Évaluation de méthodes de désherbage dans le
maïs-grain (Zea mays) avec culture intercalaire de ray-grass
annuel (Lolium multiflorum) et de trèfle incarnat
(Trifolium incarnatum).
Mémoire
Philippe Roch
Maîtrise en biologie végétale - avec mémoire
Maître ès sciences (M. Sc.)
Évaluation de méthodes de désherbage dans le
maïs-grain (Zea mays) avec culture intercalaire de
ray-grass annuel (Lolium multiflorum) et de trèfle
incarnat (Trifolium incarnatum)
Mémoire
Philippe Roch
Sous la direction de :
Résumé
Une culture intercalaire composée de ray-grass annuel et de trèfle incarnat dans le maïs-grain peut apporter de nombreux bénéfices environnementaux. Cependant, l’utilisation de ce mélange de cultures intercalaires est peu documentée avec les herbicides résiduels, le désherbage mécanique et intégré, ce qui rend la pratique très dépendante de l’usage du glyphosate. Des essais au champ ont été mis en place en 2017 et 2018 dans le maïs-grain, intercalé de ray-grass et de trèfle incarnat pour évaluer différentes méthodes de désherbage. Le volet A évalue l’utilisation de différents herbicides résiduels aux doses recommandées sur l’étiquette des produits pour l’établissement du mélange d’intercalaires. L’isoxaflutole + atrazine procure un désherbage et un établissement adéquats de l’intercalaire sur un sol sableux à 2 % d’argile, mais de la chlorose sur le trèfle apparaît sur un sol à 7 % d’argile. La mésotrione en postlevée du maïs avec le glyphosate semble causer peu de dommages aux intercalaires, tout en procurant un bon désherbage. Le volet B évalue différentes combinaisons d’isoxaflutole + atrazine à demi-dose, de désherbage mécanique avec une herse-peigne, de tembotrione/thiencarbazone et de glyphosate. L’isoxaflutole + atrazine cause des dommages au trèfle sur un loam, mais pas sur un loam sableux. La herse-peigne utilisée en prélevée et postlevée procure un désherbage de bonne qualité et un établissement d’intercalaires non négligeable sur un loam sableux, mais n’est pas adaptée au loam.
Abstract
Intercropping annual ryegrass and crimson clover in grain-corn can bring many environmental upsides. However, the use of this intercropped mix with residual herbicides, mechanical and integrated weeding is sparsely documented. Thus the practice is mainly dependant on the use of glyphosate for weeding. In-field trials were made in 2017 and 2018 in grain corn, intercropped with ryegrass and incarnate clover to assess different weeding techniques. The part A of this experience assessed the use of different residual herbicides at label dose to establish this intercropped mix. Isoxaflutole + atrazine provides a good weeding and intercrop establishment in a 2 % clay soil, but causes chlorosis to incarnate clover in a 7 % clay soil. Mesotrione + glyphosate at corn preemergence seems to provide a good intercrop’s establishment and weeding quality. Part B assess different combinations of half-dosed isoxaflutole + atrazine, mechanical weeding with spring-tine weeder, tembotrione/thiencarbazone and glyphosate. Isoxaflutole + atrazine causes chlorosis to incarnate clover on a loam soil but not on a sandy loam. Spring-tine weeder can provide a good weeding and a fair establishment of intercrop on a sandy loam but is unable to do so on a loam.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... viii
Liste des tableaux ... ix
Liste des abréviations ... xi
Remerciements ... xiii
Introduction ... 1
Chapitre 1 Revue de littérature ... 2
1.1 Les mécanismes de la compétition entre les plantes ... 2
1.1.1 Importance de la biomasse dans le contrôle des mauvaises herbes ... 2
1.1.2 Synergie entre les légumineuses et les graminées ... 2
1.1.3 Utilisation de l’azote dans un contexte de compétition entre végétaux ... 3
1.1.4 Allélopathie ... 4
1.2 Les cultures intercalaires et l’environnement ... 5
1.2.1 Réduction des pertes de sol ... 5
1.2.2 Réduction de la diffusion de contaminants agricoles ... 5
1.2.3 Réduction de la fertilisation azotée ... 5
1.3 Le ray-grass ... 6
1.3.1 Caractéristiques du ray-grass... 6
1.3.2 Culture du ray-grass ... 7
1.4 Trèfle incarnat ... 8
1.4.1 Caractéristiques du trèfle incarnat ... 8
1.4.2 Culture du trèfle incarnat ... 10
1.5 Implantation d’un intercalaire ... 11
1.6 Désherbage mécanique en intercalaire ... 12
1.6.1 Outils de désherbage mécanique ... 12
1.6.2 Désherbage mécanique avec intercalaire aux champs ... 13
1.7 Désherbage chimique en prélevée ... 16
1.7.1 Atrazine ... 16
1.7.3 Isoxaflutole ... 17
1.7.4 La tolérance des intercalaires aux herbicides de prélevée ... 18
1.8 Désherbage chimique en postlevée ... 20
1.8.1 Le Glyphosate ... 20
1.8.2 La Tembotrione ... 21
1.8.3 Le thiencarbazone-méthyl... 22
1.8.4 La tolérance des intercalaires aux herbicides de postlevée ... 22
1.9 Hypothèses et objectifs ... 23
Chapitre 2 Matériel et méthodes... 24
2.1 Conditions météorologiques ... 24
2.2 Localisation et caractéristiques des sites expérimentaux ... 25
2.3 Mise en place des parcelles expérimentales ... 26
2.4 Variables à l’étude ... 34
2.5 Dispositif expérimental ... 37
2.6 L’analyse statistique ... 38
Chapitre 3 Résultats volet A ... 40
3.1 Recouvrement des cultures intercalaires ... 40
3.1.1 Recouvrement du trèfle incarnat ... 40
3.1.2 Recouvrement du ray-grass ... 43
3.1.3 Recouvrement des deux cultures intercalaires ... 46
3.2 Biomasse sèche des cultures intercalaires ... 49
3.2.1 Biomasse sèche du trèfle incarnat ... 49
3.2.2 Biomasse sèche du ray-grass ... 51
3.2.3 Biomasse sèche des deux cultures intercalaires ... 53
3.3 Chlorose sur les cultures intercalaires ... 55
3.3.1 Chlorose sur le trèfle incarnat ... 55
3.4 Gestion des mauvaises herbes ... 58
3.4.1 Flore des mauvaises herbes aux sites expérimentaux ... 58
3.4.2 Recouvrement des mauvaises herbes ... 58
3.4.3 Biomasse sèche des mauvaises herbes ... 62
3.5 Discussion du volet A ... 64
Conclusion volet A ... 66
4.1Recouvrement des cultures intercalaires ... 67
4.1.1 Recouvrement du trèfle incarnat ... 67
4.1.2 Recouvrement du ray-grass ... 71
4.1.3 Recouvrement total des cultures intercalaires ... 75
4.2 Biomasse sèche des cultures intercalaires ... 79
4.2.1 Biomasse sèche du trèfle incarnat ... 79
4.2.2 Biomasse sèche du ray-grass ... 82
4.2.3 Biomasse sèche totale des cultures intercalaires ... 85
4.3 Gestion des mauvaises herbes ... 88
4.3.1 Liste des mauvaises herbes ... 88
4.3.2 Recouvrement des mauvaises herbes ... 88
4.3.3 Biomasse sèche des mauvaises herbes ... 91
4.4 Rendement du maïs-grain ... 94
4.4.1 Rendement en maïs-grain ... 94
4.5 Discussion ... 96
Conclusion volet B ... 98
Bibliographie ... 100
Annexe 1 : Illustration d’une année-site pour le volet A. ... 104
Annexe 2 : Illustration d’une année-site pour le volet B. ... 105
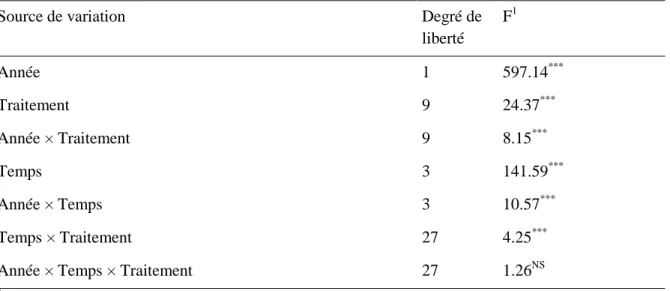
Annexe 3 Tableau d'analyse de la variance d’un bloc complet aléatoire avec mesures répétées sur deux années du volet A. ... 106
Annexe 4 Tableau d'analyse de la variance en split-plot du volet A. ... 107
Annexe 5 Liste des contrastes du volet A ... 108
Annexe 6 Tableau d'ANOVA split-split plot avec mesure répétées 4 fois du volet B ... 109
Annexe 7 Tableau d’analyse de variance en split-plot avec mesures répétées 4 fois sur un lieu du volet B ... 111
Annexe 8 Tableau d'analyse de variance en split-plot avec mesures répétées deux fois du volet B. ... 112
Annexe 9 Tableau d'analyse de la variance en split-split-plot du volet B ... 113
Annexe 10 Tableau d'analyse de variance en split-plot du volet B. ... 114
Annexe 11 Liste des contrastes du volet B ... 115
Annexe 12 : Photo d’une parcelle désherbée avec du glyphosate sur le loam à la fin d’octobre 2018. ... 116
Annexe 13 : Photo d’une parcelle désherbée avec du glyphosate sur le loam sableux à la fin
d’octobre 2018. ... 117 Annexe 14 : Photo d’une parcelle désherbée avec la herse-peigne en prélevée + postlevée sur le loam sableux à la fin d’octobre 2018. ... 118 Annexe 15 : Contrastes a priori pour le recouvrement du trèfle incarnat (%) et valeur de Fisher (F) selon l'interaction temps × traitement au volet A. ... 119 Annexe 16 : Contrastes a priori pour le recouvrement du ray-grass (%) et valeur de Fisher (F) selon l'interaction temps × traitement au volet A. ... 120 Annexe 17 : Contrastes a priori pour le recouvrement des deux cultures intercalaires (%) et valeur de Fisher (F) selon l'interaction temps × traitement au volet A. ... 121 Annexe 18 : Rendement (Mg ha-1) du maïs grain au volet A par traitement. ... 122 Annexe 19 : Contrastes a priori sur le recouvrement total de mauvaises herbes (%) et valeur de Fisher (F) pour le rang selon l’interaction traitement × site × lieu pour le volet B... 123 Annexe 20 : Contrastes a priori sur le recouvrement total de mauvaises herbes (%) et valeur de Fisher (F) selon l’interaction temps × lieu × traitement pour le volet B. ... 124
Liste des figures
Figure 1 : Biomasse de mauvaises herbes suite à un désherbage mécanique dans une culture de maïs intercalé d'une légumineuse ou non.Adaptée de Bialis et coll. (2010) ... 15 Figure 2 Réduction ou augmentation de la biomasse par différents herbicides de prélevée sur le ray-grass annuel et le trèfle rouge. ... 19 Figure 3 Précipitations totales entre mai et août à la station agronomique de Saint-Augustin-de-Desmaures en 2017 et 2018. ... 24 Figure 4 Précipitations mensuelles à la station agronomique de Saint-Augustin-de-Desmaures en 2017 et 2018. ... 25 Figure 5 Schéma d'une parcelle (Minville, 2017). ... 38
Liste des tableaux
Tableau 1 Vitesse d'atteinte de la croissance maximale et biomasse maximale de diverses espèces de
trèfles. ... 9
Tableau 2 Biomasse sèche de mauvaises herbes et rendement en grains pour une culture de maïs-grain intercalé et désherbé mécaniquement. ... 14
Tableau 3 Coordonnées géographiques des sites expérimentaux en 2017 et 2018. ... 26
Tableau 4 Caractéristiques des sols des sites expérimentaux en 2017 et 2018. ... 26
Tableau 5 Informations sur le semis de maïs. ... 27
Tableau 6 Traitements herbicides du volet A. ... 28
Tableau 7 Information sur les traitements herbicides du volet A. ... 29
Tableau 8 Traitements de désherbage du volet B. ... 30
Tableau 9 Noms commerciaux des herbicides utilisés. ... 31
Tableau 10 Information sur les traitements herbicides sur toute la surface du volet B. ... 31
Tableau 11 Information sur les traitements de herse-peigne du volet B. ... 32
Tableau 12 Information sur les traitements herbicides en bande du volet B. ... 33
Tableau 13 Informations sur le semis des cultures intercalaires. ... 34
Tableau 14 Dates de prises de donnée du volet A. ... 36
Tableau 15 Date de prises de données du volet B. ... 37
Tableau 16 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement du trèfle incarnat. ... 40
Tableau 17 Contrastes a priori pour le recouvrement du trèfle incarnat (%) et valeur de Fisher (F) selon l'interaction traitement × année. ... 42
Tableau 18 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement du ray-grass. .... 43
Tableau 19 : Contrastes a priori pour le recouvrement du ray-grass (%) et valeur de Fisher (F) selon l’interaction traitement × année. ... 45
Tableau 20 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement total des cultures intercalaires (%). ... 46
Tableau 21 Contrastes a priori pour le recouvrement total des cultures intercalaires (%) et valeur de Fisher (F) selon l'interaction traitement × année. ... 48
Tableau 22 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche du trèfle incarnat. ... 49
Tableau 23 Contrastes a priori pour la biomasse sèche du trèfle incarnat (g m-2) et valeur de Fisher (F) selon l'interaction traitement × année. ... 50
Tableau 24 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche du ray-grass. . 51 Tableau 25 Contrastes a priori pour la biomasse sèche du ray-grass (g m-2) et valeurs de Fisher (F). ... 52
Tableau 26 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche totale des cultures intercalaires. ... 53
Tableau 27 Contrastes a priori pour la biomasse sèche totale des cultures intercalaires (g m-2) et valeur de Fisher (F). ... 54 Tableau 28 Valeur de Fisher (F) de l’analyse de la variance pour la chlorose sur le trèfle incarnat. 55
Tableau 29 Contrastes a priori pour le pourcentage de chlorose sur le trèfle incarnat (%) et valeur de Fisher (F) selon l'interaction traitement × année... 57 Tableau 30 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement des mauvaises herbes annuelles. ... 58 Tableau 31 Contrastes a priori portant sur le recouvrement des mauvaises herbes (%) et valeur de Fisher selon l'interaction traitement × temps × année. ... 60 Tableau 32 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche de mauvaises herbes. ... 62 Tableau 33 Contrastes a priori sur la biomasse sèche de mauvaises herbes et valeur de Fisher (F) selon l’interaction traitement × année (g m-2). ... 63
Tableau 34 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement du trèfle incarnat. ... 67 Tableau 35 Contrastes a priori sur le recouvrement de trèfle incarnat (%) et valeur de Fisher (F) selon l’interaction site × temps × traitement. ... 69 Tableau 36 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement ray-grass. ... 71 Tableau 37 Contrastes a priori sur le recouvrement du ray-grass (%) et valeur de Fisher (F) selon l’interaction site × temps × traitement. ... 73 Tableau 38 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement total des cultures intercalaires. ... 75 Tableau 39 Contrastes a priori sur le recouvrement total des cultures intercalaires (%) et valeur de Fisher (F) selon l’interaction site × temps × traitement. ... 77 Tableau 40 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche du trèfle
incarnat. ... 79 Tableau 41 Contrastes a priori sur la biomasse sèche du trèfle incarnat (g m-2) et valeur de Fisher (F) selon l’interaction traitement × site. ... 81 Tableau 42 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche du ray-grass .. 82 Tableau43 Contrastes a priori sur la biomasse sèche du ray-grass (g m-2) et valeur de Fisher (F) selon l’interaction traitement × temps. ... 84 Tableau 44 Valeur de Fisher de l’analyse de la variance pour la biomasse sèche totale des cultures intercalaires. ... 85 Tableau 45 Contrastes a priori sur la biomasse sèche totale des cultures intercalaires (g m-2) et valeur de Fisher (F). ... 87 Tableau 46 Valeur de Fisher (F) de l’analyse de la variance pour le recouvrement des mauvaises herbes. ... 88 Tableau 47 Contrastes a priori sur le recouvrement total de mauvaises herbes (%) et valeur de Fisher (F) pour l’entre-rang selon l’interaction traitement × site × lieu. ... 90 Tableau 48 Valeur de Fisher (F) de l’analyse de la variance pour la biomasse sèche des mauvaises herbes. ... 91 Tableau 49 Contrastes a priori sur la biomasse sèche totale de mauvaises herbes (g m-2) et valeur de Fisher (F) selon l’interaction traitement × site. ... 93 Tableau 50 Valeur de Fisher (F) de l’analyse de la variance pour le rendement du maïs-grain. ... 94 Tableau 51 Contrastes a priori portant sur le rendement du maïs-grain (Mg ha-1) et valeur de Fisher (F). ... 95
Liste des abréviations
C : Biomasse maximale CI : Cultures intercalaires C1 : Contraste 1 C2 : Contraste 2 C3 : Contraste 3 C4 : Contraste 4 C5 : Contraste 5 C6 : Contraste 6 C7 : Contraste 7 C8 : Contraste 8 C9 : Contraste 9 CI : Cultures intercalaires LER : Land equivalent ratioM : Atteinte de la croissance maximale M.O. : Matière organique
NLER : Nitrogen land equivalent ratio PRÉ : Prélevée du maïs
POST : Postlevée du maïs
PRÉ-B1 : Traitement en prélevée en bande avant le passage de la herse-peigne prélevée PRÉ-B2 : Traitement en prélevée en bande après le passage de la herse-peigne en prélevée PRÉ 1 : Traitement en prélevée sur toute la surface
PRÉ 2 : Traitement en prélevée sur toute la surface après un passage de herse-peigne en prélevée POST : Traitement en postlevée sur toute la surface
Remerciements
Je tiens à exprimer ma reconnaissance à tous ceux qui ont contribué de quelques façons que ce soit à la réalisation de ce mémoire. D’abord, je tiens à remercier sincèrement mon directeur, le Dr. Gilles D. Leroux, professeur au département de Phytologie de l’Université Laval, pour son support, son esprit critique, sa rigueur et sa grande disponibilité qui ont permis l’accomplissement de ce projet.
Je tiens aussi à remercier M. David Miville, professionnel de recherche au Département de Phytologie de l’Université Laval pour son implication, son encadrement technique et sa disponibilité dans la réalisation des travaux aux champs. Un merci particulier aussi à M. Sébastien Rouane, professionnel de recherche au Département de Phytologie de l’Université Laval pour son aide inestimable au traitement et à l’analyse des données. Je remercie également M. Gaétan Daigle, consultant en statistique à l’Université Laval pour son aide précieuse avec le logiciel SAS et les statistiques.
Merci aussi à M. Francis Gagnon et M. Pascal Tessier pour leur assistance technique sur le terrain et aux étudiants de premier cycle trop souvent oubliés qui ont participé aux travaux au champ à la sueur de leur front : Audrey-Kim Minville, Christine Lebeau, Pierre-Alexandre Labranche, Roxanne Binet-Robert, Jean-François Patenaude, Marc-Antoine Chiasson, Joëlle Landry, Alexis Gloutney, Audrey Savard, Simon Goyette et Justine Clément.
Merci du fond du cœur à mes parents qui m’ont toujours soutenu et encouragé tout au long de ce projet et de ma vie en général.
Merci aussi à la famille et aux amis d’avoir été là tout au long de ces deux années.
Un dernier remerciement mais non le moindre à ma conjointe Valérie. Pour ton encouragement, ta patience, ton soutien, tes sacrifices qui m’ont permis de persévérer dans ce projet. Nous pouvons maintenant avancer ensemble dans la vie et réaliser les projets qui nous tiennent à cœur.
Introduction
Face à la multiplication des problèmes environnementaux liés à l’agriculture industrielle, les cultures de couverture s’imposent de plus en plus dans de nombreux endroits à travers le monde comme un outil pour mitiger ces problématiques. Il existe une panoplie d’espèces végétales utilisées dans ce but, comme le seigle (Secale cereale), le radis fourrager (Raphanus sativus) ou le pois fourrager (Pisum sativum) et bien d’autres encore. Elles peuvent servir de nombreux objectifs comme lutter contre les mauvaises herbes, réduire les pertes de sol et d’éléments nutritifs, fixer l’azote atmosphérique ou favoriser la vie du sol (Sustainable Agriculture Network, 1998). Il y a une vraie diversité d’usage et de techniques à travers le monde.
Le semis de ray-grass annuel (Lolium multiflorum) intercalaire dans le maïs y est une pratique connue au Québec. Le trèfle incarnat (Trifolium incarnatum) est de son côté une légumineuse qui est en symbiose avec des bactéries Rhizobium qui fixent l’azote atmosphérique, d’où l’idée de combiner les deux cultures dans un intercalaire dans la culture du maïs. Cependant, la pratique représente certaines contraintes au niveau du désherbage, notamment quant à la compatibilité d’une graminée et d’une légumineuse utilisées simultanément, avec l’emploi d’herbicides résiduels. Ainsi, l’établissement d’un intercalaire de ray-grass dans le maïs s’effectue généralement suite à une application de glyphosate en postlevée du maïs. Cependant, au moment où les cas de plantes résistantes au glyphosate se multiplient un peu partout dans le monde et commencent même à apparaître au Québec (RAP., 2019), il faut chercher à diversifier les méthodes de désherbage et ce en particulier dans la culture du maïs. Il serait dommage de perdre les gains pour l’environnement et la santé des sols procurés par ce mélange d’intercalaire du fait d’une baisse de l’efficacité du glyphosate. Les approches pour varier le désherbage sont nombreuses, comme l’utilisation d’autres familles d’herbicides, le désherbage mécanique et la combinaison de ceux-ci en lutte intégrée. Il y a cependant des risques liés à la phytotoxicité et au désherbage insuffisant pour les intercalaires.
Chapitre 1 Revue de littérature
1.1 Les mécanismes de la compétition entre les plantes
1.1.1 Importance de la biomasse dans le contrôle des mauvaises herbes
Le facteur déterminant pour le contrôle des mauvaises herbes est la biomasse de l’intercalaire (Bybee-Finley et coll., 2017). Dans le contexte d’une expérience répétée sur deux ans, avec du sorgho (Sorghum bicolor × S. sudanense), du millet perlé (Pennisetum glaucum), du niebé (Vigna unguiculata) et du crotalaire effilé (Crotalaria juncea) cultivés en monoculture et en mélange de trois ou quatre espèces, les biomasses de mauvaises herbes sont généralement significativement moins importantes lorsque la biomasse de la culture est plus élevée (Bybee-Finley et coll., 2017). Aussi, les monocultures de légumineuses semblent généralement moins compétitives face aux mauvaises herbes (Bybee-Finley et coll., 2017). Cependant, ce facteur semble être uniquement relié à la plus faible biomasse de la monoculture de légumineuses (Bybee-Finley et coll., 2017). Il y a aussi une faible corrélation négative entre le nombre d’espèces de l’intercalaire et la biomasse des mauvaises herbes mais seulement quand des graminées sont ajoutées aux légumineuses (Bybee-Finley et coll., 2017).
Donc, la biomasse totale de la culture intercalaire ressort dans une étude comme étant le facteur déterminant de la réduction de la pression de mauvaises herbes (Bybee-Finley et coll., 2017). En effet, la transmittance de PPFD (la densité de flux de photon lumineux) décline de façon exponentielle avec l’augmentation de biomasse de résidus (Teasdale et Mohler, 1993). Ainsi, plus il y a de biomasse de la culture ou de débris, plus la lumière est interceptée par celle-ci (Teasdale et Mohler, 1993). La lumière n’est donc plus disponible aux mauvaises herbes sous la culture pour assurer leur croissance (Teasdale et Mohler, 1993). Aussi, de la vesce velue vivante (Vicia villosa) contrôle mieux et plus longtemps que de la vesce velue desséchée (Teasdale et Daughtry, 1993). Ce meilleur contrôle est relié à la plus grande extinction de la lumière, au plus bas ratio de rouge sur le rouge lointain et à la plus faible amplitude de température diurne sous la vesce velue vivante que sous la vesce desséchée (Teasdale et Daughtry, 1993). Donc, un semis de culture intercalaire aurait avantage à avoir une grande biomasse et idéalement à être vivante pour maximiser le contrôle des mauvaises herbes.
1.1.2 Synergie entre les légumineuses et les graminées
La maximisation de la biomasse d’une culture étant prioritaire au contrôle des mauvaises herbes, il est important de favoriser les associations de plantes permettant d’atteindre cet objectif. Ainsi, une
plus grande biomasse est obtenue en serre de manière plus constante en utilisant un mélange de légumineuse et de ray-grass vivace qu’en utilisant une légumineuse en monoculture (Vasileva et coll., 2017). La plus grande productivité du mélange par rapport aux légumineuses seules est due à l’utilisation plus efficace de l’azote inorganique du sol par le mélange des deux types de racines (Vasileva et coll., 2017). On remarque donc une augmentation significative de la biomasse dans les mélanges par rapport aux monocultures de légumineuses, ce qui montre une synergie entre les légumineuses et les graminées (Vasileva et coll., 2017).
1.1.3 Utilisation de l’azote dans un contexte de compétition entre végétaux
L’azote du sol est un nutriment très important, tant pour la culture intercalaire que pour les mauvaises herbes. Saia et coll. (2016) ont évalué la productivité, la fixation de N2 atmosphérique
ainsi que la capacité d’être cultivées avec une graminée de plusieurs légumineuses de la région méditerranéenne. Il s’agit principalement d’espèces de légumineuses qui ne sont pas cultivées au Québec et le climat méditerranéen ne correspond pas aux conditions québécoises. Cependant, elles peuvent donner une idée du comportement de l’azote dans le contexte d’un mélange de trèfle- ray-grass, la graminée étant du ray-grass annuel. Certaines espèces de trèfles peuvent aussi avoir un intérêt sous nos conditions.
Des biomasses de cultures de couverture ont été 6,5 % plus élevées dans les mélanges que pour les légumineuses en culture pure en climat méditerranéen (Saia et coll., 2016). La proportion de légumineuses dans les mélanges peut être très variable, selon le type de légumineuses (Saia et coll., 2016). Il y a aussi une baisse de rendement total en azote dans les tissus végétaux dans les légumineuses en mélange par rapport à celles en culture pure (Saia et coll., 2016). Il y a cependant une majorité de l’azote fixée de l’atmosphère dans les légumineuses seules, mais aussi dans celles en mélange (Saia et coll., 2016). Le pourcentage de l’azote étant fixé de l’atmosphère est plus élevé dans les mélanges que dans les cultures pures de légumineuses (Saia et coll., 2016). De plus, il n’y a pas de transfert d’azote des légumineuses vers le ray-grass en raison de l’absence de différence significative en termes de teneur en azote avec des cultures annuelles sur une même année (Saia et coll., 2016).
De plus, les auteurs ont calculé le land equivalent ratio (LER), qui est la surface relative qu’il faudrait avoir de culture pure pour avoir le rendement du mélange. Avec un principe similaire, les auteurs ont aussi déterminé le nitrogen land equivalent ratio (NLER), qui est la surface de culture pure pour avoir le même rendement d’azote que le mélange. Ainsi, les mélanges de graminées et de légumineuses ont des ratios supérieurs à un, tant pour le LER que pour le NLER. Alors, il y a un
avantage en termes d’acquisition de biomasse, qui est le facteur le plus déterminant du contrôle des mauvaises herbes ainsi que sur la fixation totale d’azote (Saia et coll., 2016).
Donc, les mélanges de ray-grass et de légumineuses permettent une meilleure exploitation des ressources en azote du sol et limitent les pertes en azote du sol (par dénitrification, lessivage ou volatilisation) (Saia et coll., 2016). Alors, du point de vue des ressources du sol, la graminée permet d’exploiter au maximum les ressources en azote du sol, alors que la légumineuse utilise sa symbiose avec les Rhizobium sp. pour subvenir à ses besoins en azote (Saia et coll., 2016). La légumineuse est donc stimulée à fixer l’azote atmosphérique en raison de la rareté d’azote dans le sol, due à la graminée (Saia et coll., 2016). De plus, l’effet inhibiteur de la fertilisation azotée sur la formation de nodules fixateurs d’azote chez la féverole (Vicia faba) peut être diminué par la présence d’une graminée, forte consommatrice d’azote (Li et coll., 2009). Ainsi avec le mélange, une plus grande quantité d’azote est fixée. Cependant, il n’y a pas vraiment plus d’azote qui reste disponible dans le sol pour subvenir aux besoins des mauvaises herbes. Faisant partie de la biomasse de la culture de couverture, l’azote pourra subséquemment redevenir disponible pour la culture suivante.
1.1.4 Allélopathie
Les plantes peuvent produire des composés toxiques pour les autres plantes, lesquelles agissent par de multiples voies comme l'inhibition de la division et de l'élongation cellulaires et de la synthèse des protéines, la modification de la perméabilité membranaire et de l'absorption minérale, l’inhibition de la photosynthèse et de la respiration, ou encore en interférant avec l’action des phytohormones (Doré et coll., 2004). Les composantes toxiques peuvent également influencer la fixation biologique de l'azote, en diminuant le nombre et la taille des nodosités, leur teneur en leghémoglobine, et en réduisant la croissance des rhizobiums et celle de la culture hôte (Doré et coll., 2004). Cependant, trois principaux facteurs rendent difficile l’identification et par la suite l’application de ce principe au champ (Doré et coll., 2004). Premièrement, il est difficile de faire la différence entre compétition pour les ressources et allélopathie (Doré et coll., 2004). Deuxièmement, même s’il y a présence d’un composé phytotoxique démontré en laboratoire, il est incertain qu’il sera efficace sous les conditions du champ (Doré et coll., 2004). Troisièmement, les microorganismes ou leur absence peuvent jouer sur la phytotoxicité, en l’empêchant ou en produisant des métabolites secondaires phytotoxiques, à partir des molécules émises par les plantes (Doré et coll., 2004).
1.2 Les cultures intercalaires et l’environnement
1.2.1 Réduction des pertes de sol
Réduire les pertes de sols, surtout dans une culture de maïs avec de larges entre-rangs, représente un avantage intéressant pour réduire la pollution agricole diffuse. Cela permet de retenir le sol fertile ainsi que les nutriments qu’il contient aux champs, en plus de réduire la pollution que ces pertes peuvent occasionner, notamment dans les cours d’eau. Un intercalaire de trèfle Kura (Trifolium ambiguum) dans une culture de maïs ensilage peut réduire de 50 % le ruissellement et de 77 % les pertes de sol (Siller et coll., 2016). Du seigle (Secale cereale), semé à la suite de la récolte du maïs peut réduire le ruissellement de 67 % et les pertes de sol de 81 % (Siller et coll., 2016).
1.2.2 Réduction de la diffusion de contaminants agricoles
Par rapport au contrôle des pertes d’herbicides dans l’environnement, Gaynor et coll. (2001) ont évalué, de 1992 à 1995, les effets d’un intercalaire de ray-grass, du travail de sol et du contrôle du drainage et de la subirrigation sur les pertes d’atrazine, de métribuzine et de métolachloredans la culture du maïs. Ainsi, un intercalaire de ray-grass pris seul semble abaisser les pertes de ces trois herbicides utilisés dans le maïs (Gaynor et coll., 2001). On observe la même tendance pour le travail réduit par rapport au travail conventionnel du sol (Gaynor et coll., 2001). Cependant, pour les trois herbicides, le travail réduit combiné à l’intercalaire de ray-grass a toujours montré des pertes significativement plus faibles que le travail conventionnel sans intercalaire (Gaynor et coll., 2001). Donc, l’intercalaire de ray-grass combiné au travail de sol réduit peut contribuer à diminuer la perte par le ruissellement en surface des herbicides (Gaynor et coll., 2001). Les précipitations doivent cependant être assez importantes pour évaluer cette diminution du ruissellement (Gaynor et coll., 2001). Donc, un intercalaire peut réduire le ruissellement et la perte de sol. Pour ce qui est de la perte d’atrazine, de métolachlore et de métribuzine, le ray-grass peut permettre de réduire leur perte s’il est combiné avec un travail réduit du sol
1.2.3 Réduction de la fertilisation azotée
Les légumineuses ont la possibilité de fixer l’azote atmosphérique, lequel peut par la suite redevenir disponible pour une culture principale (Sustainable Agriculture Network, 1998). Sur une moyenne de quatre ans, il est possible d’obtenir dans l’orge un rendement comparable d’une monoculture fertilisée avec de l’azote inorganique et d’une culture d’orge (Hordeum vulgare) non fertilisée mais intercalée de pois fourrager (Pisum sativum) (Jensen, 1996). Dans le maïs, sur une période de quatre
ans, le rendement peut être augmenté par la présence d’une légumineuse intercalaire comme le trèfle rouge (Trifolium pratense) (Scott et coll., 1987). Une graminée intercalaire, comme le ray-grass va plutôt consommer l’azote qu’en produire, mais peut capter les surplus de fertilisation azotée et la remettre en partie disponible lors de la sénescence de ses tissus (Sustainable Agriculture Network., 1998.
1.3 Le ray-grass
1.3.1 Caractéristiques du ray-grass
Le terme «ray-grass» est un terme assez général. Il ne désigne pas une seule espèce en particulier, mais bien un genre regroupant plusieurs espèces possédant des caractéristiques bien différentes. Les deux principales espèces sont le ray-grass annuel ou italien (Lolium multiflorum) et le ray-grass vivace (Lolium perenne) (Sarrantonio, 1994). Le genre Lolium fait partie de la famille des Poacea et est originaire d’Europe (Sarrantonio, 1994).
Aussi, le ray-grass italien est une graminée cespiteuse qui ne produit pas de rhizome (Hall, 1992). De plus, il fleurit lorsque l’ensoleillement dépasse 11 heures par jour (Hall, 1992). Il ne survit généralement pas plus d’une saison en climat nordique. Les cultivars peuvent cependant être divisés selon leur floraison, soit en alternatif et en non alternatif (Breune, 2014). En effet, le cultivar non alternatif ne produit pas de semence l’année d’implantation alors que l’alternatif, aussi connu sous le nom de ‘Westerwold’, peut ou non produire des graines lors de l’année d’établissement, ce qui peutainsi laisser une repousse l’année suivante (Breune, 2014).
Le ray-grass vivace est aussi une plante cespiteuse sans rhizome et sa floraison nécessite une période de dormance à des températures froides (Hall, 1992). De plus, les variétés de ray-grass, tant vivace qu’italien, peuvent être séparées en deux groupes selon leur ploïdie, c’est-à-dire le nombre de chromosomes (Hall, 1992). Il y a les diploïdes et les tétraploïdes (Hall, 1992). Dans les diploïdes, chaque chromosome est présent en deux copies alors que dans les tétraploïdes, c’est quatre copies (Hall, 1992). Ainsi, le ray-grass vivace tétraploïde possède de plus larges feuilles, produit moins de talles, mais des plus grosses et une croissance plus ouverte, entraînant une moins grande couverture de sol (Hall, 1992). Pour ce qui est de la semence et de la plantule, celle du tétraploïde est plus grosse, mais la croissance et la persistance des diploïdes sont plus importantes (Hall, 1992).
1.3.2 Culture du ray-grass
Pour ce qui est de la croissance, les deux espèces ont une croissance rapide, mais le ray-grass annuel s’établit plus rapidement (Sarrantonio, 1994). Ce dernier peut atteindre de 60 à 120 cm alors que le vivace peut atteindre de 30 à 90 cm de hauteur et persister de 3 à 4 ans dans le Nord-est des États-Unis (Sarrantonio, 1994). Aussi, les deux espèces ont un système racinaire dense, fibreux et profond, sont de gros consommateurs d’azote et d’eau, sont vulnérables aux conditions sèches et aux hautes températures et sont versatiles quant au type de sol sur lequel ils poussent (Sarrantonio, 1994). Ainsi, le ray-grass arrête sa croissance lorsque la température atteint 22-25℃, puis la reprend avec vigueur après la récolte du maïs (Breune et coll., 2015).
Quelques conclusions peuvent être tirées avec ces informations quant à l’usage en mélange du ray-grass avec le trèfle dans une culture de maïs, on peut tirer quelques conclusions avec ces informations. D’abord, les deux espèces du genre Lolium sont bien à l’abri de la chaleur excessive et du manque d’humidité dans les entre rangs de maïs. De plus, le sol d’un champ de maïs est généralement bien fertilisé, en raison des besoins élevés en azote de ce dernier. Cependant, si la fertilisation s’effectuet uniquement sur le rang de maïs, l’entre-rang peut être pauvre en azote d’où l’intérêt d’y semer une plante fixatrice d’azote atmosphérique. Ensuite, le développement agressif des ray-grass en fait de bons compétiteurs face aux mauvaises herbes. Aussi, le cycle de vie annuel convient mieux à l’utilisation dans l’entre-rang du maïs, qui est une culture annuelle. Ainsi, en établissant une espèce annuelle dans les entre-rangs, on réduit les risques de la voir réapparaître l’année suivante, surtout si elle a été détruite par le gel ou par un contrôle mécanique ou chimique. Sa gestion en est ainsi facilitée. De plus, son établissement et sa croissance plus rapides la première année rendent le ray-grass annuel plus compétitif face aux mauvaises herbes que le ray-grass vivace lors de son établissement.
Cependant, le ray-grass n’a presque pas d’activité allélopathique (Fujii, 2001). Si on le compare au seigle (Cereale secale), qui possède un pourcentage d’inhibition de 71 %, le ray-grass annuel n’obtient que 26 % (Fujii, 2001). Sa capacité à lutter contre les mauvaises herbes est donc essentiellement reliée à la compétition pour la lumière et les ressources.
En raison de ces différentes caractéristiques, l’étude s’arrête sur le choix du ray-grass annuel en mélange avec le trèfle incarnat.
1.4 Trèfle incarnat
1.4.1 Caractéristiques du trèfle incarnat
Il est important de caractériser le trèfle incarnat. Pour déterminer certaines caractéristiques utiles de différentes espèces de trèfles, dont de trèfle incarnat, dans un contexte de culture de couverture pour lutter contre les mauvaises herbes, den Hollender et coll. (2007) ont évalué huit espèces de trèfles. Le tableau 1 montre que le trèfle incarnat arrive à sa croissance maximale plus lentement que le trèfle perse ou le trèfle d’Alexandrie, qui croissent plus rapidement. Il est comparable pour cette caractéristique au trèfle alsike, qui est plus lent à arriver à maturité que les trèfles rouge, sous-terrain et blanc (den Hollender et coll., 2007). Le trèfle incarnat produit aussi une biomasse totale supérieure aux trèfles perse, rouge sous-terrain et blanc, mais comparable au trèfle d’Alexandrie et inférieure au trèfle Alsike (den Hollender et coll., 2007).
Tableau 1 Vitesse d'atteinte de la croissance maximale et biomasse maximale de diverses espèces de trèfles.
Espèce de trèfle M (jours après semis)1 C (g m-2)1
Trèfle Alsike 84a 773,5 a (Trifolium hybridum) Trèfle d’Alexandrie 67c 535,1bc (Trifolium alexandrinum) Trèfle incarnat 76ab 633,7b (Trifolium incarnatum) Trèfle perse 54d 422,5d (Trifolium resupinatum) Trèfle rouge 71bc 520,0cd (Trifolium pratense) Trèfle sous-terrain 71bc 448,5cd (Trifolium subterraneum) Trèfle blanc (large)
76ab 469,7cd
(Trifolim repens) Trèfle blanc (moyen)
75bc 424,9d
(Trifolium repens)
Adapté de den Hollender (2007); 1 M = atteinte de la croissance maximale; C = biomasse sèche maximale m-2.
De plus, en culture de couverture, en compétition avec de la moutarde des champs (Sinapis arvense), le trèfle incarnat produit une moins grande biomasse et contrôle moins bien cette mauvaise herbe que le trèfle d’Alexandrie et Alsike (Ross et coll., 2001). Cependant, il produit une biomasse et permet un contrôle significatif de la moutarde, comparativement à un témoin enherbé (Ross et coll., 2001).
Le trèfle incarnat germe et pousse rapidement en conditions fraiches et humides où il peut former un couvert dense qui compétitionne les mauvaises herbes (Sarrantonio, 1994). Aussi, il pousse bien dans tous les types de sols si le drainage est convenable, mais performe particulièrement bien dans
les sols loameux avec beaucoup d’humus. Il possède une bonne tolérance à l’ombre et s’acclimate aux pH compris entre 5,0 et 7,0 (Sarrantonio, 1994). Néanmoins, il est peu tolérant à la chaleur et à la sécheresse (Sarrantonio,1994).
À la lumière de ces informations, le trèfle incarnat, en raison de sa vigueur, de sa tolérance à l’ombre qui est essentielle sous le maïs, de ses propriétés allélopathiques et de sa versatilité quant au type de sol et de pH, ressort généralement comme la meilleure espèce de trèfle pour un mélange intercalaire avec le ray-grass dans le maïs. Dans un champ mal drainé, le trèfle Alsike pourrait cependant mieux performer. Sarrantonio (1994) recommande, lorsqu’utilisé en mélange, une dose de semis de 1,8-2,6 kg/ha pour le trèfle incarnat et de 0,6-0,9 kg/ha pour le ray-grass annuel.
Le trèfle incarnat possède un pourcentage d’inhibition des mauvaises herbes de 64 %, confirmant ainsi son potentiel allélopathique (Fujii, 2001). De plus, White et coll. (1989) ont évalué le potentiel allélopathique de résidus et d’extraits aqueux de légumineuses sur différentes plantes, dont le maïs, le ray-grass, la gloire du matin (Ippomoea sp.) et le trèfle incarnat (Trifolium incarnatum). Du sol provenant d’une parcelle ayant supporté l’année précédente une culture de trèfle incarnat n’a pas d’effet phytotoxique résiduel sur les mauvaises herbes lorsque les résidus de trèfle incarnat sont retirés (White et coll., 1989). Cependant, un extrait aqueux de trèfle incarnat peut grandement réduire la germination de moutarde des champs et de gloire du matin (White et coll., 1989). Le ray-grass peut aussi y être sensible, mais pas le maïs (White et coll., 1989). Cela montre bien la présence d’une toxine permettant d’inhiber principalement la germination de mauvaises herbes, mais aussi celle du ray-grass. Cependant, il est difficile d’extrapoler les effets de ce seul résultat dans un contexte aux champs, l’extrait aqueux étant étudié en conditions de laboratoire. En champs, des expériences montrent que la levée du maïs est faiblement inhibée lorsque les débris de trèfle sont incorporés et pas du tout, lorsqu’ils sont laissés en surface (White et coll., 1989). La croissance du maïs n’a pas été diminuée significativement par les résidus incorporés et peut être augmentée par les résidus laissés en surface (White et coll., 1989). Donc, le trèfle incarnat possède une substance allélopathique soluble dans l’eau avec peu d’effets négatifs sur le maïs (White et coll., 1989). Le but des producteurs agricoles étant d’avoir un rendement élevé en maïs-grain, il est important qu’un éventuel composé allélopathique ne soit pas toxique pour cette culture.
1.4.2 Culture du trèfle incarnat
Sous les conditions du Nord-Est américain, le trèfle incarnat est utilisé comme culture annuelle d’été, bien qu’il puisse être utilisé comme une annuelle d’hiver dans le sud de la Pennsylvanie (Sustainable Agriculture Network, 1998). Ce trèfle a besoin de conditions fraîches et humides, ce
qui limite son utilisation dans les zones plus sèches et chaudes (Sustainable Agriculture Network, 1998). Cette culture peut pousser dans tous les types de sols bien drainés, particulièrement les loams sableux, mais tolère mal les argiles lourdes, les sols saturés en eau, plus acides qu’un pH de 5 ou encore alcalins. (Sustainable Agriculture Network, 1998). Les carences en phosphore et en potassium empêchent aussi la fixation d’azote atmosphérique par cette plante (Sustainable Agriculture Network, 1998). Aussi, cette culture fournit une source d’azote au sol, produit une bonne biomasse, peut fournir du nectar aux insectes pollinisateurs et des abris aux prédateurs, contribue à prévenir le lessivage d’azote et de pesticides, et ce particulièrement lorsqu’associée au ray-grass annuel (Sustainable Agriculture Network, 1998). Le trèfle incarnat est aussi utilisé en compagnie d’autres cultures, poussant bien avec les céréales, les petites graminées et les autres trèfles (Sustainable Agriculture Network, 1998). Il est aussi utilisé en culture de couverture pure, en rotation avec d’autres cultures pouvant bénéficier d’une légumineuse, comme la pomme de terre (Solanum tuberosum), ou suite à une culture de céréale pour un couvert automnal ou avant le semis du maïs dans les climats plus chauds (Sustainable Agriculture Network, 1998). C’est aussi une culture utilisée en pâturage annuel et peut repousser s’il est brouté ou tondu avant l’apparition des bourgeons (Sustainable Agriculture Network, 1998). Pour s’en débarrasser, le désherbage mécanique est très efficace et une tonte lorsque la plante est en fleur peut être suffisante pour la détruire (Sustainable Agriculture Network, 1998). De plus, la métribuzine, le glufosinate, le paraquat, la cyanazine et le glyphosate peuvent la détruire chimiquement (Sustainable Agriculture Network, 1998).
1.5 Implantation d’un intercalaire
D’abord, il est important de bien sélectionner le stade de la culture pour le semis de l’intercalaire afin de pouvoir réduire ou éliminer les impacts négatifs possibles de l’intercalaire sur la culture de maïs et également tenter de maximiser la biomasse de l’intercalaire pour profiter au maximum de ses bénéfices. Le semis du ray-grass au stade 5-8 feuilles du maïs est sécuritaire pour le rendement de ce dernier et permet au ray-grass d’atteindre le stade 3-4 feuilles, théoriquement nécessaire à sa survie sous le maïs (Breune et coll., 2015). D’autres études suggèrent qu’un semis de ray-grass intercalaire au stade 4-6 feuilles du maïs ne réduit pas le rendement du maïs (Scott et coll., 1987). Cependant, un semis de ray-grass intercalaire au stade 11 feuilles du maïs l’empêche de se développer suffisamment avant la fermeture du couvert par le maïs, ce qui réduit ainsi grandement la biomasse produite (Breune et coll., 2015). Le trèfle incarnat a aussi été semé avec succès au Michigan en culture intercalaire dans le maïs lorsque ce dernier a atteint de 40 à 60 cm de hauteur (Sustainable Agriculture Network, 1998). Le trèfle incarnat peut cependant être semé en culture
pure après que les derniers risques de gels sont passés et produire ainsi des plantules robustes (Sustainable Agriculture Network, 1998). Sous ces conditions et à ce stade de semis, le trèfle a accumulé en fin de saison 270 kg MS/ha et 9 kg N/ha et n’a pas occasionné de pertes de rendement au maïs, ce qui semble très prometteur pour cette utilisation (Sustainable Agriculture Network, 1998).
Pour la méthode de semis de l’intercalaire, il existe deux méthodes principales, soit à la volée ou incorporé (Piché Dumontier, 2017). Le semis à la volée peut s’effectuer avec un épandeur à engrais ou un semoir porté (Piché Dumontier, 2017). Le semis d’intercalaire peut être effectué avec un semoir à la volée avec des tubes de descente (Piché Dumontier, 2017). L’incorporation peut se faire grâce à l’ajout de peignes ou d’étançons munis de soc au semoir (Piché Dumontier, 2017). De plus, incorporer le semis de ray-grass permet de favoriser la germination et d’augmenter significativement la biomasse sèche aérienne de ray-grass (Piché Dumontier, 2017)
1.6 Désherbage mécanique en intercalaire
1.6.1 Outils de désherbage mécanique
De nombreux outils peuvent être utilisés dans le contrôle mécanique des mauvaises herbes en grandes cultures. Parmi ceux-ci, trois types sont principalement utilisés : la houe rotative, la herse-peigne et le sarcleur.
La houe rotative est constituée de roulettes avec des extrémités en formes de cuillère qui pulvérisent le sol. Elle déracine, mutile et enterre ainsi les mauvaises herbes. Elle est conçue pour les sols labourés avec peu de résidus de récolte (Coulombe et Douville, 2000). Elle est efficace contre les mauvaises herbes aux stades fils blancs et cotylédons et perd de l’efficacité à partir du stade d’une feuille des mauvaises herbes (Coulombe et Douville, 2000). Elle peut être utilisée en prélevée, à condition que le coléoptile de maïs soit à plus de 3 cm souse la surface et en postlevée jusqu’au stade 6 feuilles du maïs (Coulombe et Douville, 2000). Elle peut être utilisée à des vitesses de 10 à 20 km/h sous peine de voir son efficacité diminuée (Coulombe et Douville, 2000). Finalement, la houe ne se montre pas efficace dans les sols où il ne se forme pas de croûte à la surface du sol. Elle permet de désherber le rang et l’entre-rang simultanément, mais n’est pas efficace contre la plupart des mauvaises herbes vivaces comme le chiendent (Coulombe et Douville, 2000).
La herse-peigne est composée de dents vibrantes à ressort qui brassent le sol et déracinent, mutilent ou enterrent les mauvaises herbes à une profondeur d’environ un 2,5 cm (Coulombe et Douville, 2000). Elle est conçue pour les sols légers avec peu de résidus où elle s’avère plus performante que
la houe rotative (Coulombe et Douville, 2000). Elle permet d’avancer à une vitesse allant de 6 à 12 km/h en postlevée et 10 à 15 km h-1 en prélevée (Coulombe et Douville, 2000). Elle est efficace contre les mauvaises herbes au stade fil blanc, cotylédons et première vraie feuille, mais elle n’est plus efficace au-delà (Coulombe et Douville, 2000). Relativement aux stades du maïs où elle peut être utilisée, elle est efficace en prélevée jusqu’à ce que le coléoptile de maïs soit à 3 cm sous la surface du sol, ceci pour éviter qu’il ne soit endommagé par les dents. Elle peut être de nouveau utilisée au stade 4 feuilles du maïs (Coulombe et Douville, 2000). Finalement, elle permet de désherber le rang et l’entre-rang simultanément mais n’est pas efficace contre les mauvaises herbes vivaces (Coulombe et Douville, 2000).
Le sarcleur entre les rangs consiste en des socs à patte d’oie qui déracinent et enterrent les petites mauvaises herbes et sectionne les plus grosses (Coulombe et Douville, 2000). Il en existe de plusieurs types : léger, mi-lourd et lourd et avec différents types de dents (Coulombe et Douville, 2000). En général, plus ils sont lourds, plus leur travail et agressif et permet de lutter contre des mauvaises herbes aux stades plus avancés (Coulombe et Douville, 2000). Généralement, les sarcleurs peuvent être utilisés à une vitesse allant de 6 à 10 km/h, ce qui est inférieure à la houe et la herse-peigne et ne permettent qu’en général le désherbage de l’entre-rang et non du rang lui-même, mais peuvent ralentir le développement de certaines vivaces comme le chiendent (Elymus repens) ou la prêle des champs (Equisetum arvense) (Coulombe et Douville, 2000).
1.6.2 Désherbage mécanique avec intercalaire aux champs
Abdin et coll. (2000) ont évalué le succès de différentes cultures intercalaires semées dans les entre-rangs de maïs suite au contrôle mécanique des mauvaises herbes dans deux sites au Québec (tableau 2).
Tableau 2 Biomasse sèche de mauvaises herbes et rendement en grains pour une culture de maïs-grain intercalé et désherbé mécaniquement.
Culture Biomasse sèche de mauvaise herbe dans l’entre rang à la deuxième semaine d’août (g m-2) Rendement en grains (Mg ha-1) Seigle d’automne 4,8b 8,8a Vesce velue 4,1b 9,0a
Trèfle rouge + ray-grass 1,7bc 9,3a
Trèfle blanc +ray-grass 2,9b 8,8a
Trèfle sous-terrain 2,4bc 8,7a Mélilot jaune 2,6bc 8,7a Trèfle Perse 3,9b 8,6a Trèfle fraisier 5,4ab 8,9a Trèfle incarnat 2,2b 9,7a luzerne 2,5b 8,8a Trèfle d’Alexandrie 1,8bc 9,4a
Travail mécanique seul 1,2bc 9,4a
Désherbé à la main 0,2c 8,9a
Contrôle chimique 1,2bc 8,7a
Témoin enherbé 10,7a 7,0b
Adapté de Abdin et coll. (2000).
Au tableau 2, on constate que les biomasses sèches de mauvaises herbes mesurées à la deuxième semaine d’août sont comparables pour toutes les cultures intercalaires précédées d’un désherbage mécanique, mais sont aussi identiques au traitement de contrôle mécanique uniquement (Abdin et coll., 2000). Donc, le ray-grass accompagné de trèfle blanc ou rouge et le trèfle incarnat ne permettent pas de réduire la biomasse de mauvaises herbes suite à un désherbage (Abdin et coll., 2000). C’est la qualité du désherbage initial qui détermine la biomasse de mauvaises herbes et non la culture intercalaire (Abdin et coll., 2000). Les rendements en maïs-grain ne sont pas augmentés ou diminués par la présence de cultures intercalaires (Abdin et coll., 2000). On peut en conclure que les intercalaires ne réduisent pas significativement les rendements du maïs par rapport au témoin désherbé. Ainsi, dans le cadre de cette expérience, les intercalaires ont été inutiles contre les mauvaises herbes présentes au début août, car elles n’ont pas significativement réduit les mauvaises herbes suite au contrôle mécanique.
Cependant, l’expérience d’Abdin et coll. (2000) ne présente pas de résultats sur les mauvaises herbes plus tard en saison. En effet, le contrôle mécanique des mauvaises herbes peut promouvoir l’émergence de mauvaises herbes (Sparks et coll., 1995). Ainsi, ces dernières n’ont pas été prises en compte par Abdin et coll. (2000). Donc, l’efficacité du contrôle mécanique est généralement
inférieure à celle du désherbage chimique, augmentant la pression de mauvaises herbes et donc la banque de semence de mauvaises herbes dans le sol (Gallandt ,2014).
Aussi, Bilalis (2010) a utilisé des intercalaires de pois chiche et de fève (Phasoleus vulgaris) dans le maïs en agriculture biologique suite à un contrôle mécanique des mauvaises herbes. Sur la figure 1, adaptée de Bilalis et coll. (2010), il est visible qu’à la suite d’un semis de maïs, il y a peu de différence entre un maïs avec ou sans intercalaire; une différence peut cependant apparaître avec l’avancement de la saison de croissance (Bilalis et coll., 2010). Une culture intercalaire de légumineuses peut diminuer la biomasse de mauvaises herbes suite à un désherbage mécanique dans le maïs (Bilalis et coll., 2010). Cela peut donc être un atout en agriculture biologique, à condition que l’intercalaire soit suffisamment compétitive avec les mauvaises herbes.
Figure 1 : Biomasse de mauvaises herbes suite à un désherbage mécanique dans une culture de maïs intercalé d'une légumineuse ou non. Adaptée de Bialis et coll. (2010)
Aussi, en régie biologique, le passage du sarcleur peut être une contrainte temporelle importante pour le semis d’intercalaire et son développement (Laliberté, 2014). En effet, le passage du sarcleur peut reporter le semis au stade 10 à 12 feuilles du maïs, empêchant ainsi le bon développement du ray-grass (Laliberté, 2014). 0 5 10 15 20 25 30 35 40 60 80 100 120 140 Bi oma ss e d e ma uv ai se s h erb es (g /m 2)
jours après le semis
Maïs Maïs+fève Maïs+pois chiche
1.7 Désherbage chimique en prélevée
1.7.1 Atrazine
L’atrazine est un herbicide du groupe 5, homologuée au Canada dans la culture du maïs (Fortin, 2017). C’est un inhibiteur du photosystème II qui se lie à des sites de liaison du domaine Qb,
bloquant ainsi le transport des électrons, ce qui inhibe la photosynthèse (Fortin, 2017). Les triazines se déplacent dans le xylème et le phloème des feuilles des plantes et s’accumule dans les points de croissance (Fortin, 2017). L’atrazine est mobile dans la plante et absorbé via les feuilles mais surtout par les racines. L’atrazine est appliquée au sol est absorbée par la plante par la suite (Fortin, 2017).
L’atrazine est efficace contre une large gamme de mauvaises herbes à feuilles larges et contre la folle-avoine (Avena fatua) (Bayer, 2018a). Le maïs lui y est tolérant grâce à sa capacité à métaboliser rapidement l’atrazine en un composé non phytotoxique (Fortin, 2017). La moutarde des oiseaux (Brassica rapa) a été la première mauvaise herbe résistante à l’atrazine répertoriée au Québec (Fortin, 2017). Il existe au Québec des cas confirmés de résistances à l’atrazine chez la moutarde des oiseaux, le chénopode blanc (Chenopodium album), et l’amarante à racine rouge (Amaranthus retroflexus) (Fortin,2017). L’atrazine a une faible toxicité aiguë mais possède une toxicité à long terme extrêmement élevée (SAgE Pesticides, 2018). Elle a notamment montré des effets neurotoxiques et une toxicité endocrinienne chez les animaux (SAgE Pesticides, 2018).
1.7.2 Mésotrione
La mésotrione (vendue sous le nom commercial de CALLISTO® par exemple) est un herbicide du groupe 27, de la famille des tricétones (Syngenta, 2018). Elle peut être utilisée pour la répression de plusieurs espèces de dicotylédones et de graminées dans le maïs en prélevée et en postlevée (Mitchell et coll., 2001). Elle est dérivée d’une phytotoxine naturellement produite chez une plante de la famille des Myrtacées originaire d’Australie : Callistemon citrinus (Mitchell et coll., 2001). La mésotrione agit par l’inhibition compétitive de l’HPPD synthase, une enzyme qui fait partie de la voie de la conversion de la tyrosine, un acide aminé, en plastoquinone, elle-même un cofacteur de l’enzyme phytoène désaturase, une enzyme nécessaire à la formation des caroténoïdes (Mitchell et coll., 2001). Le maïs est naturellement résistant à la mésotrione, puisqu’il est capable de la métaboliser rapidement en sous-produits non toxiques (Mitchell et coll., 2001).
La mésotrione résiste à l’hydrolyse à tous les pH rencontrés dans les sols (SAgE pesticides, 2018). Sa dégradation s’effectue par photolyse à la surface du sol (SAgE pesticides, 2018). Sa demi-vie à la surface du sol est de 28,9 jours et dans l’eau, elle est de 86 à 96 jours (SAgE pesticides, 2018). Elle est de faiblement à modérément persistante dans les sols. En effet, sa demi-vie y est, en condition aérobie de 8 à 31,5 jours et en anaérobiose, de 3,6 à 11 jours (SAgE pesticides, 2018). Elle a une valeur de Koc variant de 39 à 70 mg l-1, la rendant mobile dans les sols avec ainsi un potentiel de lessivage élevé et un risque de contaminer l’eau (SAgE pesticides, 2018). Aussi, l’adsorption de la mésotrione sur les sols est reliée principalement au pH du sol et dans une moindre mesure, à la matière organique dans le sol (Dyson et coll., 2002). Lorsque le pH augmente, la mésotrione est moins fortement adsorbée sur les sols et par conséquent sa demi-vie dans le sol diminue (Dyson et coll., 2002).
Elle a une faible toxicité aiguë, n’étant qu’irritante pour les yeux, mais sa toxicité à long terme est élevée (SAgE pesticide, 2018).
1.7.3 Isoxaflutole
L’isoxaflutole est un herbicide du groupe 27 (Fortin, 2017). L’herbicide provoque des blanchissements à jaunissements en affectant l’enzyme HPPD (p-hydroxyphényl pyruvate désoxygénase), laquelle intervient dans la synthèeses des plastoquinones (Pallet et coll., 1998) La plante semsible devient ainsi translucide et très vulnérable au soleil (Fortin, 2017). Cet herbicide est en général appliqué en présemis ou en prélevée et est absorbé par les racines et les jeunes pousses des mauvaises herbes (Fortin, 2017). Il est utilisé pour le contrôle de nombreuses espèces de feuilles larges et de graminées en pré-émergence dans la culture du maïs (Pallet et coll., 1998). Après absorption foliaire ou racinaire, il est rapidement converti en dikétonitrile (Pallet et coll., 1998). La susceptibilité des plantes à l’herbicide est corrélée à la quantité de dikétonitrile présente dans les tissus des bourgeons végétaux (Pallet et coll., 1998). La résistance du maïs à cet herbicide provient d’une métabolisation rapide du dikétonitrile en acide benzoïque inactif (Pallet et coll., 1998). Sa demi-vie au sol en condition aérobie est de 2 jours, ainsi il se dégrade rapidement dans les sols et n’y est pas persistant (SAgE pesticides, 2018). Il possède une k/oc variant de 93 à 123 mg/l, le rendant de très à modérément mobile dans les sols (SAgE pesticides, 2018). Il n’est cependant pas à risque de contaminer les eaux souterraines en raison de sa faible persistance dans les sols et de sa sensibilité à l’hydrolyse (SAgE pesticides, 2018). De plus, il est faiblement volatile avec une pression de vapeur de 1,0 x 10-6 Pa à 20 °C (SAgE pesticides, 2018). Dans les sols, le dikétonitrile tout comme l’isoxaflutole peut être adsorbé sur les argiles ainsi que sur la matière organique du sol
(Rice et coll., 2004). Par la suite, l’isoxaflutole et le dikétonitrile adsorbés sur les argiles peuvent être relâchés au courant de la saison (Rice et coll., 2004). Cependant, le dikétonitrile est plus facilement disponible que l’isoxaflutole en solution dans le sol en plus d’avoir une plus grande activité biologique, persistance et solubilité dans l’eau (Rice et coll., 2004). Il est donc plus à risque que l’isoxaflutole de causer des dommages aux cultures. Le cyprosulfamide permet cependant de réduire les dommages pouvant être causés au maïs et ainsi rendre plus sécuritaire cet herbicide (Robinson et coll., 2013). De plus, une augmentation du taux de matière organique dans le sol augmente la sorption de l’herbicide dans le sol (Rice et coll., 2004). Le pH aussi a un effet très prononcé, la baisse du pH du sol amenant une hausse de l’adsorption de l’isoxaflutole (Rice et coll., 2004). L’augmentation du taux d’argile et de matière organique augmente aussi la sorption du dikétonitrile, mais la baisse du pH favorise la désorption de ce composé secondaire (Rice et coll., 2004).
L’isoxaflutole n’a qu’une faible toxicité aiguë, mais la toxicité chronique est extrêmement élevée (SAgE pesticides, 2018). Il a en effet un effet cancérigène chez de nombreux mammifères et pourrait affecter le développement chez le chien et le rat (SAgE pesticides, 2018). En 2017, on ne recense aucun cas avéré de résistance aux isoxazoles (Fortin, 2017).
1.7.4 La tolérance des intercalaires aux herbicides de prélevée
Un des principaux facteurs pouvant nuire à l’établissement des cultures intercalaires dans le maïs peut être relié aux herbicides résiduels. En effet, ceux-ci peuvent être appliqués dans le maïs et contrôler pendant un certain temps les mauvaises herbes. Il est donc possible qu’ils interagissent avec le trèfle incarnat et le ray-grass, semés en intercalaire.
Le ray-grass intercalaire ne tolère pas bien la plupart des herbicides utilisés en prélevée dans le maïs à l’exception de l’atrazine et de l’isoxaflutole (Piché Dumontier, 2017). En effet, le S-métolachlore, le diméthénamide/saflufénacil, le pyroxasulfone et la pendiméthaline n’ont pas procuré de biomasse comparable au traitement de glyphosate en postlevée et au traitement désherbé à la main dans une expérience réalisée au Québec (Piché-Dumontier, 2017). L’atrazine et l’isoxaflutole ont cependant procuré des biomasses de ray-grass comparables au glyphosate en postlevée.
Aussi, Wallace et coll. (2017) ont analysé les effets de différents types d’herbicides en prélevée sur le ray-grass annuel et le trèfle rouge (Trifolium pratense). Les expériences se sont déroulées entre 2013 et 2015, dans le Mid-Atlantic américain, principalement sur des loams sableux et des loams limoneux.
Figure 2 Réduction ou augmentation de la biomasse par différents herbicides de prélevée sur le ray-grass annuel et le trèfle rouge.
Adaptée de Wallace et coll. (2017)
La figure 2 tirée de Wallace et coll. (2017) montre les effets des différents herbicides sur le ray-grass annuel et le trèfle rouge. La moyenne est le point noir et l’intervalle de confiance est la ligne noire, on peut donc en conclure que la réponse du trèfle rouge et du ray-grass a différé considérablement selon le type d’herbicide utilisé. Les dégâts causés par les chloroacétamides au ray-grass annuel étaient du plus fort dommage au plus faible : le S-métolachlor, le diméthènamid-P et l’acétochlor (Wallace et coll.,2017). Aucun dommage significatif des anti-feuilles larges (rimsulfuron, atrazine, mésotrione et saflufénacil) n’a été causé sur le ray-grass annuel (Wallace et coll., 2017). Pour ce qui est du trèfle rouge, les antigraminées, le saflufénacil, le rimsulfuron et l’atrazine n’ont pas causé de dommages significatifs (Wallace et coll., 2017). Cependant, la mésotrione peut causer des dommages supérieurs à 80 % au trèfle rouge (Wallace et coll., 2017). Donc, plusieurs types de mélanges d’herbicides résiduels sur un intercalaire de trèfle et de ray-grass pourraient être utilisés sur des loams limoneux, surtout en demi-dose pour réduire les possibles dommages : acétochlor + atrazine, acétochlor + rimsulfuron, et saflufénacil + diméthènamid-P (Wallace et coll., 2017). Sur un loam sableux, les dommages sont généralement plus importants que sur un loam limoneux (Wallace et coll., 2017).
Traitement
Biomasse de ray-grass annuel (proportion du témoin)
Biomasse de trèfle rouge (proportion du témoin)
1.8 Désherbage chimique en postlevée
1.8.1 Le Glyphosate
Le glyphosate est un herbicide du groupe 9 et c’est le seul herbicide appartenant à ce groupe (Fortin, 2017). Il existe sous différentes formes de sels, mais c’est toujours la même matière active (Fortin, 2017). Il inhibe l’action de l’EPSP synthase, une enzyme responsable de la synthèse de trois acides aminés aromatiques, le tryptophane, la tyrosine et la phénylalanine (Fortin, 2017). Le glyphosate est habituellement appliqué au feuillage en post-émergence des mauvaises herbes. C’est un herbicide de contact, systémique et non sélectif (Fortin, 2017). Il peut même être utilisé pour réprimer plusieurs vivaces (Fortin, 2017). Cependant, parce qu’il est non sélectif, des variétés de cultures OGM (Roundup Ready ®) ont été développées pour le tolérer (Fortin, 2017). Le glyphosate peut ainsi être utilisé sans risques pour un désherbage en postlevée dans une culture possédant cette technologie (Fortin, 2017).
De plus, le glyphosate est très fortement adsorbé par les sols, l’inactivant rapidement (Fortin, 2017). Sa koc étant de 500 à 6000 mg l
-1
, il est de légèrement mobile à immobile dans les sols, rendant son potentiel de lessivage faible (SAgE pesticides, 2018). Le glyphosate est, sous sa forme acide, de faiblement persistant à persistant dans les sols en condition aérobie (de 0,8 à 151 jours) (SAgE pesticides, 2018). Dans l’eau sa persistance est de faiblement persistant à persistant et il est stable à l’hydrolyse (SAgE pesticides, 2018). Il est aussi non volatile (SAgE pesticides, 2018). Aussi, son produit de dégradation est l’AMPA (acide aminométhylphosphonique), qui est aussi de faiblement persistant à persistant dans les sols en condition aérobie et dans l’eau (SAgE pesticides, 2018). Son potentiel de lessivage est aussi faible, car il est immobile à modérément mobile dans les sols (SAgE pesticides, 2018).
Sa toxicité aiguë varie de modérément à légèrement toxique, cependant, cette toxicité est plutôt reliée à la formulation qu’à la matière active (SAgE pesticide, 2018). On recense notamment de l’irritation chez le rat (SAgE pesticides, 2018). Pour ce qui est de la toxicité chronique, elle est faible et les principaux effets ont été constatés à des doses très élevées chez le rat et la souris (SAgE pesticides, 2018).
Pour ce qui est de la résistance, il y a à l’échelle planétaire 42 espèces de mauvaises herbes résistantes et parmi celles-ci l’amarante tubculée (Amarantus rudis), la petite herbe à poux (Ambrosia ambrosifolia), l’herbe à poux géante (Ambrosia trifida ), la vergerette du Canada
(Conyza canadensis), le kochia à balais (Kochia scoparia) ont été répértorié au Canada depuis 2002 (Fortin, 2017). De plus, il y a eu confirmation de la présence de moutarde des oiseaux (Brassica rapa) résistante au glyphosate au Québec (RAP, 2017). Trois mécanismes de résistance au glyphosate ont été identifiés. Premièrement, l’exclusion de la molécule, qui est généralement vacuolaire, mais qui peut aussi être liée à la nécrose rapide des tissus foliaires chez la grande herbe à poux, empêchant la translocation à l’ensemble de la plante (Sammons et Gaines ,2014). Deuxièmement, il y a la surexpression du gène responsable de la formation de l’EPSP synthase, entrainant une surproduction de l’enzyme (Sammons et Gaines, 2014). Troisièmement, il y a la mutation du site d’action du glyphosate sur l’EPSP synthase, empêchant ainsi l’action de l’herbicide (Sammons et Gaines ,2014).
Il est intéressant de noter que l’apparition du gène de résistance au glyphosate chez la moutarde des oiseaux est liée au transfert du transgène de résistance au glyphosate en provenance du canola cultivé (RAP, 2017). Il est donc important d’améliorer les bonnes pratiques culturales et d’appliquer les principes de lutte intégrée aux mauvaises herbes pour limiter l’apparition de résistance au glyphosate, qui demeure très utilisé et aux herbicides en général, ceci au Québec et ailleurs.
1.8.2 La Tembotrione
La tembotrione est un herbicide du groupe 27, de la famille des tricétones du même groupe que la mésotrione, son mode d’action lui est très semblable (SAgE pesticides, 2018). Pour ce qui est de sa persistance, elle est biodégradée dans les sols et dans l’eau à une vitesse variant de faible à élevée et sa demi-vie varie de 4,5 à 194 jours dans les sols (SAgE pesticides, 2018). En milieu terrestre aérobie, elle est dégradée en moins de 15 jours, mais en anaérobiose, sa demi-vie est supérieure à 120 jours (SAgE pesticides, 2018). Sa phototransformation est relativement élevée dans les sols et l’hydrolyse est absente à tous les pH évalués (SAgE pesticides, 2018). Elle est très soluble dans l’eau et sa faible Koc de 23 ml g-1 la rend très peu adsorbé par la matière organique (SAgE pesticides, 2018). Selon sa constante de Henry de 1,7 x 10-11 atm m3 mol-1, elle n’est pas volatile, mais elle serait en mesure d’atteindre les eaux souterraines (SAgE pesticides, 2018).
Sa toxicité aiguë est élevée, car c’est un sensibilisant cutané et à long terme, la tembotrione possède une toxicité élevée sur les yeux, le foie et les reins (SAgE pesticide, 2018). Elle est classée comme possédant des évidences suggestives de cancérogénicité chez l’humain, mais le risque chez l’humain ne peut être pleinement évalué et de plus, elle serait un perturbateur endocrinien (SAgE pesticide, 2018).