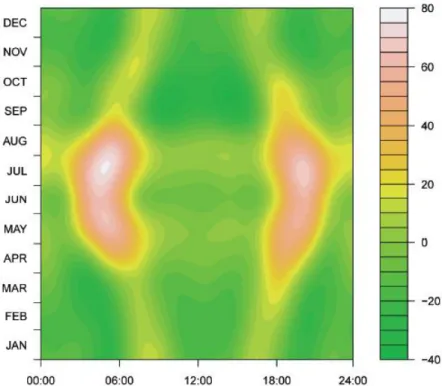
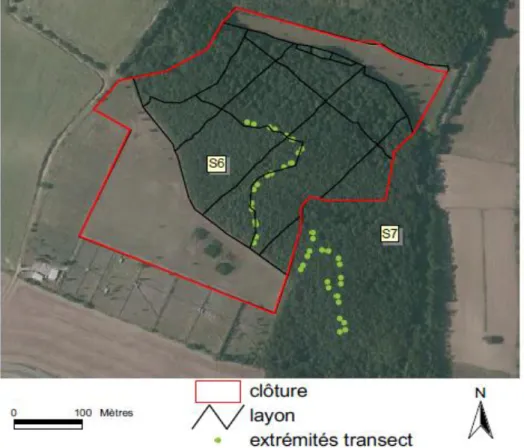
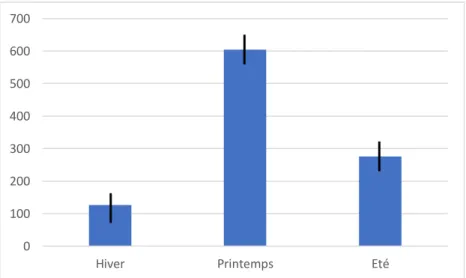
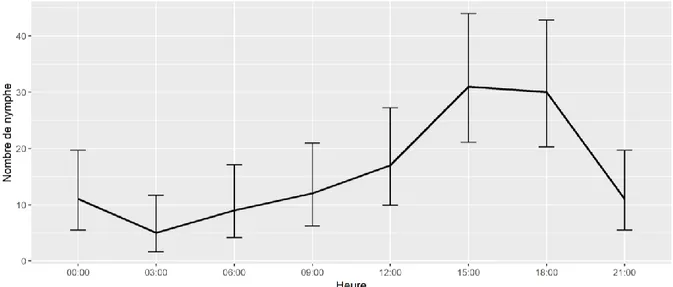
Rythme d’activité de la tique Ixodes ricinus : une synchronisation avec l’activité du chevreuil ?
70
0
0
Texte intégral
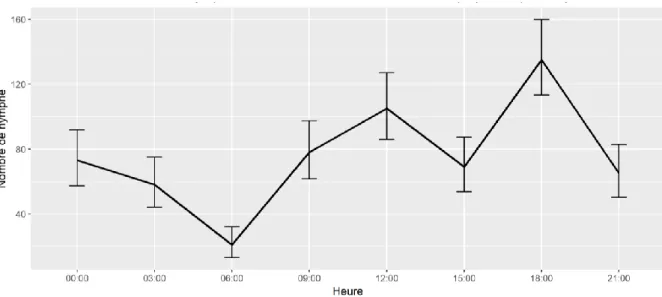
Figure
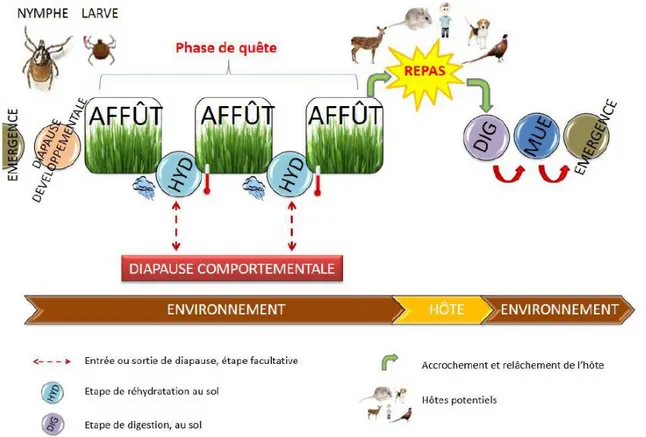
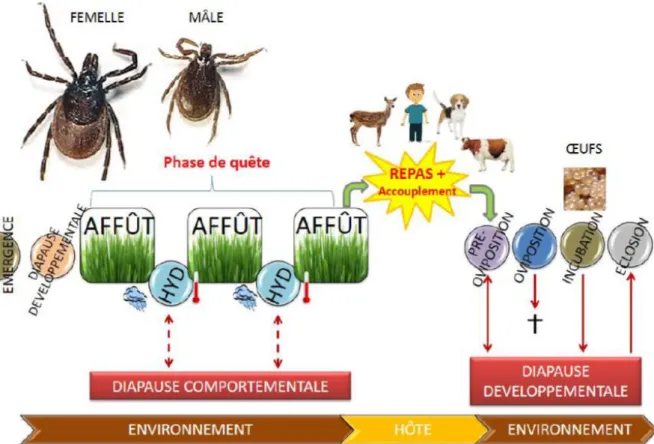
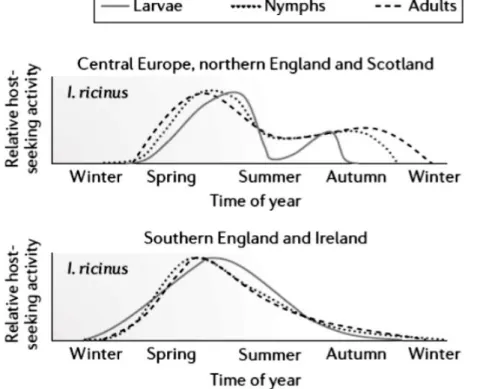
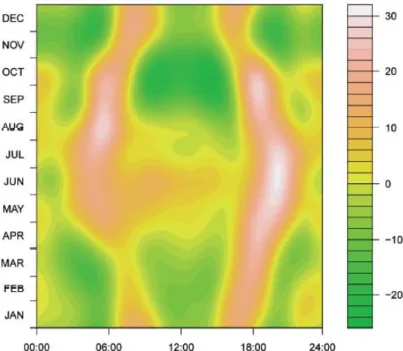
+7

Documents relatifs