hydrolysés ont démontré des valeurs ORAC entre 35,80 et 38,78 µmol TE/g, alors que S. longicruris a démontré une bioactivité plus élevée. Un extrait aqueux de poids moléculaire de > 10 kDa avait une valeur d’environ 250 µmol TE/g (Bondu et al., 2014).
2.1.3.2. Capacité inhibitrice de l’enzyme de conversion de l’angiotensine-I (ECA)
Les effets antihypertenseurs sont fréquemment mesurés indirectement en utilisant des tests in vitro. La capacité d’un composé ou d’un mélange de composés à inhiber
16
l’activité de l’enzyme de conversion de l’angiotensine-I en angiotensine-II (ECA) est alors mesurée (Jao et al., 2012). L’angiotensine-II est responsable de la vasoconstriction, ce qui fait augmenter la pression artérielle (Fitzgerald et al., 2004). Les deux principaux groupes de composés inhibiteurs de l’ECA étudiés chez les macroalgues sont les peptides ou protéines et les polyphénols.
En plus des effets antioxydants, les phlorotannins des algues brunes ont une très grande capacité d’inhiber l’ECA. La Figure 4 présente la double action des phlorotannins comme inhibiteurs de l’ECA et antioxydants. Afin de comparer l'efficacité des différents phlorotannins extraits d’Ecklonia cava, Wijesinghe et al. (2011) ont mesuré leur concentration minimale pour inhiber 50% de l’activité de l’ECA (IC50). Ils ont montré que le dieckol avait un IC50 de 1,47 ± 0,04 mM, alors que
les autres phlorotannins, comme le triphlorethol-A et l’eckstolonol, avaient des valeurs variant entre 2,01 et 2,95 mM. Le captopril, un composé contenu dans un médicament prescrit aux personnes atteintes de l’hypertension, a été utilisé comme contrôle et avait une valeur d’IC50 de 0,025 ± 0,90 µM (Wijesinghe et al., 2011).
Figure 4. Mécanismes potentiels des phlorotannins sur l'ECA et les radicaux libres
17
En plus de démontrer des effets antioxydants, des extraits d’algues enrichis en peptides et protéines ont été étudiés par plusieurs chercheurs pour la capacité d’inhiber l’ECA (Harnedy & Fitzgerald, 2011). Quant aux macroalgues québécoises, une étude a montré qu’un extrait de peptides de faible poids moléculaire (< 10 kDa) issus d’extraits de P. palmata hydrolysés avec de la chymotrypsine avait un IC50 de
460,05 mg/mL (Bondu et al., 2014). L’identification des protéines précurseures ayant une meilleure activité provenant des mêmes types d’extraits a permis de retrouver l’enzyme de RuBiSCo, des allophycocyanines et des phycocyanines. Ces derniers extraits à une concentration de 5 mg/mL avaient des activités inhibant entre 49,10 et 67,74% de l’ECA (Beaulieu et al., 2016). D’autres études ont mesuré directement la capacité antihypertensive de P. palmata en utilisant des essais in vivo sur un modèle animal du rat. Une hydrolyse des protéines de cette algue rouge avec l’enzyme papaïne a été effectuée et la purification du peptide actif a permis d’identifier la séquence en acides aminés du peptide suivant : IRLIIVLMPILMA. Cet extrait de protéines hydrolysées, ayant une valeur d’IC50 de 3,34 mM, a été donné à
des rats à une dose de 50 mg/kg de la masse de l’animal. Les résultats après 24 heures ont révélé une baisse de tension comparable au témoin traité avec du captopril (Fitzgerald et al., 2014a).
2.1.4.
Digestibilité
Malgré leur richesse nutritionnelle, la disponibilité des protéines de macroalgues n’est pas très élevée. En effet, plusieurs auteurs soulignent la faible digestibilité des protéines algales (Fujiwara-Arasaki et al., 1984; Galland-Irmouli et al., 1999; Mabeau & Fleurence, 1993). Des équipes de chercheurs ont évalué la digestibilité des protéines in vitro à partir d’extraits d’algues. Les extraits solubles de plusieurs macroalgues démontraient une digestibilité d’environ 50% avec la pancréatine à pH basique et de 70% avec la pronase par rapport à la protéine de référence, qui était la caséine (Fujiwara-Arasaki et al., 1984). La même tendance a été observée avec de la poudre de P. palmata, avec une valeur de 56% en utilisant la pepsine et la pancréatine porcine (Galland-Irmouli et al., 1999).
18
Plusieurs raisons peuvent expliquer la faible digestibilité des protéines des macroalgues. Certains composés contenus dans les macroalgues, tels les polyphénols, peuvent inhiber les enzymes responsables de la digestion (Chew & Rodman, 1979). De plus, les polysaccharides rendent inaccessibles les sites de coupure des protéines par les enzymes digestives. Acton et al. (1982) a démontré que le xylane présent dans les macroalgues et retrouvé principalement dans les algues rouges et vertes, peut réduire la digestibilité des protéines en les «entrapant» dans un réseau.
Une des solutions explorées afin d’augmenter la digestibilité des protéines algales est la fermentation. En utilisant des moisissures comme Aspergillus orysae, Rhizopus macroscopus et Trichoderma pseudokoningii pour effectuer la fermentation, la digestibilité des protéines a plus que doublée et peut atteindre 73% (Fleurence, 2004). La raison de cette amélioration est la capacité de ces moisissures à produire des glycosidases, comme l’endo- et l’exo-xylanase, la glucanase, la β-glucosidase et la cellobiohydrolase qui dégradent la paroi végétale cellulosique (Huang et al., 1983; Wey, et al., 1994).
2.1.5.
Produits
alimentaires
et
aliments
fonctionnels
additionnés de macroalgues
En raison de leur haute teneur en nutriments et de leur potentiel bioactif, les opportunités de production d’aliments fonctionnels à partir de macroalgues sont grandes. Plusieurs publications scientifiques ont présenté des travaux effectués sur des produits carnés dans l’objectif de réduire les maladies cardiovasculaires (Cardoso et al., 2015). Par exemple, une algue brune, L. japonica a été intégrée dans un pâté de poulet et de porc afin de favoriser la réduction du taux de glucose sanguin, de la proportion de lipoprotéines sanguines de basse densité, ainsi que de la teneur en cholestérol plasmatique (Hyeon & Hwa, 2014). Aussi, dans une formulation d’un jambon, la réduction en NaCl a été également possible en le remplaçant partiellement par un extrait de P. palmata. Une réduction pouvant atteindre jusqu’à 35% du sodium ajouté a été possible sans affecter la qualité organoleptique du produit (Barbieri et al., 2016). Un autre exemple concerne des
19
essais en laboratoire qui ont été réalisés pour produire un pain enrichi d’extrait de P. palmata. Le but de cette dernière étude était d’obtenir un pain ayant la capacité inhibitrice de l’ECA. Les résultats ont démontré que l’extrait de protéines hydrolysées de cette macroalgue n’avait pas affecté la qualité organoleptique du pain. De plus, même après la cuisson, l’activité inhibitrice de l’ECA était toujours présente avec l’ajout de 4% de cet extrait de protéines dans la formulation (Fitzgerald et al., 2014a).
2.2.
Le fromage
2.2.1.
Généralité
L’histoire des fromages québécois est beaucoup plus jeune que celle des fromages européens. L’influence de la culture anglaise a longtemps limité la production du fromage au Québec à celui de type Cheddar (Les Producteurs de lait du Québec, 2017). Le grand développement des variétés de fromages du Québec n’a commencé seulement qu’à la fin du XXe siècle. Aujourd’hui, plus de 450 variétés de
fromages différents sont reconnues et produites au Québec (Les Producteurs de lait du Québec, 2017). Ceci démontre la connaissance et le savoir-faire fromager grandissants au Québec.
2.2.2.
Différentes variétés de fromages
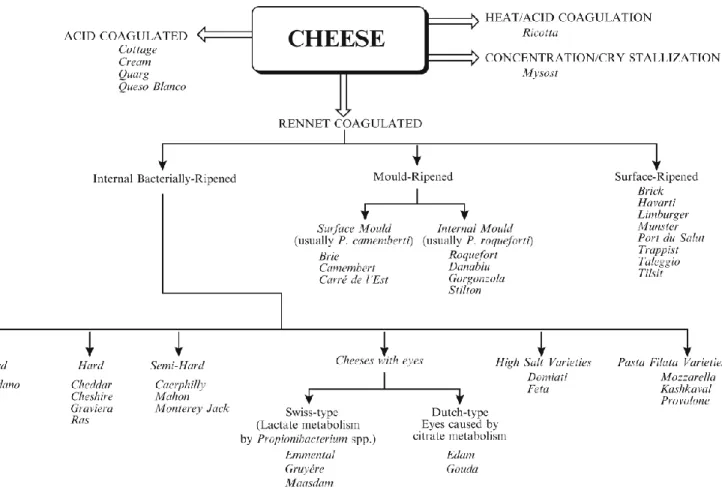
Les variétés de fromages peuvent être classées de plusieurs façons. Au-delà de l’origine biologique du lait (vache, chèvre, brebis, etc.), les fromages peuvent être divisés selon leur texture, soit comme pâte molle, semi-ferme, ferme ou très ferme (Fox & McSweeney, 2004). Une autre façon de classer les fromages est selon leur type d’affinage. Par exemple, un fromage de type Camembert est considéré comme un fromage à pâte molle à croute fleurie. Le diagramme suivant présente les principales variétés des fromages (Fig. 5).
20
Figure 5. Variétés des fromages classés selon leur méthode de coagulation et
d'affinage et leur texture (tirée intégralement de Fox et al., 2000).
2.2.3.
Fabrication du fromage
La fabrication fromagère passe par trois étapes importantes, soit la coagulation, l’égouttage et l’affinage (Johnson & Law, 2010). Le but ultime de la fabrication du fromage est la concentration du lait et ainsi d’augmenter sa durée de conservation (Johnson & Lucey, 2006).
La première étape de la production fromagère est la coagulation. Elle se traduit par la dénaturation des micelles de caséines du lait afin de former un gel, nommé le caillé (Fox et al., 2000). Cette étape commence par une acidification du lait par les ferments de bactéries lactiques inoculés. Des bactéries mésophiles sont ajoutées au lait pour transformer le lactose en acide lactique. L’acidification doit se faire jusqu’à pH 5,0-5,3 pour les fromages à pâte ferme et à pH 4,6 dans le cas des fromages à pâte molle, où un réseau de caséines se forme à ce moment (Johnson
21
& Law, 2010). Certains fromages se limitent seulement à cette étape de coagulation. D’autres nécessitent une autre réaction biochimique pour terminer la coagulation. Dans ce dernier cas, de la présure est ajoutée dans le lait acide pour favoriser davantage la fermeté du caillé. La présure commerciale est composée principalement de l’enzyme de la chymosine. C’est une coagulation dite enzymatique (Ruettimann & Ladisch, 1987).
La deuxième étape de la fabrication fromagère est l’égouttage qui consiste, en quelque sorte, à concentrer le lait. En effet, l’égouttage a pour but de retirer une grande partie de liquide du coagulum. Le caillé est coupé, brassé et mis en moule. Ainsi, le lactosérum est expulsé de la masse et seuls les protéines coagulées, principalement les caséines, le gras et les minéraux restent dans la matrice (St-Gelais & Tirard-Collet, 2002). Le salage du caillé vient ensuite et est très important afin de terminer l’égouttage. De différentes manières, en surface, dans la masse ou en saumure, ce procédé consiste à soutirer une quantité d’eau supplémentaire du caillé grâce à la force osmotique (Guinee, 2004).
Finalement, à l’étape de l’affinage, le fromage développe son goût et ses arômes. Des réactions biochimiques se produisent à l’intérieur du fromage lorsqu’il est entreposé dans des conditions adéquates. Des enzymes naturelles du lait, ou provenant des ferments bactériens et parfois fongiques, hydrolysent les protéines et les lipides pour libérer des composés aromatiques et volatils typiques de chaque fromage. Cette étape peut durer de quelques jours à plusieurs années (St-Gelais & Tirard-Collet, 2002).
Les fromages à pâte molle et à croute fleurie sont des fromages à affinage de courte durée, environ quelques semaines. Plusieurs activités biochimiques et microbiologiques s’y déroulent et sont dues à leur microflore d’affinage complexe. Cette complexité est, entre autres, la raison pour laquelle la structure de ces fromages peut être fragile et facilement perturbée (Spinnler & Gripon, 2004). La Figure 6 présente les principales étapes de production d’un fromage de type Camembert.
22
Figure 6. Protocole de production de fromage à pâte molle et à croute fleurie de
23
2.2.4.
Les caillés modèles
Le fromage étant une matrice complexe et contenant une très grande diversité de composés, il est difficile de produire un milieu de culture lui ressemblant. De plus, produire des fromages selon les traitements désirés peut rapidement devenir incontrôlable et couteux (Rehman et al., 2001). Ce sont pour ces raisons que depuis plusieurs années, des chercheurs se sont tournés vers les caillés modèles pour l’étude du comportement du fromage (Lessard et al., 2012; Singh & Kristoffersen, 1971). Le caillé modèle est une imitation très proche du fromage régulier, puisqu’il est fait à partir du caillé fromager qui a été stabilisé. Ainsi, cette matrice est composée de tous les éléments présents dans le fromage et offre la possibilité de contrôler la composition de celui-ci (Rehman et al., 2001). Grâce au caillé modèle, le développement de la microflore d’affinage, le changement de la structure du fromage et l’apparition des composés volatils peuvent être plus facilement étudiés (Boutrou et al., 2002; Di Cagno et al., 2006; Jollivet et al., 1993).
2.2.5.
Défauts de contamination du fromage
La contamination des produits fromagers par des pathogènes (Salmonella spp., Escherichia coli O157:H7, Listeria monocytogenes et Staphylococcus aureus) est une préoccupation très importante des producteurs. Des outils de contrôle, comme un système de contrôle de la qualité (HACCP) et des règles d’hygiène tout au long de la production permettent de minimiser les risques (Donnelly, 2004). Par contre, d’autres problèmes de contamination, provenant de la microflore d’altération, peuvent aussi survenir et provoquer des défauts de qualité et d’esthétique du produit fini.
Dans certains cas, le résultat de la contamination se solde en des défauts de croute ou de présentation. Souvent, ces défauts proviennent de la microflore normale du fromage. C’est un développement non désiré ou trop important d’un ferment provenant du fromage lui-même ou de la microflore de l’environnement du lieu de production (Eck & Gillis, 2006). Parmi ces problèmes, très fréquents chez les fromages à pâte molle et à croute fleurie, les accidents du «bleu» mènent à des tâches bleu-verdâtre sur la surface du fromage. Une des causes est la présence de
24
Penicillium expansum, une moisissure qui se développe mieux dans un milieu plus acide que Penicillium camemberti (Guéguen, 1988). Il existe également des accidents nommés «poil de chat», qui se présentent sous forme de duvet blanchâtre qui peut devenir grisâtre à noirâtre. Ce défaut est dû à des moisissures dont, entre autres, Mucor racemorus provenant de l’environnement qui contaminent la surface des fromages (Devoyod, 1988; Guéguen, 1992). Les fromages à pâte molle sont aussi assujettis au défaut de la «graisse» qui survient quand Geotrichum candidum se développe abondamment. La surface du fromage devient alors plissée et graisseuse, ce qui empêche le développement de P. camemberti (Guéguen, 1988, 1992).
2.2.6.
Rôles des constituants du fromage lors de l’affinage
Afin d’éviter tout problème relié aux défauts qui peuvent survenir lors de l’affinage, les rôles des principaux agents d’affinage doivent être bien compris, notamment dans le cas des fromages ayant une microflore complexe comme celle du type Camembert.2.2.6.1. Sel
Le sel est nécessaire dans le fromage et y occupe plusieurs fonctions (Fig. 7). La première est sa contribution au goût. Le sel a la capacité de masquer et de rehausser certaines saveurs développées dans le fromage durant son affinage (Hardy, 2006). La deuxième concerne son rôle dans l’expulsion de l’eau contenue dans la matrice lors de l’égouttage final du fromage. De plus, dans certains cas, il aide à former la croute du fromage. Également, le sel contrôle les réactions biochimiques (enzymatiques) et microbiologiques (contrôle des ferments, inhibition des microflores délétères). Ainsi, le fromage peut être mieux conservé (Hardy, 2006). La présence de sel dans un aliment comme le fromage abaisse son activité de l’eau (Aw). Les réactions biochimiques qui participent au développement de saveurs et d’arômes du fromage ont besoin d’une Aw minimale. Par exemple, les réactions enzymatiques comme celles catalysées par les peroxydases sont inactives à Aw 0,85, alors que les lipases peuvent aller jusqu’à 0,1 (Ludwig, 1963). Cette
25
influence du sel sur les activités enzymatiques contribue aussi au développement de la texture du fromage (Fox et al., 2000). Également, en réduisant l’activité de l’eau, cela permet de contrôler le développement de l’activité des ferments. Ainsi, l’Aw du fromage diminue durant l’affinage, car le sel contribue à retirer l’eau de la matrice fromagère (Marcos & Esteban, 1982). Ainsi, ce changement d’Aw explique en partie la succession de la microflore d’affinage du fromage. Des bactéries moins halophiles peuvent se développer initialement. Ensuite, elles laisseront leur place à d’autres microorganismes plus résistants aux basses valeurs d’Aw (Hardy, 2006). Par contre, l’Aw seule ne peut pas contrôler le développement des microorganismes, car certains non désirables peuvent tout de même croître. En effet, les valeurs de l’Aw des fromages sont de 0,985 pour le Camembert, 0,950 pour le Cheddar et de 0,917 pour le parmesan (Rüegg & Blanc, 1981). Certaines moisissures, désirables ou non, sont capables de se développer à partir d’une Aw de 0,70 (Hardy, 2006). La Figure 7 résume les rôles du sel dans le fromage.
Figure 7. Contribution du sel pour la conservation, la sécurité et la qualité générale
du fromage (tirée intégralement de Guinee, 2004).
2.2.6.2. Ferments d’affinage
Plusieurs facteurs influencent le fromage durant son affinage. L’un des plus importants est sans doute la présence des microorganismes, soit la microflore d’affinage (Fox et al., 2000). Pour un fromage à pâte molle et à croute fleurie comme le Camembert, le rôle des ferments est primordial. Le phénomène de succession des microflores est déterminé par la composition chimique de la matrice fromagère.
26
Initialement, les bactéries lactiques sont introduites dans le lait lors de la fabrication, afin de réduire le pH. Ainsi, le caillé est un milieu sélectif pour la croissance microbienne. Seuls les organismes acidophiles, comme les levures et moisissures, ont la capacité de croître (Spinnler & Gripon, 2004).
Des levures, comme G. candidum, capables de résister à des pH acides (< 5), apparaissent durant les premières journées de l’affinage (Leclercq-Perlat et al., 2004). En métabolisant le lactate et en produisant de l’ammoniac, la croissance de G. candidum engendre l’élévation du pH de la surface du caillé, ce qui permet à des moisissures plus acidosensibles de se développer (Boutrou & Guéguen, 2005). Après 6 à 7 jours d’affinage, le mycélium de la moisissure P. camemberti est visible à la surface du fromage. Cette croissance est favorisée par l’élévation du pH, accentuée par G. candidum. Après 2 à 3 jours, la surface du fromage est complètement recouverte du duvet blanc de la moisissure. Ce recouvrement est voulu, car sa présence protège le fromage des autres microorganismes non désirables par encombrement de l’espace. Sa croissance accélère le changement du pH de la surface en consommant également le lactate disponible (Spinnler & Gripon, 2004). Ainsi, le pH au centre du caillé est plus acide qu’à la surface créant alors un gradient. Tout au long de l’affinage, l’élévation du pH progresse graduellement vers le centre. La Figure 8 présente les directions que prennent les différents composés responsables du changement de pH du caillé.
27
Figure 8. Gradient des composés responsables de l’alcalinisation du caillé du
fromage Camembert (tirée intégralement de Spinnler & Gripon, 2004).
De plus, l’évolution du pH du caillé se rapprochant de la neutralité favorise l’activité des différentes enzymes présentes. Ces enzymes sont déjà présentes dans le lait (plasmine), ajoutées lors de la fabrication (présure) ou produites par les ferments lactiques ou lors de l’affinage (Boutrou & Guéguen, 2005). Elles hydrolysent des protéines et des lipides du caillé, conduisant ainsi à l’apparition des composés aromatiques et volatils responsables de la saveur et du changement de la texture typique du fromage (Boutrou & Guéguen, 2005).
Protéases
Initialement, l’activité de la présure est très dominante, puisqu’elle est plus active à un pH acide. Le caillé du fromage de type Camembert contient une plus grande quantité de présure à l’intérieur de sa masse (Vassal & Gripon, 1984). L’apparition de la caséine-αs1-I est détectable partout à travers le caillé, ce qui est le résultat de
la dégradation de la caséine-αs1 par la présure. Cette activité est toutefois ralentie
et inhibée à la suite de l’augmentation du pH du milieu (Vassal & Gripon, 1984). Ensuite, à un pH plus près de la neutralité, ce sont les exo- et endoprotéases des levures et moisissures, combinées à celles des bactéries lactiques, qui sont activent.
28
L’hydrolyse des caséines α et β continue à se réaliser et à engendrer l’apparition des peptides et des acides aminés qui à leur tour sont transformés en composés aromatiques (Adda et al., 1982). Une activité trop importante des protéases peut toutefois conduire à la génération d’amertume dans le fromage (Vassal & Gripon, 1984).
Lipases
Les levures et les moisissures qui composent la microflore secondaire du Camembert sont capables de produire une large gamme de lipases. Ces enzymes dégradent les triglycérides du caillé pour libérer des di- et des monoglycérides ainsi que des acides gras libres (Spinnler & Gripon, 2004). La transformation de ces composés lipidiques est très importante pour la qualité organoleptique du fromage. En ce qui concerne la texture, la présence d’une grande quantité de monoglycérides permet de donner au fromage une tendreté en bouche. Ceux-ci agissent comme agents émulsifiants, réduisant la taille des globules de gras de la matrice du fromage (Miettinen et al., 2002). Par contre, cette texture n’est pas uniforme dans l’ensemble du fromage. Les lipases étant sensibles aux pH acides, elles commencent à agir seulement en dessous de la croute du fromage et progressivement vers le centre. C’est pour cette raison qu’un fromage Camembert jeune a une texture coulante seulement à la périphérie située sous la croute (Spinnler & Gripon, 2004). Finalement, les acides gras sont des précurseurs des composés aromatiques du fromage. Ce sont les moyens et petits acides gras (4-12 carbones) qui offrent d’ailleurs les meilleurs seuils de perception olfactifs (Molimard & Spinnler, 1996).
2.2.7.
Bioactivités
De nombreuses études ont démontré les bioactivités in vitro des produits laitiers, notamment celles provenant des peptides. Dans le cas des produits fermentés comme le fromage, des effets immunomodulateurs, anticancéreux, hypocholestérolémiants, antimicrobiens, antihypertenseurs et antioxydants ont été mis en évidence (Choi et al., 2012; Fitzgerald & Murray, 2006; Hernández-Ledesma et al., 2005). Le Tableau 2 présente les différentes bioactivités des peptides et leurs protéines précurseures.
29
Tableau 2. Différentes bioactivités des produits laitiers fermentés et leurs protéines précurseures (tirée intégralement de Choi et al., 2012).
(Gobbetti et al., 2002; Hayes et al., 2007; Meisel & Bockelmann, 1999; Möller et al., 2008; Smacchi & Gobbetti, 2000; Tidona et al., 2009)
2.2.7.1. Antioxydants
Plusieurs auteurs mentionnent que les peptides antioxydants proviennent principalement de l’activité protéolytique de plusieurs bactéries lactiques ajoutées ou indigènes, telles que Lactococcus lactis, Lactobacillus helveticus et Lactobacillus delbrueckii ssp. bulgaricus (Korhonen & Pihlanto, 2006). Les peptides antioxydants ont surtout été retrouvés dans les extraits solubles (Kuchroo & Fox, 1982). Selon le type de fromage, la capacité antioxydante peut varier. L’analyse du fromage Pecorino a montré une valeur ORAC stable de 20,40 µmol TE/g durant les 60 jours de son affinage (Branciari et al., 2015). Une autre étude a montré que la capacité antioxydante du fromage Cheddar augmentait durant les quatre premiers