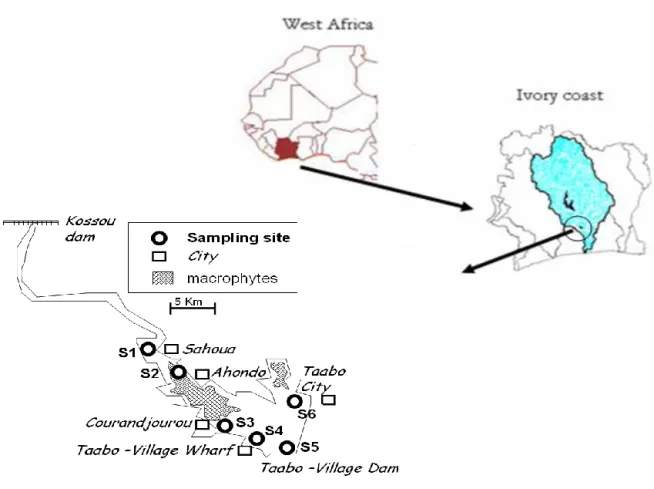
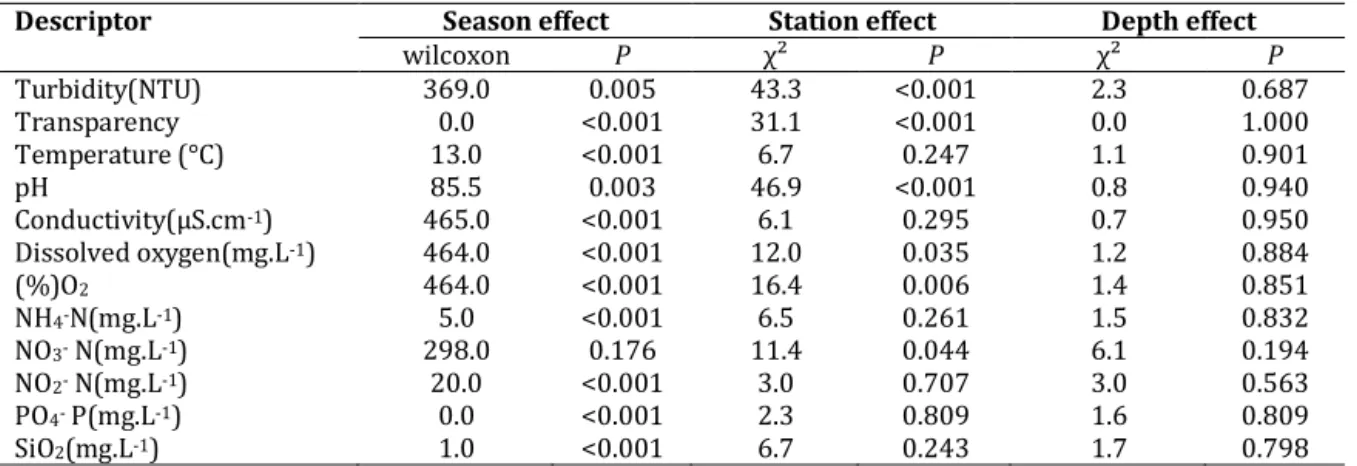
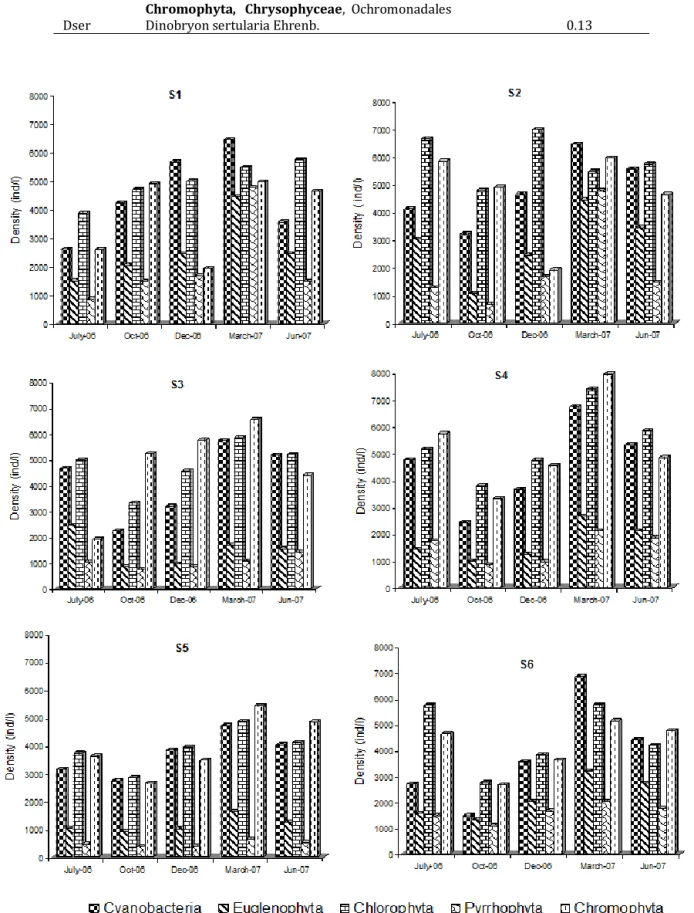
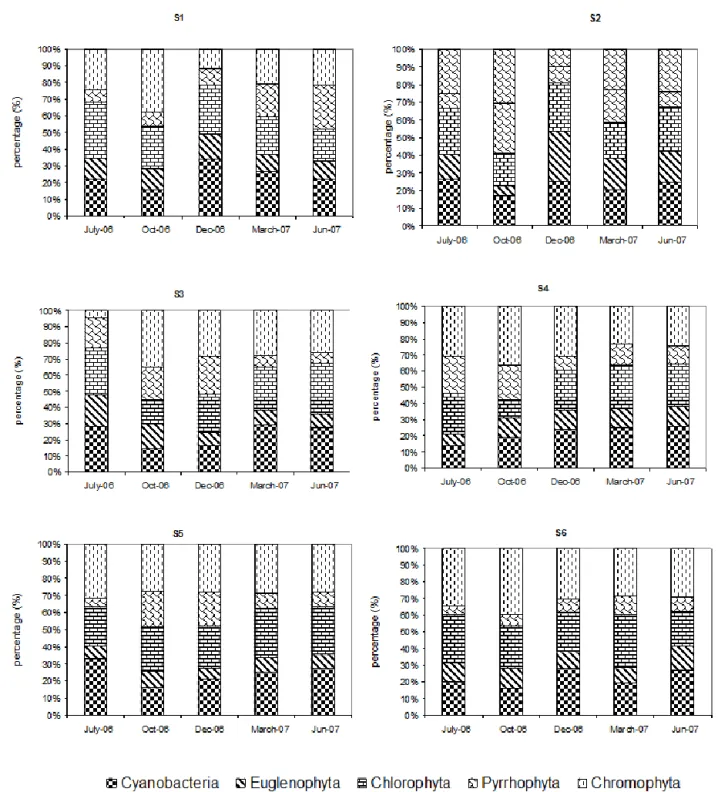
Dynamic and structure of phytoplankton community and environment in the lake Taabo (Côte d’Ivoire)
18
0
0
Texte intégral
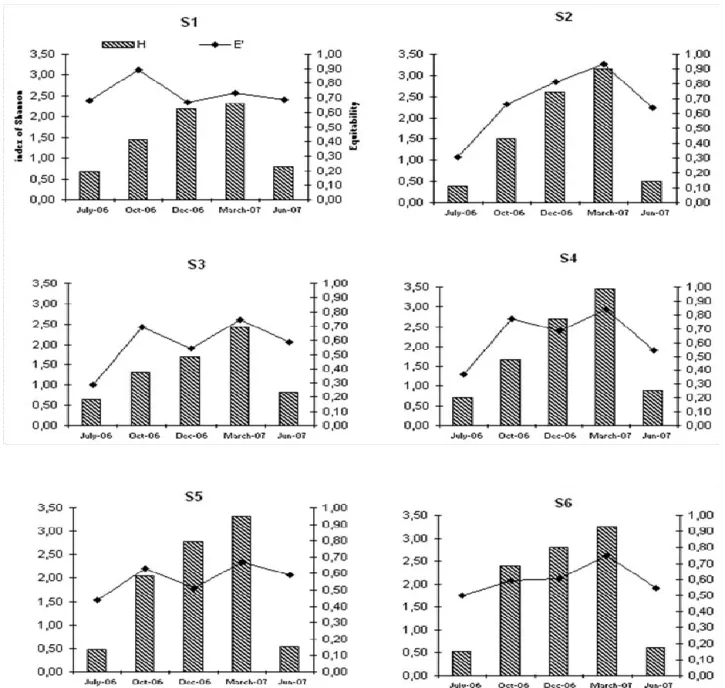
Figure


+7



Documents relatifs
