Identification d'un complexe de dégradation des microARNs chez le nématode Caenorhabditis elegans
Texte intégral
Figure
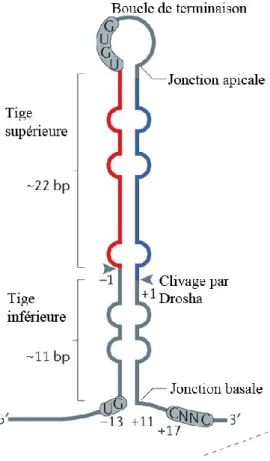
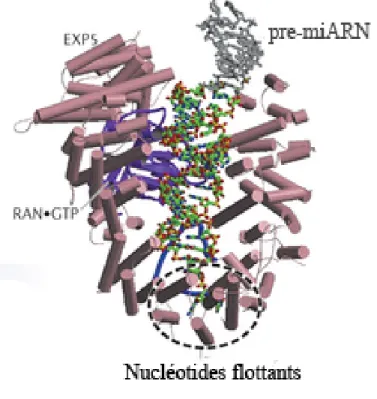
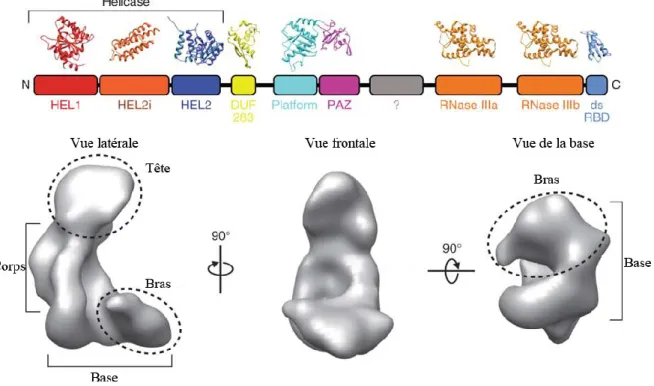
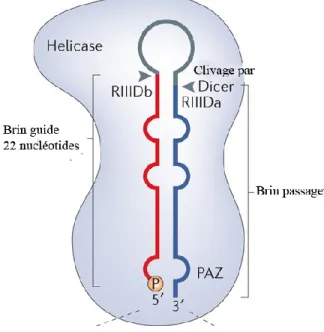
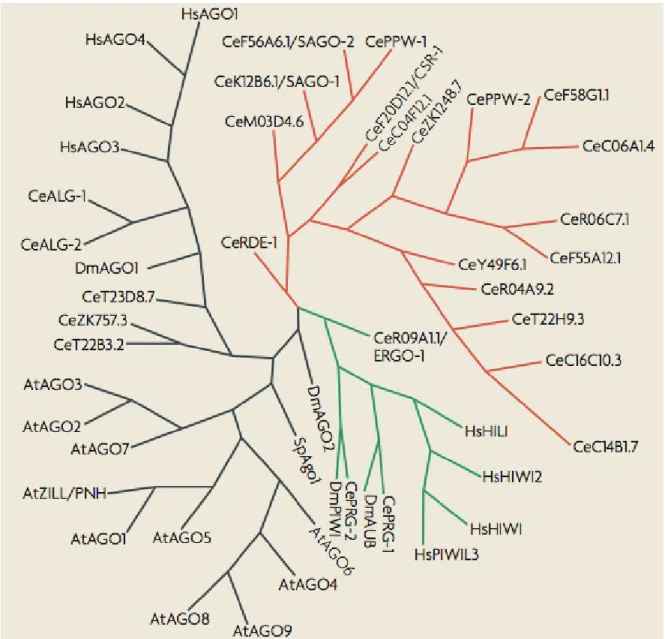
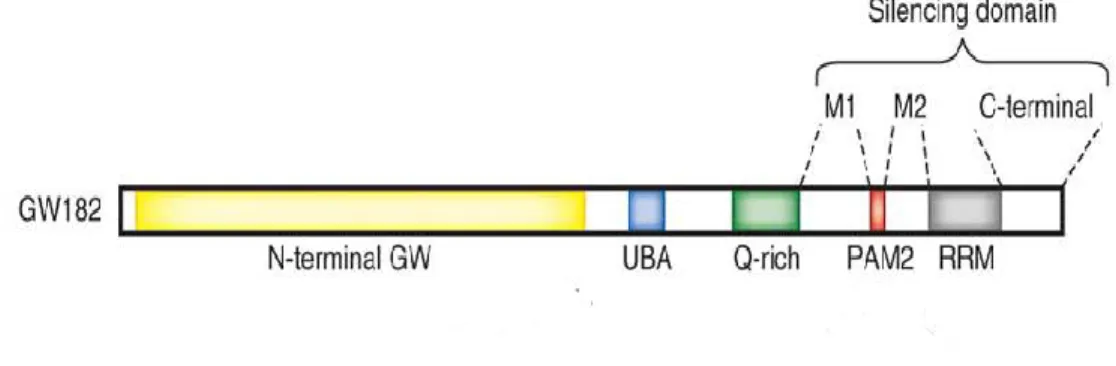

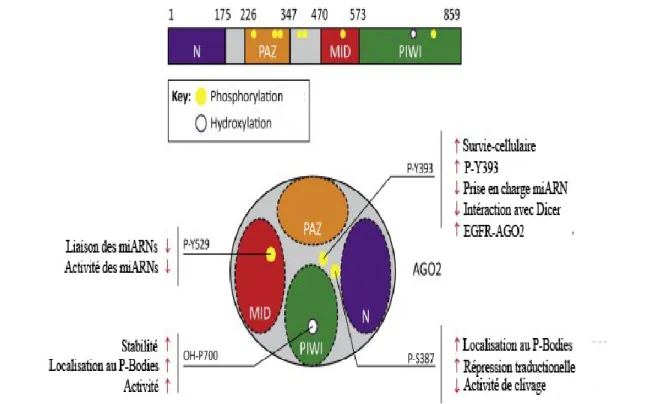
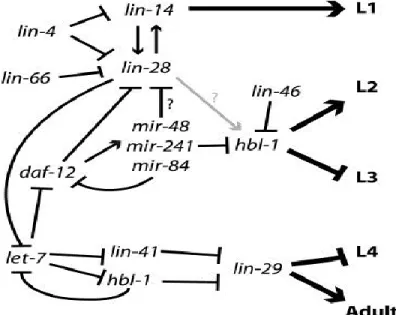
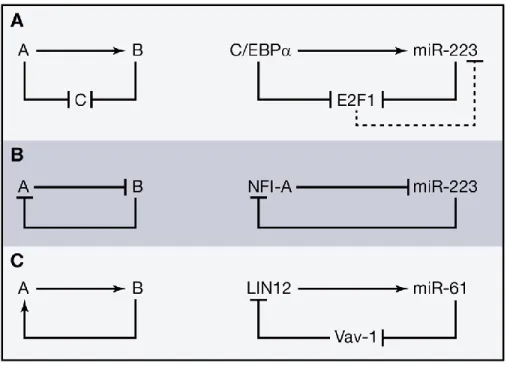
Documents relatifs
Les raisons qui poussèrent les médecins à interdire, à ignorer, à tolérer ou à encourager les pratiques artistiques entre les murs de leurs établissements
Nous pouvons ainsi faire un lien avec la définition des soins palliatifs qui « cherchent à améliorer la qualité de vie des patients et de leur famille, face
Les chapitres suivants tenteront donc de répondre à la question suivante : en fonction des deux hypothèses retenues (absolue et.. relative), quels devraient être les
That question is asked to two experiment in the Northern East of France: the first experiment is dedicated to step by step design of autonomous mixed crop dairy systems
A résistance de roche et mésoporosité de pâte équivalentes, il semble que le calcaire gris poreux initialement à l’état sec ait des propriétés mécaniques sur mortier
Boulé et Genon analysent tout d’abord, dans quelques- unes des critiques écrites par Guibert pour Le Monde et reproduites dans Photo, inéluctablement, la façon dont
Pour les dattes molles, principalement la variété Ghars, le conditionnement consiste à les tasser dans des peaux de mouton ou dans des sacs de jute ou de toile, actuellement dans
By performing a physical analysis on single Compton-scattered photons we first establish, in Section 2, the so-called imaging equation which relates the single scattered photon
